-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
谷禾健康
色氨酸是连接“吃什么”“肠道菌群状态”与“大脑和情绪”的关键枢纽。它不仅是蛋白质合成必需氨基酸,也是血清素与褪黑素的重要前体,并可通过犬尿氨酸通路参与NAD+的从头合成。
近年来,大量研究发现:色氨酸在不同代谢通路之间的“分流”,很大程度上受肠道微生物群及饮食结构(如多酚、膳食纤维、碳水化合物和脂肪比例)共同调控。这种调控一旦失衡,不仅会改变肠道屏障和免疫状态,还可通过肠–脑轴深刻影响情绪、认知以及多种神经精神和代谢性疾病的发生发展。
在这一背景下,围绕“肠道微生物–色氨酸代谢–肠脑轴”的研究迅速升温:一方面,人们逐步解析拟杆菌、梭菌、乳杆菌、双歧杆菌等代表菌属如何通过多样的色氨酸代谢途径生成吲哚类、犬尿氨酸类及血清素等关键代谢物;另一方面,肠易激综合征、自闭症谱系障碍、抑郁症、多发性硬化、阿尔茨海默病、帕金森病以及代谢综合征等疾病中,色氨酸代谢及其微生物调控异常的证据也不断被报道。
基于此情况,饮食干预、益生菌、粪菌移植乃至纳米技术等多种策略,正被尝试用于重塑肠道微生态、调整色氨酸代谢稳态,以期为肠脑轴相关疾病提供新的防治思路。
本文将系统梳理色氨酸的膳食来源和主要代谢通路,重点介绍肠道微生物中特定菌属参与色氨酸代谢的机制,阐述其在肠脑轴中的作用及与多类神经、免疫和代谢性疾病的关联,并总结目前通过饮食、益生菌、抗生素及新兴技术调控色氨酸稳态的研究进展和潜在应用价值。
▸ 色氨酸的重要性
编辑
色氨酸(Trp)是一种人体无法合成、需从饮食获取的必需氨基酸,也是血清素、褪黑素和NAD+等多种人体重要化合物的前体。
血清素:一种神经递质,在情绪调节和肠道蠕动中发挥作用;
褪黑素:一种调节睡眠-觉醒周期的激素;
NAD+:是500多种酶促反应所必需的,并且在几乎所有主要生物过程的调节中起着关键作用。
总得来说,可以概括为五个关键词:
•情绪与睡眠:是血清素和褪黑素的唯一氨基酸前体,直接影响情绪稳定、抗压能力、睡眠节律和食欲控制。
•免疫与神经保护:通过犬尿氨酸通路调节免疫反应与神经系统状态,在炎症和精神疾病中扮演关键角色。
•能量与修复:可转化为烟酸,支持 NAD⁺ 合成,保障细胞能量代谢、抗氧化、防老化和 DNA 修复。
•生长与组织结构:参与全身蛋白质合成,对儿童生长发育、肌肉维持和器官功能至关重要。
•肠道与肠—脑轴:影响肠道屏障功能和菌群代谢,通过肠—脑轴联系情绪、免疫和代谢健康。
▸ 色氨酸的主要膳食来源
①肉类
禽肉:鸡肉、鸭肉、火鸡肉(尤其是去皮鸡胸肉);
猪肉、牛肉、羊肉:瘦肉部分色氨酸含量相对更高。
②鱼类和其他水产
深海鱼:三文鱼(金枪鱼、鲭鱼等)、鳕鱼;
其他常见鱼类:草鱼、鲤鱼、罗非鱼等;
甲壳类:虾、蟹;
软体类:鱿鱼、贝类等。
③蛋类和乳制品
鸡蛋:尤其是蛋白和蛋黄整体算上;
奶及奶制品:牛奶、酸奶、奶酪(尤其是硬质奶酪,蛋白浓缩后单位重量色氨酸含量更高)。
④豆类及豆制品
豆类:黄豆、黑豆、毛豆、扁豆、豌豆、鹰嘴豆、红豆、芸豆等;
豆制品:豆腐、豆干、豆皮、素鸡、豆浆(色氨酸有一定稀释,但作为日常饮品仍有贡献)。
⑤坚果与种子
南瓜籽、葵花籽;
芝麻(黑白芝麻);
花生;
腰果、杏仁、核桃、开心果等坚果。
⑥谷物与伪谷物
全谷物:燕麦、糙米、荞麦、全麦面包/面条;
其他:藜麦(伪谷物,氨基酸组成较优)、玉米、小米等。
⑦水果(含量较低)
香蕉、牛油果(鳄梨)、榴莲;
一些干果:葡萄干、枣干等(因为脱水后营养被“浓缩”);
大多数鲜果(苹果、梨、橙子、葡萄、西瓜、草莓等)蛋白含量本身就低,色氨酸自然也少,更多作用是提供维生素、矿物质和膳食纤维。
注:咖啡豆中也含有较高含量的色氨酸。
▸ 色氨酸推荐摄入量
世界卫生组织建议最佳摄入量为4毫克/千克/天,主要用于成年人维持日常需求。
★ 不同年龄和阶段的人需求有所不同
婴儿需要更高的色氨酸摄入量,推荐剂量为13毫克/千克/天;
儿童推荐摄入量为6毫克/千克/天;
孕妇为7毫克/千克/天。
为保证获得足够氨基酸及其健康效应,研究还评估了色氨酸的可耐受最高摄入量(UL),结果为4.5g/天。不过迄今尚无因膳食色氨酸过量而致不良反应的报道。
注:高碳饮食会影响脑干和下丘脑的5-羟色胺(5-HT,也称为血清素)。膳食纤维通过调节肠道菌群也改变色氨酸代谢。微量营养素亦调控色氨酸/犬尿氨酸(KYN)通路,如维生素B6缺乏损害犬尿氨酸酶并可能降低某些脑区5‑HT合成。
食物中约90%的色氨酸由小肠吸收,吸收后,主要经犬尿氨酸(KYN)途径和血清素途径代谢。
体内约95%的色氨酸(Trp)在Trp 2,3‑二加氧酶(TDO)、吲哚胺 2,3‑二加氧酶1(IDO1)和吲哚胺 2,3-双加氧酶2(IDO2)作用下分解为喹啉酸(QA)、烟酸等产物;仅约 1%–2% 的色氨酸在色氨酸羟化酶(TpH)和芳香族氨基酸脱羧酶催化下转化为5‑羟色胺(5‑HT)。
通过5-HT、KYN和吲哚途径进行的色氨酸代谢
doi.org/10.1016/j.jfutfo.2024.09.006.
★ 肠道菌群参与重要的色氨酸代谢
若进入结肠,肠道微生物群也可以通过各种代谢途径产生多种色氨酸代谢产物。
注:早在1897年,即发现大肠杆菌和霍乱弧菌可将色氨酸转化为吲哚,提示色氨酸代谢在微生物过程及其对宿主环境影响中的重要性。
微生物群可以通过在宿主体内产生代谢物直接调节色氨酸代谢,或通过参与犬尿氨酸和血清素的产生间接影响这种代谢,而色氨酸代谢的变化会影响微生物的增殖和微生物群落的多样性。目前已鉴定出许多微生物色氨酸代谢途径,包括拟杆菌属、梭菌属、双歧杆菌、链球菌属和大肠杆菌的代谢途径。
虽然某些细菌色氨酸代谢产物的转化可以在分子层面上易于定义,但识别产生的具体代谢物类型却十分复杂。这种复杂性源于不同微生物催化酶的差异。在某些情况下,两个或多个细菌之间的协作可能是色氨酸代谢物生成的必要条件。
拟杆菌属(Bacteroides)
吲哚是通过色氨酸酶(Trpase)的酶促活性产生的,这种酶广泛存在于多种肠道细菌中,包括拟杆菌属、梭菌属和大肠杆菌。
√卵型拟杆菌、多形拟杆菌等可代谢产生吲哚乙酸
拟杆菌是人体和动物肠道中的一种主要菌群,一些拟杆菌属物种,如脆弱拟杆菌(Bacteroides fragilis)、卵形拟杆菌(Bacteroides ovatus)和多形拟杆菌(Bacteroides thetaiotaomicron),可以产生吲哚乳酸(ILA)和吲哚乙酸(IAA)。
√不同拟杆菌代谢色氨酸的产物影响不同(有的减轻肠道炎症、有的影响精神状态)
研究发现,肠道中不同拟杆菌的相关代谢物的影响不同。例如B.thetaiotaomicron可提高芳烃受体(AHR)配体吲哚乙酸(IAA)和吲哚丙酸(IPA)水平,增强 AHR 激活及相关T细胞转录因子表达。在结肠炎模型中,该菌通过激活AHR并调控CD4⁺T 细胞分化,减轻炎症性肠病。
微生物色氨酸分解物对宿主生理的作用
编辑
doi: 10.1038/s41467-018-05470-4.
通过分析无菌和单菌定植小鼠盲肠及粪便样本证实,B.ovatus可提高体内IAA浓度,重塑免疫细胞群,减轻炎症,并在三硝基苯磺酸诱导性结肠炎模型中上调结肠 IL‑22 表达;吲哚代谢物本身也能刺激免疫细胞分泌 IL‑22。研究人员认为,B.ovatus 产生的 IAA 通过促进 IL‑22 生成而对结肠炎具有保护作用。
还有研究表明,B.thetaiotaomicron和B.fragilis可诱发小鼠压力相关的抑郁样行为(如焦虑和绝望),而B.ovatus无此效应。
√同属不同菌种产生的代谢物可能差异显著
亲缘关系密切的属的不同菌株之间也存在显著的代谢差异,这很可能是由于微生物的遗传代谢和化学环境存在差异。使用四种不同的拟杆菌属菌株重建的小鼠回肠中色氨酸/犬尿喹啉酸(KYNA)途径的多种变化,支持了代谢调控的这种变异性。拟杆菌属还可能诱导中枢5-羟色胺代谢紊乱,使小鼠对应激诱导的行为障碍敏感。
√膳食缺乏色氨酸还会降低拟杆菌丰度
膳食中摄入的色氨酸也会显著影响拟杆菌属的生长状态和数量变化。研究表明,当饮食中实施严格的 色氨酸限制时,不仅会明显降低粪便中拟杆菌的丰度,还会同步重塑和改变整个肠道微生物群的组成与结构。
梭菌属(Clostridium)
梭菌属中的某些物种在吲哚衍生物的合成中发挥着重要作用。Clostridium drakei是一种能够产生乙酸的细菌,与C.scatologenes类似,可通过色氨酸的分解代谢转化3-甲基吲哚。3-甲基吲哚是一种常见的肠道代谢产物,又称粪臭素,被认为会导致猪肉产生异味。它可由脆弱拟杆菌(B.fragilis)和梭菌属(Clostridium)产生。
√丁酸梭菌抗肥胖作用还源于色氨酸代谢调节信号通路
丁酸梭菌(Clostridium butyricum)可以在肠道中产生丁酸盐,具有缓解多种疾病的潜力,包括炎症性肠病和肥胖症。
几种丁酸梭菌菌株具有抗肥胖作用,但这些作用并非完全通过提高肠道中丁酸盐的浓度来实现,还主要通过改变肠道微生物群和色氨酸(Trp)代谢来实现。丁酸梭菌可能通过产生吲哚和吲哚衍生物来激活芳香烃受体(AHR)信号通路,并调节宿主的代谢和免疫系统,从而改善肥胖。
√梭菌可能富含多种色氨酸代谢途径
最近的体外实验发现,B.thetaiotaomicron通过向大肠杆菌提供单糖,抑制其产生吲哚的能力,从而增加了对生孢梭菌(Clostridium sporogenes)的色氨酸供应,进而促进了吲哚乳酸和吲哚丙酸的产生。通过在小鼠饮食中补充富含色氨酸的食物,并增加梭菌属、乳杆菌属和双歧杆菌属的数量,重塑了小鼠肠道微生物的结构和组成,从而改善了类似抑郁的行为。
据报道,梭菌属(Clostridium)可通过色氨酸(Trp)的代谢产生吲哚、吲哚衍生物和色胺。研究发现,该属的菌株具有产生喹啉酸(QA)的潜力,且预测Clostridium hathewayi具有QA途径。然而,该菌株中缺失了负责合成QA途径前体犬尿氨酸(KYN)的基因,这表明QA途径具有代谢共享性,且包括梭菌属在内的部分生物以KYN为底物,而KYN是由包括芽孢杆菌属(Bacillus)、伯克霍尔德氏菌属(Burkholderia)和假单胞菌属(Pseudomonas)在内的另一组细菌产生的。
肠道微生物群通过肠脑轴影响色氨酸代谢
编辑
doi.org/10.1016/j.jfutfo.2024.09.006.
总体来看,梭状芽孢杆菌富含多种色氨酸代谢途径,相关分析表明其在肠道中具有更强的色氨酸代谢潜力。
乳杆菌属(Lactobacillus)
乳杆菌是正常肠道微生物群的重要成员,主要包括植物乳杆菌(Lactiplantibacillus plantarum)、干酪乳杆菌(Lactobacillus casei)、嗜酸乳杆菌(Lactobacillus acidophilus)、鼠李糖乳杆菌(Lactobacillus rhamnosus)、罗伊氏乳杆菌(Lactobacillus reuteri)、清酒乳杆菌(Lactobacillus sakei)和弯曲乳杆菌(Lactobacillus curvatus)。
√乳杆菌通过上调吲哚乙酸水平可以缓解结肠炎
乳杆菌可通过芳香族氨基酸氨基转移酶和吲哚乳酸脱氢酶(ILDH)将色氨酸转化为吲哚-3-醛(IAld)和吲哚乳酸(ILA)。研究人员筛选出色氨酸代谢活性最高的 L. plantarum,并联合 Trp 观察其在 DSS 诱导溃疡性结肠炎中的作用。
结果显示,该菌上调了吲哚乙酸(IAA)水平及 AHR mRNA 表达,进而激活 IL-22/STAT3 信号通路。通过比较不同 L. plantarum 菌株的基因组和代谢组,认为 IAA 可能是导致菌株间结肠炎缓解效果差异的关键代谢物。
√鼠李糖乳杆菌通过代谢色氨酸增强肠道屏障
筛选出可在肠道定植并在体外代谢色氨酸生成吲哚衍生物的L. rhamnosus MN-431。该菌同时上调孕烷 X 受体(PXR)和芳烃受体(AHR)的表达:大量 ILA 和 IAld 作为 AHR 配体,而共享配体 IA 主要结合 PXR,通过上调 PXR、AHR 并下调核因子 κB(NF-κB),增强肠道屏障功能。
尽管其具体吲哚代谢产物及体内代谢情况尚未完全明确,但多项证据提示L.rhamnosus对肠道益生健康具有潜在益处。
√罗伊氏乳杆菌产生的吲哚-3-醛有助于抑制结直肠癌
研究表明,吲哚-3-醛(I3A)是罗伊氏乳杆菌来源的膳食色氨酸(Trp)分解代谢产物,可诱导芳香烃受体依赖的环磷酸腺苷反应元件结合蛋白(CREB)活性,从而增强 Tc1 细胞(分泌干扰素-γ(IFN-γ)的 CD8⁺ T 细胞)的效应功能,并提高免疫检查点抑制剂的抗肿瘤效果。
当 I3A 自小肠转移至肿瘤微环境后,罗伊氏乳杆菌可激活肿瘤中的 I3A–AHR–CD8⁺ T 细胞轴,促进分泌 IFN-γ 的细胞毒性 CD8⁺ T 细胞活化。有研究发现,他汀类药物也可降低结直肠癌(CRC)风险,且该作用依赖肠道微生物群。分析显示,给药后肠上皮细胞 IDO1 基因表达受抑制,肠道色氨酸水平升高,罗伊氏乳杆菌丰度显著增加,更多 Trp 被其代谢为吲哚-3-乳酸(ILA),且二者呈正相关。ILA 通过调节 TH17 反应和维持屏障完整性抑制 结直肠癌,其中视黄酸受体相关孤儿核受体 γt 和 AHR 共同参与。
双歧杆菌属(Bifidobacterium)
双歧杆菌是经顺产和母乳喂养婴儿肠道微生物群的关键成员。母乳中色氨酸(Trp)含量高于大多数婴儿配方奶粉,且富含益生元,有助于双歧杆菌等有益菌在婴儿肠道定殖。
双歧杆菌是新生儿最早定殖的微生物之一,在其生理发育中发挥重要作用。一项研究显示,约90%新生儿在出生后肠道中检出双歧杆菌,该属在成人肠道菌群中仍占约 3%–5%。常见种类包括长双歧杆菌、短双歧杆菌、青春双歧杆菌和假长双歧杆菌等。
√双歧杆菌产生吲哚乳酸有助于抑制条件致病菌
检测从人类和动物胃肠道分离的双歧杆菌,发现吲哚乳酸(ILA)是其培养上清液中的色氨酸代谢物。来自婴儿粪便的长双歧杆菌和短双歧杆菌产生的 ILA 多于其他来源分离的菌株,因此婴儿肠道中双歧杆菌定殖所形成的 ILA 可能为其在结肠菌群中提供竞争优势。ILA 具有抗菌活性,可抑制大肠杆菌在肠道中的定植。
其他肠道微生物群
√大肠杆菌、霍乱弧菌
研究发现,大肠杆菌和霍乱弧菌(V. cholerae)可将色氨酸转化为吲哚。大肠杆菌具有活跃的色氨酸和吲哚转运蛋白,色氨酸酶可将色氨酸转化为吲哚,在大肠杆菌和乳酸菌中均有表达。然而,负责吲哚后续代谢的具体微生物酶途径,以及吲哚在其他共生菌中的分布和活性仍不明确。
√肠球菌、假单胞菌
多项研究表明,部分细菌可产生色胺(tryptamine)等神经活性分子,或通过影响肠嗜铬细胞/炎症状态间接调控宿主5-HT合成。
肠道微生物将 Trp 作为吲哚、血清素和褪黑素合成前体加以代谢,从而减少宿主可利用的 Trp。假单胞菌可利用现有色氨酸合成血清素,并通过其毒性及细胞间信号传导,使肠道菌群诱导循环 Trp 水平下降,进而影响 5-羟色胺能神经传递,以及中枢神经系统和肠神经系统(ENS)的功能。
√消化链球菌
消化链球菌(Peptostreptococcus)能够代谢色氨酸(Trp),从而产生吲哚-3-丙烯酸(IA,indole-3-acrylicacid)等吲哚类代谢物。IA对肠上皮屏障的功能有积极作用,并有助于减少免疫细胞中的炎症反应。Peptostreptococcus russellii具有苯丙氨酸脱水酶基因簇(fldAIBC),该基因簇与生孢梭菌(C. sporogenes)中的fldAIBC基因簇同源,且对苯丙氨酸的代谢至关重要。
同样,厌氧消化链球菌编码完整的fldAIBC基因簇,并产生色氨酸代谢产物吲哚丙酸(IPA)和异亮氨酸(IA),而口腔消化链球菌则缺乏初始因子fldI,导致色氨酸代谢产物水平较低。
小结
围绕特定肠道菌属或菌种的色氨酸代谢开展研究,有望为微生物群靶向调控色氨酸代谢提供新思路,例如通过基因序列同源性筛选具色氨酸代谢能力的菌株。
此类研究有助于构建基于肠道微生物群的精准干预策略,以改善 Trp 代谢失衡相关疾病。聚焦特定菌种或菌株与色氨酸的相互作用,科学家可开发更精细的调控手段,优化这一关键代谢通路,从而促进整体健康并应对多种代谢性疾病。
肠道微生物群通过产生激素、免疫因子和代谢物,在大脑行为和认知发育中发挥重要作用。出生后,肠道细菌的早期定殖可促进中枢神经系统发育。肠道微生物群影响中枢神经系统的结构和功能,并可调节情绪。
★ 肠道微生物群通过产生激素、信号分子等影响神经系统功能
动物研究表明,微生物群失衡常导致焦虑样行为;相反,特定益生菌干预或重建微生物群则有助于改善焦虑相关行为:缺乏正常肠道微生物群会严重干扰中枢神经传递,而肠道微生物群恢复后,小鼠的焦虑行为可恢复正常。
肠道微生物群可通过产生神经信号分子或调节内分泌细胞激素影响中枢神经系统功能。研究证实,口服婴儿双歧杆菌后,大鼠脑内 5-HTP、5-HIAA 和苯乙酸水平显著高于对照组。肠道微生物群还能通过影响内分泌细胞皮质酮水平调节 HPA 轴反应强度,在应激反应中发挥关键作用,并调控促肾上腺皮质激素释放因子和脑肠肽等激素的分泌。
注:脑肠肽是一类同时存在于大脑和肠道的活性肽类物质,通过脑-肠轴双向调节消化、代谢、情绪等生理功能,是神经、内分泌、免疫系统的重要信号分子。
编辑
★ 色氨酸代谢是肠脑轴中的重要组成部分
肠脑轴(GBA)描述了大脑与肠道之间的双向交叉。GBA拥有复杂的沟通系统,包括中枢神经系统(CNS)、肠神经系统(ENS)和胃肠道(GI)之间的双向相互作用。
脑肠相互作用主要有三个机制:神经通路、内分泌通路和免疫通路,微生物积极参与GBA内的代谢途径,影响微生物-肠-脑轴(MGBA)相关代谢产物。这些包括微生物产生的神经递质、短链脂肪酸,以及色氨酸等的直接或间接代谢。这些过程调节下丘脑-垂体-肾上腺(HPA)轴和细胞因子的产生,以调节免疫力。
色氨酸代谢及其相关代谢物对肠脑轴至关重要。微生物肠脑轴直接影响情绪和心理状态,还与免疫系统密切相关,并且与胃肠道慢性疾病相关。
▸ 血清素通路与肠脑轴
色氨酸(Trp)及其代谢物的三条关键代谢途径在大脑与肠道间的沟通中发挥重要作用。约 90% 的体内血清素由肠嗜铬细胞(EC)合成:Trp 先经色氨酸羟化酶1(TPH1)转化为5-羟色氨酸(5-HTP),随后进一步生成血清素(5-HT)。血清素作为核心胃肠信号分子,在肠道内外神经元之间传递信息,调节肠道蠕动与运动、上皮分泌、血管扩张及营养吸收,从而影响整体消化功能。
√色氨酸代谢调节血清素水平进而影响多种组织
在中枢神经系统与胃肠道的双向通讯中,血清素在中枢和外周均作为神经递质发挥作用。外周血清素不能通过血脑屏障,而色氨酸可跨越血脑屏障并在中枢合成血清素。因此,色氨酸代谢的改变会影响外周色氨酸的利用和中枢色氨酸水平,从而调节中枢血清素代谢。
经肠脑轴介导的胃肠神经调控可通过改变血清素信号传递,影响中枢神经系统的神经传递。早期神经元具有类似内分泌器官的功能,可从大脑释放多种生物活性物质(如血清素),既参与脑发育调控,又调节内脏靶器官(如胃肠道)及全身多种组织的发育。
例如,血清素(5-HT)通过其受体参与心血管调节,这些受体在调控心血管反射以及交感、副交感神经活动中具有重要生理作用。血清素受体分布于多种心血管组织及参与心血管调节的中枢区域,其激活可对心血管功能产生多方面影响。中枢血清素缺乏会导致 5-HT 能通路异常、血清素能支配减弱,并因血清素下降而诱导海马脑源性神经营养因子表达升高,引起过度神经支配。血清素水平改变也被认为与自闭症、精神分裂症等神经精神疾病的发生密切相关。
√血清素是肠神经系统的关键信号分子
肠神经系统(ENS)由胃肠道壁内的神经成分构成,是一个独立而复杂的神经网络,可在相对独立于中枢神经系统的情况下调节肠功能。
但 ENS 又通过交感和副交感神经与中枢神经纤维相连,对肠蠕动和局部血流等基本胃肠功能至关重要。5-HT 能神经元是 ENS 中最早出现的神经元之一,而 5-HT 是 ENS 的关键信号分子。
5-HT 受体的激活与成人期神经发生和神经保护相关;成人 ENS 的正常发育依赖5-HT受体4特异性信号,神经元来源的 5-HT 与黏膜来源的 5-HT 通过互补作用共同促进肠神经系统成熟。
▸ 犬尿氨酸路径与肠脑轴
将色氨酸代谢为犬尿氨酸(KYN)的关键酶包括色氨酸 2,3‑二加氧酶(TDO)、吲哚胺 2,3‑二加氧酶1(IDO1)和吲哚胺 2,3-双加氧酶2(IDO2)。其中,TDO以肝脏表达为主;肠道黏膜及免疫相关组织中IDO1更为关键。
√犬尿氨酸代谢影响中枢神经系统,并与神经疾病相关
约95%的色氨酸经犬尿氨酸通路(KP)代谢为烟酰胺和NAD+。TDO 负责启动 KP,是该通路的主要限速酶;肝外 KP 分支则由两种 IDO 酶调控,IDO 主要在黏膜组织(如免疫系统和肠道)中活跃。KYN 通常先被羟基化为 3-HK,再转化为 3-HAA;HAA 通过非酶促中间反应迅速生成喹啉酸(QA),并进一步转化为 NAD+,为 KP 的主要终产物。另一支 KP 则在 KAT 作用下生成犬尿氨酸(KYN),参与神经调节。
KYN 及其代谢物可影响中枢神经系统,并与多种神经系统疾病相关。约 60% 的外周 KYN 可通过血脑屏障进入中枢,在星形胶质细胞中转化为犬尿喹啉酸(KYNA),其水平与神经保护相关;而喹啉酸(QA)由小胶质细胞产生,其升高与神经元兴奋性毒性有关。KYNA 与 QA 之间的失衡被认为与抑郁症、精神分裂症和多发性硬化等神经系统疾病的发生密切相关。
注:在肠神经系统(ENS)和中枢神经系统(CNS) 中,犬尿喹啉酸(KYNA)作为 N-甲基-D-天冬氨酸受体和 α7 烟碱型受体的拮抗剂,同时又是G蛋白偶联受体 GPR35 的激动剂。
▸ 色氨酸代谢物与肠脑轴
肠道微生物群可直接利用色氨酸(Trp),将约 4%–6% 转化为吲哚、色胺及吲哚酸衍生物。
肠道菌群将色氨酸转化为色胺和吲哚-3-丙酮酸(IPYA),后者进一步生成吲哚、吲哚-3-乙醛(IAAld)和吲哚乳酸(ILA)。IAAld 可转化为吲哚乙酸(IAA)和色氨酸,IAA 还可继续代谢为粪臭素;ILA 则可转化为 IA,随后生成 IPA。
√色氨酸经肠道微生物代谢产生的吲哚和色胺影响神经系统
吲哚是调节肠道的重要信号分子。吲哚及其衍生物(如 IAA)可作为 AHR 配体,通过激活小胶质细胞和星形胶质细胞介导中枢神经系统炎症。AHR 配体可能通过干扰素α受体1相关通路,发挥抗炎和神经保护作用,从而调控中枢神经系统炎症。
吲哚作为 AHR 配体,还能通过影响电压门控钾通道和线粒体 NADH 脱氢酶,调节小鼠结肠 L 细胞中胰高血糖素样肽-1(GLP-1)的分泌。研究证实,肠源性吲哚代谢物可能作用于扩展奖赏网络,尤其是杏仁核–伏隔核以及杏仁核–前岛叶皮层通路。
微生物代谢产物扩散穿过肠道屏障后,可经迷走神经将信号由胃肠道传递至中枢神经系统。在大鼠中鞘内常规注射吲哚会升高脑内吲哚和色胺水平,激活迷走神经,降低运动活动,并诱导 c-Fos 过度表达。c-Fos 是迷走神经激活的重要标志,提示吲哚可激活肠黏膜中的迷走神经传入纤维。
色氨酸(Trp)作为必需氨基酸,不仅是蛋白质合成的基本原料,也是连接肠道微生物群与中枢神经系统的重要代谢枢纽。色氨酸代谢一旦在通路比例或关键代谢物水平上发生失衡,既可影响情绪与认知等神经精神功能,又与抑郁症、精神分裂症、多发性硬化等中枢神经系统疾病,以及多种慢性胃肠道疾病的发生和进展密切相关,因此成为近年来肠脑轴相关疾病研究与干预的核心焦点之一。
肠脑轴相关疾病及治疗策略
编辑
doi.org/10.1016/j.jfutfo.2024.09.006.
▸ 肠易激综合征(IBS)
肠易激综合征(IBS)是一种常见的肠道功能紊乱性疾病,可表现为腹痛、腹泻以及伴发的焦虑和抑郁。
√肠脑轴失衡是IBS的重要特征
肠脑轴(GBA)失衡是 IBS 发病机制的突出特征之一,而肠道微生物群及其代谢产物在其中起关键调控作用。多项针对 IBS 患者粪便样本的研究发现,在门水平上,Bacillota和Bacteroidota比例升高,而双歧杆菌(Bifidobacterium)和乳杆菌(Lactobacillus)比例降低;链球菌(Streptococcus)和瘤胃球菌(Ruminococcus)数量亦有所增加。
肠道微生物群结构和组成的改变与多种胃肠道疾病相关,包括 IBS。据估计,约10%的IBS发生于胃肠炎之后,此类患者的微生物特征与腹泻型 IBS(IBS-D)及单纯腹泻患者高度相似。胃肠蠕动异常是 IBS 的重要表现之一,而肠道微生物群及其代谢物可通过影响肠道神经元、胶质细胞和肠壁肌层巨噬细胞等多种途径调节胃肠动力。
√肠易激综合征患者存在血清素合成及功能受损
目前关于 IBS 机制的研究部分聚焦于色氨酸向血清素(5-HT)的转化通路,该通路同时调节胃肠功能和情绪。据报道,IBS 患者结肠和直肠中色氨酸羟化酶1(TPH1)表达降低,导致 5-HT 合成及其功能受损。
还观察到 IBS 患者的犬尿氨酸(KYN)水平升高,IDO 活性显著增强,并且与症状严重程度呈正相关;与此同时,KYN 代谢产物犬尿喹啉酸(KYNA)水平下降,KYNA/KYN 比值降低,提示 KYNA 与 IBS 发病呈负相关。KYNA 对中枢神经系统和肠道具有抗炎、镇痛及情绪保护作用,因此其代谢失衡可能构成 GBA 功能紊乱的重要基础。
▸ 自闭症谱系障碍(ASD)
自闭症谱系障碍(ASD)是一种复杂的神经发育障碍,主要影响沟通、社交能力和行为。许多儿童和成人 ASD 患者伴有明显的胃肠道症状,如便秘和腹泻,且这些症状与 ASD 的严重程度相关。
√自闭症患者肠道多样性降低
研究表明,这些胃肠道表现与肠道微生物群及其代谢物失衡有关,可影响胃肠功能和神经生物学状态。ASD 及其他神经发育障碍患者的粪便微生物群α多样性降低,且与健康对照组相比,自闭症谱系障碍患者的Bacillota与Bacteroidota比值升高。
√某些微生物可能通过影响色氨酸可用性参与自闭症的发病机制
几项评估自闭症患者肠道微生物丰度差异的研究将自闭症症状与Prevotella、Coprococcus、Veillonellaceae丰度较低联系起来。
脆弱拟杆菌(一种胰蛋白酶合成细菌),可能会降低自闭症患者的色氨酸可用性。非色氨酸衍生的微生物代谢产物也可能起到因果作用,一项观察自闭症小鼠母体免疫激活(MIA)模型中肠道微生物代谢产物的研究显示,微生物代谢产物4-乙基苯基硫酸盐增加了46倍,如果小鼠被脆弱拟杆菌定殖,则其正常化。
√自闭症患者存在色氨酸代谢异常
多项研究提示ASD患者存在色氨酸(Trp)代谢异常:血浆Trp水平降低,KYN/Trp比值升高,说明 Trp 代谢由血清素合成通路偏向犬尿氨酸通路(KP)。
研究还发现,ASD 患者细胞利用 Trp 作为能量底物的能力低于对照组,提示 Trp 代谢减弱可能通过影响大脑早期发育、线粒体稳态及脑内免疫代谢通路而发挥作用。进一步证明,ASD 儿童与对照儿童在尿代谢物上的主要差异集中于色氨酸和嘌呤代谢通路,ASD 儿童更倾向于将 Trp 代谢为黄尿酸和犬尿喹啉酸,同时减少 KYNA 生成。对这些代谢异常的深入认识有望促进 ASD 的早期和更可靠的诊断。
▸ 抑郁症
抑郁症是一种常见的精神障碍,也是当代社会致残的主要原因之一。患者常表现为情绪低落、绝望、快感缺失、疲乏及睡眠障碍。
√肠道微生物群可能通过多条通路影响抑郁
有研究发现,抑郁症患者的肠道微生物群组成和结构与健康人明显不同,提示“肠–脑”调控具有双向性。肠道微生物群可能通过影响多条在抑郁中被扰乱的通路(包括色氨酸代谢和免疫系统)参与其发病。血清素可用性下降是抑郁症的重要特征,而其他色氨酸代谢产物(如犬尿氨酸)同样与抑郁密切相关,这些代谢改变很可能由肠道微生物群驱动。
√色氨酸及血清素水平紊乱影响抑郁症状
作为关键神经递质,血清素(5-HT)参与调控中枢神经系统的应激与适应反应,其紊乱与抑郁发作密切相关。突触间隙 5-HT 含量下降可诱发抑郁,且抑郁患者突触后膜中 5-HT 受体密度与敏感性均降低。降低吲哚胺 2,3‑二加氧酶(IDO)表达、提高脑内5-HT水平被认为是潜在治疗靶点。
血浆色氨酸降低及犬尿喹啉酸(KYNA)/色氨酸(Trp)比值升高与抑郁相关;同时,抑郁患者血浆 KYNA/喹啉酸(QA)比值下降:KYNA 通过激活 GPR35 调节 cAMP 生成,并抑制交感神经元和星形胶质细胞的Ca2+通道,降低炎症转录因子 NF-κB 表达,抑制炎症相关通路,具有神经保护和抗炎作用;相反,喹啉酸(QA)具神经毒性,参与抑郁发生。肠上皮 GPR35 还能通过微生物群调控 IAld 与 ILA 等代谢产物的平衡;GPR35 缺失可诱导类抑郁行为,而在此背景下补充吲哚-3-醛(IAld)具有潜在抗抑郁效应。
√肠道微生物群通过调节色氨酸代谢影响抑郁
色氨酸代谢的紊乱是抑郁症病理的重要因素,肠道微生物群在影响这一代谢过程中起着关键作用。研究发现褪黑素以微生物群依赖性的方式改善色氨酸代谢,缓解卵巢切除小鼠的抑郁样行为。
细菌通过多种机制在抑郁症中发挥作用,益生菌与其他细菌的结合可以通过多种机制缓解抑郁症。探索多种益生菌疗法、益生元和共生菌对于进一步了解它们恢复色氨酸代谢和缓解抑郁行为的潜力至关重要。
▸ 多发性硬化症
多发性硬化症(MS)是一种常见的中枢神经系统自身免疫性疾病,也是青年人致残的主要原因之一。研究发现,MS 患者的色氨酸(Trp)水平显著低于健康对照,且 Trp 代谢关键产物犬尿喹啉酸(KYNA)与喹啉酸(QA)的平衡变化会通过调节犬尿氨酸通路(KP)影响疾病进程,从而有望缓解 MS 症状。
√丁酸梭菌调节色氨代谢减轻多发性硬化症
实验性自身免疫性脑脊髓炎(EAE)是研究 MS 的经典动物模型。研究显示,该模型中肠道微生物群可产生犬尿喹啉酸(KYNA),进而招募并激活 GPR35 阳性巨噬细胞,促进 EAE 发展。特别值得注意的是,在无菌小鼠中,只有能够产生 KYNA 的大肠杆菌菌株才能恢复 EAE 表型,而野生型大肠杆菌不足以诱导相同效应,突出了 KYNA 在该过程中的特异作用。
鉴于肠道微生物群在其中发挥关键调控作用,通过益生菌(如丁酸梭菌)调节色氨酸代谢通路,有望减轻神经退行性疾病。肠源性色氨酸代谢物还能作为 AHR 配体调控星形胶质细胞,从而影响中枢神经系统炎症。
▸ 神经退行性疾病
神经退行性疾病包括阿尔茨海默病(AD)和帕金森病(PD)等,其中 PD 的特征为多巴胺能神经元逐渐丧失。研究表明,与对照相比,PD 患者血浆和血清色氨酸浓度明显降低,血清色氨酸的下降可能与其常见的心理症状相关。AD 患者外周血中色氨酸降解增加,同样导致血清色氨酸水平下降。
√调节色氨酸代谢影响中枢神经从而减轻帕金森病
在帕金森病(PD)中,3-羟基犬尿氨酸(3‑HK)水平升高,可能通过诱导氧化应激损伤神经元;而 KYNA 水平降低,外源性 KYNA 注射可减轻 PD 症状。
进一步研究发现,通过调节 TDO 活性和色氨酸代谢,可影响中枢神经系统内外多种KYN通路神经活性代谢物的水平;抑制 TDO 既能增强神经保护、减少蛋白毒性、预防记忆减退,又有助于缓解胃肠功能障碍,有望成为帕金森病治疗的新靶点。
√色氨酸代谢产物调控阿尔茨海默病的Aβ沉积
阿尔茨海默病(AD)的主要病理特征为神经原纤维缠结和β淀粉样蛋白(Aβ)沉积。小胶质细胞和星形胶质细胞在 AD 的进展中起关键作用,色氨酸及其代谢物可调控这两类细胞的活化,从而改善认知功能并抑制 Aβ 沉积。
阿尔茨海默病患者体内3-羟基犬尿氨酸(3‑HK)和喹啉酸(QA)水平均升高,其中 QA 在多种神经系统疾病中已被证实具有神经毒性。相比之下,犬尿喹啉酸(KYNA)具有神经保护作用;部分研究发现,KYNA 和5-羟基哚烯乙酸(5-HIAA)能诱导蛋白酶 Neprilysin 的表达,进一步支持色氨酸代谢产物在脑内调控 Aβ 水平的作用。
注:Neprilysin(脑啡肽酶,NEP)是一种锌依赖性金属蛋白酶,在人体内具有多种重要的生理功能,主要涉及肽类物质的降解。
▸ 代谢综合征
代谢综合征(MetS)包括腹型肥胖、高血压、血脂异常和胰岛素抵抗等心血管危险因素。
√代谢综合征患者的色氨酸代谢紊乱
在肥胖和 MetS 人群中,犬尿氨酸(KYN)、犬尿喹啉酸(KYNA)和喹啉酸(QA)水平升高,犬尿氨酸/色氨酸比值上升,且尿酸、甘油三酯与色氨酸(Trp)向 KYN 的转化率呈正相关。受炎症状态调节的吲哚胺2,3-二氧化酶1(IDO1)将色氨酸代谢为犬尿氨酸。犬尿氨酸途径的抑制剂可能用于治疗代谢综合征。
外周血清素(5‑HT)水平升高及色氨酸羟化酶1(TPH1)多态性与肥胖相关。研究表明,缺失 TPH1 的小鼠在高脂饮食下可避免肥胖、胰岛素抵抗和非酒精性脂肪肝,提示外周 5‑HT 抑制能量消耗,而肥胖可反过来提高外周 5‑HT 水平。不过,也有研究发现 5‑HT 与肥胖呈负相关,说明其调控机制复杂且尚未完全阐明。此外,降低 5‑HT 活性的 TPH2 变异会增加抑郁症患者发生MetS的风险。
√色氨酸转化产物吲哚衍生物调控代谢
微生物群通过色氨酸转化产生的几种吲哚衍生物可能在代谢综合征的发病机制中起作用。
吲哚本身已被证明可以刺激肠内分泌 L 细胞产生胰高血糖素样肽-1 (GLP-1),这是一种刺激胰腺 β 细胞分泌胰岛素的肠降血糖素。这种机制涉及快速抑制刺激 GLP-1 分泌的电压门控 K+ 通道,但受 ATP 合成抑制的长期影响控制,减少GLP-1分泌。
除神经系统异常外,肠脑轴(GBA)相关疾病患者还常伴有便秘、腹泻等胃肠道症状,其中肠道微生物群失调及其代谢物改变是重要原因。
因此,越来越多研究尝试通过靶向肠道微生物群治疗此类疾病。愈来愈多证据将胃肠功能障碍与肠脑轴相关疾病联系起来,肠道极可能成为这类患者的潜在治疗靶点。
饮食
饮食被认为是影响微生物色氨酸代谢的重要因素。
√多酚、膳食纤维调节肠道菌群及色氨酸代谢
最新研究发现,小米中的多酚化合物奎宁酸通过改善肠道微生物群的组成并增加微生物代谢物吲哚乙酸和犬尿氨酸发挥抗炎作用,为预防阿尔茨海默病(AD)提供了潜在靶点。
膳食纤维可以减少吲哚的产生,同时促进其他有益健康的色氨酸代谢产物的生成。
√富含麦麸的饮食调节色氨酸代谢物的合成和转化
富含麦麸的饮食有效地抑制了色氨酸向犬尿氨酸途径代谢物的转化,同时增加了褪黑激素和微生物分解代谢物,即吲哚-3-丙酸、吲哚-3-乙醛和 5-羟基-吲哚-3-乙酸。
麦麸增加了促进健康的细菌(例如,Akkermansia和Lactobacillus),它们与色氨酸衍生的吲哚类代谢物显著相关。
富含麦麸的饮食还可有效调节与免疫功能相关的微生物转化和色氨酸合成(即增加 AhR 和 IL-22 的结肠表达),同时改善葡萄糖和脂质稳态,以及增加肠道健康促进菌的丰度。
√碳水化合物:影响色氨酸代谢速率
微生物色氨酸代谢速率可能受肠腔内营养物质(如碳水化合物)可利用性变化的影响。
体外研究显示,从仔猪粪便中分离的一株利用色氨酸的细菌,在可消化碳水化合物(葡萄糖)存在时,主要将色氨酸用于细菌蛋白质合成;而在不可消化碳水化合物(低聚果糖)存在时,则以其为底物产生吲哚。
进一步研究表明,通过添加低聚果糖、抗性淀粉等不可消化碳水化合物,可提高碳水化合物可利用性,促进其代谢并增加短链脂肪酸生成,同时减少色氨酸降解及吲哚类化合物的产生。
这些结果说明,提高碳水化合物可利用性可抑制肠道微生物对色氨酸的降解,进而影响循环色氨酸库。相反,碳水化合物供应增加还能促进肠道血清素(5‑HT)合成,并与胃肠道运动增强相关——这一点已在口服多糖的小鼠实验中得到证实。微生物产生的短链脂肪酸增加也可能参与其中。
√高脂肪饮食:抑制色氨酸向吲哚代谢物的转化
研究表明,高脂饮食会耗尽小鼠盲肠中的微生物代谢产物吲哚乙酸和色胺,提示在高脂状态下微生物色氨酸降解途径减弱。
同时,高脂饮食显著提高小鼠肠道 IDO 活性,促进色氨酸向犬尿氨酸的分解代谢。肠道微环境在高脂暴露下发生改变,抑制了微生物将色氨酸转化为吲哚类代谢物,尤其是吲哚‑3‑丙酸、吲哚‑3‑乳酸和吲哚乙酸盐。这些代谢物是 AhR 激动剂,在免疫调节中具有关键作用。
益生菌
益生菌,如属于乳杆菌属和双歧杆菌属的细菌,对色氨酸代谢产生有益影响。
√益生菌促进血清素合成
一方面,益生菌,如乳杆菌和双歧杆菌中的物种,可以直接将色氨酸转化为血清素。
另一方面,一些益生菌乳杆菌菌株,如干酪乳杆菌,可以通过增加TPH1表达间接促进结肠血清素合成。
√益生菌与犬尿氨酸途径的调节密切相关
与血清素水平升高相一致,大鼠口服约氏乳杆菌(Lactobacillus johnsonii)无细胞上清液后,血清犬尿氨酸水平及肠道 IDO 活性均下降。连续口服约氏乳杆菌 8 周的人体试验亦观察到血清犬尿氨酸降低,且色氨酸含量呈上升趋势。
此前研究还发现,大鼠补充益生菌婴儿双歧杆菌(Bifidobacterium infantis)可升高色氨酸水平,并降低血循环中犬尿氨酸/色氨酸比值。
上述结果提示,部分乳杆菌和双歧杆菌益生菌可能通过抑制犬尿氨酸途径来调节宿主色氨酸代谢。
慎用抗生素
√抗生素会调节肠道微生物群对色氨酸的代谢
已有研究报道,抗生素所致的微生物变化会影响犬尿氨酸通路:微生物群耗竭可提高小鼠和猪体循环中色氨酸的可用性,并减弱其沿犬尿氨酸通路的代谢。同时,一些研究也发现,抗生素诱导的菌群改变影响猪体内微生物色氨酸降解途径的激活。随着循环色氨酸升高,口服抗生素减少了空肠中色氨酸的可用性,并降低猪大肠中微生物色氨酸脱羧活性,却增加了大肠中吲哚及其衍生物含量。
有趣的是,最新研究发现,将主要作用于大肠菌群的广谱抗生素输注至回肠末端,反而使血循环中色氨酸水平下降,并增强微生物对色氨酸的降解,从而提高大肠吲哚水平。与既往结果相对,这提示肠道微生物群在应对不同抗生素干预时,对色氨酸代谢具有复杂而独特的调节作用。
其他方法
√粪菌移植通过影响肠道菌群调节色氨酸代谢
粪便微生物群移植(FMT)也被证明对受体结肠内菌群的色氨酸代谢具有调节作用,通过设计合成菌群,可以改善肠道微生物群的功能障碍,包括色氨酸代谢。
√纳米技术、增加多糖利用位点
一些能够代谢色氨酸的稀有细菌,如B.thetaiotaomicron,可以通过多个多糖利用位点来增加其丰度。纳米技术是一种在微观层面操纵肠道微生物群相互作用的新方法。该技术在诊断和治疗肠脑轴(GBA)相关疾病方面不断取得进展
还有研究人员设计了一种口服活生物治疗剂,利用乳酸乳球菌(Lactococcus lactis),该治疗剂能增强小肠靶向性并促进外源性乳酸乳球菌的产生,从而实现大脑功能的精确调控。
下表总结了相关疾病患者中益生菌、抗生素及饮食干预等的临床试验,显示这些干预可更有效缓解神经系统疾病相关的胃肠症状。
编辑
编辑
doi.org/10.1016/j.jfutfo.2024.09.006.
注:然而,目前尚无确切证据证明这些干预能缓解神经系统问题,难以直接肯定或否定其疗效,因为它们可能对由不同因素导致的神经系统疾病产生不同影响。
肠道微生物群、肠脑轴与色氨酸代谢共同构成了贯穿局部肠道环境与全身神经功能的核心网络。不同菌群通过竞争性利用色氨酸、分泌多种吲哚及其衍生物、调节血清素及犬尿氨酸等关键代谢通路,不仅重塑肠黏膜屏障和局部免疫微环境,也通过迷走神经、循环代谢物和炎症信号将“肠道事件”转译为情绪、认知及行为的改变。可以说,肠道微生物群对色氨酸代谢“走向”的精细调控,是肠脑轴发挥双向调节功能的重要生物学基础。
在此背景下,许多神经精神及肠道相关疾病可被重新理解为“微生物群–色氨酸代谢–肠脑轴”失衡的不同表现:一端是菌群多样性下降、关键代谢菌减少或致病菌增殖,另一端则是色氨酸流向偏移、保护性代谢物不足或神经毒性代谢物累积。
饮食模式变化、抗生素使用、感染、压力等多种因素,都可能通过扰动肠道微生态和色氨酸代谢,推动疾病的发生与进展。这一视角为从源头干预肠道微生物群、重构肠脑轴稳态提供了理论支撑。
未来,围绕这三者间的关系仍有大量问题有待解答。需要进一步解析不同菌群在色氨酸代谢网络中的分工与协同,明确哪些代谢通路、哪些受体信号是真正决定肠脑轴“方向”的关键节点;需要通过严谨设计的临床试验评估饮食干预、益生菌制剂、粪菌移植及新型靶向技术在神经精神结局上的真实获益与适用人群;更需要在此基础上构建以肠道菌群特征和色氨酸代谢谱为核心的分层管理与预测体系,推动从“经验调节微生态”走向“精准重塑肠脑轴”。
随着基础研究与转化医学的不断推进,以肠道微生物群‑肠脑轴‑色氨酸代谢为靶点,有望成为未来干预多类神经及代谢相关疾病的关键突破口。
注:本账号内容仅供学习和交流,不构成任何形式的医疗建议。
主要参考文献
Yuxuan Xia, Chuan Zhang, Leilei Yu, Qingsong Zhang, Arjan Narbad, Wei Chen, Qixiao Zhai, Fengwei Tian,Tryptophan metabolism and the gut-brain axis: focus on specific gut microbial genera,Journal of Future Foods,Volume 6, Issue 5,2026,Pages 740-752,ISSN 2772-5669,https://doi.org/10.1016/j.jfutfo.2024.09.006.
Seo SK, Kwon B. Immune regulation through tryptophan metabolism. Exp Mol Med. 2023 Jul;55(7):1371-1379.
Roager HM, Licht TR. Microbial tryptophan catabolites in health and disease. Nat Commun. 2018 Aug 17;9(1):3294.
Agus A, Planchais J, Sokol H. Gut Microbiota Regulation of Tryptophan Metabolism in Health and Disease. Cell Host Microbe. 2018 Jun 13;23(6):716-724.
Kundi ZM, Lee JC, Pihlajamäki J, Chan CB, Leung KS, So SSY, Nordlund E, Kolehmainen M, El-Nezami H. Dietary Fiber from Oat and Rye Brans Ameliorate Western Diet-Induced Body Weight Gain and Hepatic Inflammation by the Modulation of Short-Chain Fatty Acids, Bile Acids, and Tryptophan Metabolism. Mol Nutr Food Res. 2021 Jan;65(1):e1900580.
Mörkl S, Butler MI, Holl A, Cryan JF, Dinan TG. Probiotics and the Microbiota-Gut-Brain Axis: Focus on Psychiatry. Curr Nutr Rep. 2020 Sep;9(3):171-182. doi: 10.1007/s13668-020-00313-5. Erratum in: Curr Nutr Rep. 2020 Sep;9(3):183.
Yano JM, Yu K, Donaldson GP, Shastri GG, Ann P, Ma L, Nagler CR, Ismagilov RF, Mazmanian SK, Hsiao EY. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell. 2015 Apr 9;161(2):264-76.
Zhang Y, Fan Q, Hou Y, Zhang X, Yin Z, Cai X, Wei W, Wang J, He D, Wang G, Yuan Y, Hao H, Zheng X. Bacteroides species differentially modulate depression-like behavior via gut-brain metabolic signaling. Brain Behav Immun. 2022 May;102:11-22.

谷禾健康
明明滴酒未沾,却突然出现醉酒症状:说话含糊、步态不稳、意识模糊,严重者甚至被交警查出酒驾。
这听起来像天方夜谭,但在医学上,这是一种真实存在的罕见疾病——自酿综合征(Auto-Brewery Syndrome, ABS),也称为肠道发酵综合征。
其核心特征是:在未摄入外源性酒精的情况下,肠道微生物异常发酵碳水化合物,产生并吸收过量乙醇,使血液/呼气酒精水平可达到甚至超过法定阈值,从而出现类似醉酒的一系列神经与胃肠道症状。由于临床表现与饮酒高度相似,自酿综合征在现实中常被误解为酒精依赖、精神问题或刻意隐瞒饮酒史,患者往往经历漫长的误诊与社会压力,甚至常面临法律纠纷。
研究显示,70%的自酿综合征患者报告曾被朋友或家人怀疑撒谎,40%因此失去工作。提高公众和医疗界对ABS的认知,是改善患者生活质量的关键。
近期发表在《Nature Microbiology》的一项研究,通过对22名ABS患者和21名健康家庭成员的系统分析,首次揭示了这种罕见病的微生物学机制,为我们打开了一扇理解肠道菌群与人体健康关系的新窗口。
编辑
本文从自酿综合征的临床表现切入,结合最新《Nature Microbiology》上发表的研究队列从“菌群组成—功能通路—代谢物—体外发酵验证”四个层面系统呈现 ABS 的微生物学机制:发作期以变形菌门(肠杆菌科)异常富集为特征,血液酒精浓度与其相对丰度正相关;宏基因组功能注释显示混合酸发酵、异型乳酸发酵及乙醇胺利用等产醇相关通路显著增强,代谢组学提示乙酸显著升高并与醉酒程度关联。
在此基础上,本文进一步梳理易感人群与诱因、诊断思路与鉴别要点,并结合宏基因组与代谢证据说明产乙醇菌群的形成与维持机制;鉴于常规干预在部分患者中疗效有限且复发风险较高,本文还引入 FMT 个案,作为微生态“重置”策略的临床补充证据,为理解菌群干预与症状逆转之间的关联提供线索。
编辑
从未喝酒,为何会醉酒?
自酿综合征(简称ABS)是一种由于肠道微生物异常发酵碳水化合物,产生过量乙醇(也就是酒精)而导致的罕见代谢紊乱疾病。
患者在没有摄入任何酒精的情况下,血液酒精浓度可达到甚至超过法定酒驾标准(80 mg/dL),出现典型的醉酒症状。这种疾病最早于1948年由Ladkin和Davies首次报道,当时一名5岁非洲男孩因肠道异常发酵导致严重腹胀和穿孔死亡。
在日本,这种疾病被称为”meitei-sho”(肠内酒精发酵综合征),并在20世纪70年代有过系列报道。而在现代医学中,自酿综合征仍然常常被误诊为酒精依赖或精神疾病,许多患者在确诊前往往经历数年的误诊和社会误解。
患者的真实困境:从酒鬼到患者
Malik等人在2019年报道了一个典型案例:一名46岁男性在接受头孢类抗生素治疗拇指外伤后,出现性格改变、抑郁和脑雾症状。一次清晨,他因疑似酒驾被捕,血液酒精浓度高达200 mg/dL,但他坚称未饮酒。经过多次辗转,最终通过粪便检测发现大量酿酒酵母(Saccharomyces cerevisiae),确诊为自酿综合征(ABS)。
“最令人心碎的是,连医生和警察都不相信我没有喝酒。我被贴上了酒鬼的标签,失去了工作和朋友的信任。直到确诊ABS,我才终于洗清了冤屈。”
—— ABS患者自述
这种疾病不仅带来身体上的痛苦,更带来严重的社会和法律问题。据统计,已有多起ABS患者因酒驾被起诉后,通过医学证据最终被判无罪的案例。
如果说自酿综合征(ABS)是一个没喝酒也会醉的怪病,那么它的本质,其实就是肠道微生态,被重塑成了一座高效的小型酿酒厂。这一章节,我们就沿着最新《Nature Microbiology》队列研究的证据,看看这座酿酒厂是如何一步步搭建起来的。
自酿综合征:从正常发酵到病理酿酒
正常情况下,肠道菌群处于动态平衡状态,少量乙醇的产生会被肝脏迅速代谢。但在ABS患者中,这种平衡被打破,特定微生物过度生长,将摄入的碳水化合物大量转化为乙醇。
而在 ABS 患者身上,发生了三件关键的事:
1
菌群结构改变
原本占主导的共生菌减少,某些能高效产乙醇的细菌占位。
2
代谢通路重编程
多条能把糖和乙醇胺酿成酒的通路被全面激活。
3
代谢产物堆积,并进入血液
乙醇与相关代谢物(尤其是乙酸)在肠道内大量生成、被吸收,超出肝脏清除能力,如果超过一定量,甚至会诱发系统性中毒。
下面我们分步骤拆解这三层变化。
菌群失衡:谁在主导酿酒?
变形菌门(Proteobacteria)显著富集
在这项纳入 22 名经严格口服葡萄糖激发试验证实的 ABS 患者、21 名家庭伴侣和一组健康对照的研究中,宏基因组测序有一个醒目的共同点:
编辑
编辑
⭐️ 三个关键的酿酒菌(产乙醇)
多变量分析(校正家庭环境影响)指出了三种重要的菌群:
大肠杆菌(Escherichia coli)
肺炎克雷伯菌(Klebsiella pneumoniae)
Ruminococcus gnavus
⭐️ 有益菌下降
编辑
总的来说,ABS患者整个微生态从多样而稳定,滑向“低多样性 + 病理性富集”的状态。
产乙醇代谢通路拉满
如果说上述菌群失衡回答了谁在干,那代谢通路分析则是回答了他们在干什么。
研究利用宏基因组功能注释,对代谢通路进行比对,发现:
其中,与乙醇生产直接相关、在发作期明显富集的关键通路包括:
编辑
⭐️ 混合酸发酵通路(Mixed‑acid fermentation)
编辑
⭐️ 异型乳酸发酵通路(Heterolactic fermentation)
⭐️ 乙醇胺利用通路(Ethanolamine utilization)
乙醇胺来自于肠道上皮细胞膜磷脂的分解,是一种内源性氮源和碳源。一些细菌可以通过专门的乙醇胺利用操纵子,把乙醇胺转化为乙醛、乙酰‑CoA,最终还原为乙醇。
在 ABS 发作期:
编辑
⭐️ 基因水平与血液酒精浓度高度相关
研究者发现,所有与酒精脱氢酶活性相关基因总丰度,与患者的血液酒精浓度高度相关(Spearman R = 0.72,P=2.7×10⁻⁶)。
编辑
这就把微生物基因层面与临床醉酒程度直接关联起来,不是只有菌在那儿摆着,而是在真正参与乙醇的大量生产。
体外实验证实:这些菌真的在酿酒
为了避免一切只是推测,研究团队还做了一个非常关键的实验——体外粪便厌氧培养测乙醇。
取 100 mg 粪便,在含葡萄糖的厌氧培养基中培养 24 小时,并在 0、6、24 小时测定培养液中的乙醇浓度(HPLC/酶法)。结果显示:
ABS 发作期样本:中位乙醇浓度约 14.47 mg/dL;
同一患者缓解期样本:约 8.76 mg/dL;
家庭伴侣样本:约 5.00 mg/dL。
ABS 发作期(flare)样本的体外产乙醇能力更强,并且与采样时的血液酒精浓度相关。
关键菌群:细菌而不是真菌
自 ABS 被提出以来,很多早期病例把矛头指向肠道酵母过度生长,临床上因此常常首选抗真菌药物治疗。但这项队列研究给出了更细致的答案。
在体外培养体系中加入不同药物:
广谱抗生素——氯霉素
广谱抗真菌药——两性霉素B
代谢产物:乙酸是ABS的生物标志物
除乙醇外,ABS患者的粪便代谢组学分析显示,ABS 发作期患者的粪便乙酸水平显著高于家庭伴侣(P=1.2×10⁻⁵),且与血液酒精浓度呈正相关(R=0.6, P=0.00018)。
乙酸是乙醇代谢的中间产物,同时也可作为底物被某些细菌用于乙醇合成,形成乙醇-乙酸循环,这可能是ABS患者乙醇持续产生的另一重要原因。
编辑
在菌群–代谢物相关性分析中:
乙酸水平与 Escherichia、Blautia 等菌属呈正相关;与 Akkermansia 等典型有益菌负相关。
也就是说,比起频繁做口服葡萄糖激发试验,监测粪便乙酸或许在随访中更具可行性。
抗生素
多项研究表明,抗生素使用是ABS常见的诱因。抗生素会破坏肠道正常菌群平衡,导致产乙醇菌过度生长。
在Malik报道的案例中,患者在使用头孢氨苄(cephalexin)3周后出现ABS症状。
另一项研究发现,长期使用阿莫西林-克拉维酸、甲硝唑等广谱抗生素的患者,肠道变形菌门比例显著增加,ABS风险提。
基础疾病:肠道动力障碍与吸收不良
ABS常与以下基础疾病相关:
短肠综合征
肠道切除后,食物通过加快,碳水化合物未充分吸收即进入结肠,成为产乙醇菌的底物。
克罗恩病
肠道炎症和狭窄导致食物滞留,促进细菌过度生长。
糖尿病
高血糖环境促进酵母和某些细菌生长,研究显示糖尿病患者内源性乙醇水平显著高于健康人。
胃轻瘫
胃排空延迟导致食物在胃肠道停留时间延长,增加发酵机会。
饮食因素:吃太多碳水化合物
饮食中的碳水化合物是肠道菌群产乙醇的原料。研究发现,ABS患者在摄入高碳水化合物食物(如面包、 pasta、含糖饮料)后,症状明显加重,血液酒精浓度可在2-8小时内达到峰值。
一项病例报告显示,一名ABS患者在食用披萨和苏打水后,血液酒精浓度迅速升至400 mg/dL,出现严重醉酒症状。
遗传因素:酒精代谢酶的个体差异
酒精脱氢酶(ADH)和乙醛脱氢酶(ALDH)的基因多态性可能影响ABS的临床表现。
亚洲人群中常见的ALDH2*2突变(导致乙醛脱氢酶活性降低),可能使ABS患者更容易出现乙醛蓄积,加重醉酒症状。
警惕这些不寻常的醉酒症状
ABS的临床表现与普通醉酒相似,但具有以下特点:
关键诊断试验:碳水化合物激发试验
碳水化合物激发试验具体步骤如下:
1. 患者需禁酒48小时,空腹8小时
2. 基线血液酒精浓度和尿酒精代谢物(乙基葡萄糖醛酸苷、乙基硫酸酯)检测
3. 口服100-200克葡萄糖(或标准化碳水化合物餐)
4. 在0.5、1、2、4、8、12、24小时监测血液酒精浓度。
5. 若任何时间点血液酒精浓度≥0.01 g/dL(10 mg/dL),且排除外源性酒精摄入,则可诊断ABS。
注意:该试验需在严格医疗监督下进行,因为部分患者可能出现严重醉酒,甚至酒精中毒。
微生物检测
通过肠道菌群检测,可以从产乙醇相关菌群(如大肠杆菌、肺炎克雷伯菌)是否超标,以及相关代谢通路是否异常,来辅助判别是否存在自酿综合征的可能。
排除其他疾病
ABS需与以下疾病鉴别:
-酒精依赖
患者通常隐瞒饮酒史,尿酒精代谢物检测阳性
-肝性脑病
肝功能异常,血氨升高,无肠道产乙醇证据
-糖尿病酮症酸中毒
血糖显著升高,尿酮体阳性
-药物中毒
如苯二氮䓬类、阿片类药物过量
-罕见神经系统疾病
如发作性共济失调、卟啉病
饮食干预:减少酿酒原料
低碳水化合物饮食是ABS的基础治疗。具体建议包括:
一项病例系列研究显示,80%的ABS患者在严格低碳水饮食(每日碳水化合物<50克)6周后,症状显著改善,血液酒精浓度恢复正常。
抗生素治疗
根据微生物检测结果选择针对性药物,例如利福昔明、甲硝唑等。需注意,抗生素可能进一步破坏肠道菌群平衡,因此通常仅在严重病例中短期使用,可以考虑同时联合益生菌治疗。
益生菌调节
益生菌可通过竞争营养和黏附位点,抑制产乙醇菌生长。研究显示,Lactobacillus acidophilus、Bifidobacterium infantis等菌株可降低ABS患者肠道pH值,减少乙醇产生。
一项病例报告显示,一名ABS患者在使用多菌株益生菌(含12种细菌)1.5年后,症状缓解,可正常饮食。
长期管理:预防复发的关键
ABS治疗后复发率较高,长期管理需注意:
虽然前述多种常规治疗手段(包括抗生素、限制碳水、益生菌等)能在部分病例短期内缓解 ABS 症状,但对病情较重或反复发作的病例而言,疗效往往不够理想,难以在短时间内达到预期的控制目标。
对于难治性ABS,粪菌移植(FMT)可能是一种值得考虑的选择。该研究报道了一例ABS患者接受多轮FMT并长期随访的案例。这不仅丰富了我们对ABS发生机制的认识,也为探索菌群干预在临床治疗中的应用潜力提供了重要参考。
因此,我们将在下一章对 FMT 的这个案例进行专门介绍。
这个FMT案例是围绕单个患者的多轮 FMT、长期随访与宏基因组分型,系统性地提供了微生态干预下 ABS 逆转的因果证据。
粪菌移植干预过程和结果
患者背景
第一次 FMT:暂时好转
在 FDA 单例扩展使用批准下,接受:
结果:
第二次FMT
在首次FMT后9个月接受了第二次FMT。
采样时间线
编辑
这次口服万古霉素、甲硝唑和复方新诺明预处理3天,随后进行肠道准备。
注:
文中对该方案的解释:第一次用利福昔明与新霉素主要为肠腔内抗生素,全身吸收很少;
而第二次加入的甲硝唑与复方新诺明为可全身吸收抗生素,旨在不仅作用于肠腔内微生物,也可覆盖肠腔外/黏膜相关部位,包括绒毛刷状缘与肠隐窝等微生物,可能以生物膜或低代谢状态持续存在的部位。
给药与维持:预处理后先给 3 次、每次 15 粒胶囊;随后在不再预处理的情况下,用同一供者来源胶囊进行每月维持 15 粒,持续 6 个月。
辅助措施(促进定植):
同时嘱咐每日服用一大汤匙马铃薯淀粉,约 60% 抗性淀粉,溶于水;
复合益生菌每日 1 粒,含 Bifidobacterium infantis、Clostridium butyricum、Clostridium beijerinckii、Anaerobutyricum hallii、Akkermansia muciniphila,以促进移植菌群的持续存在/定植。
结果:
肠道菌群变化与临床关联
粪菌移植后菌群结构变化
第一次 FMT 后的缓解期样本,在主成分图上与供体的菌群组成聚在一起;
症状复发时,样本又跑回类似初始发作期的菌群结构。
第二次 FMT 后,菌群持续稳定且不同于术前/家庭成员菌群。
编辑
主成分贡献最大的菌
编辑
发作期 E. coli 丰度高;K. pneumoniae、R. gnavus 未显著增加。
编辑
FMT后大肠杆菌丰度、血液酒精浓度和发酵通路基因显著降低。
代谢和临床指标相关性
菌群应答、血乙醇浓度、AST/ALT、产醇相关通路丰度紧密相关。
编辑
同一时间轴上同时给出 BAC(血/呼气酒精)、AST/ALT、以及 fermentation pathway enrichment(混合酸发酵/异乳酸发酵/乙醇胺利用等的富集指标),并与菌群变化/发作缓解标注对应。
症状与供者菌株植入率正相关;与术前菌株保留(post-FMT 与 pre-FMT 共享比例)负相关。
编辑
总的来说,对这位患者而言,ABS 的发生与一个特定高产乙醇耐乙醇的大肠杆菌菌株群落密切相关;通过 FMT + 系统性抗生素预处理,成功完成菌群置换,症状得以长期缓解。
自酿综合征(ABS)表面上是一种罕见而离奇的疾病,却在机制、诊断与干预的层层展开中,清晰指向同一个核心事实:肠道菌群具备强大的代谢能力,其结构与功能的失衡,足以在人体内酿出可被检测、可致症状的乙醇水平。
面向未来,ABS 的意义并不止于罕见病本身。研究提示,即便在非 ABS 人群中也可能存在低水平内源性乙醇产生,这一现象或与非酒精性脂肪肝、肥胖等代谢异常相关;例如,肺炎克雷伯菌产生的乙醇可诱导动物出现肝脏脂肪变性。由此,ABS 也可能成为理解“菌群—代谢疾病”连接机制的窗口,并为常见代谢病提供新的干预靶点。
治疗层面,ABS 有望走向更具针对性的精准医疗路径:
与此同时,提高诊断率同样关键——ABS 的真实患病率可能被低估(有研究估计每 10 万人中或有 1–2 例),推动临床认知、完善流程并探索更便捷的检测手段(如肠道菌群检测),将有助于患者更早被识别、更少陷入误解与法律困境。
当肠道菌群喝醉,我们需要做的不仅是控制血液酒精和缓解症状,更是把 ABS 放回宿主—微生物共生系统的框架中重新理解:这并非人体的例外,而是微生物代谢能力在特定条件下的极端呈现。
通过提高认知、规范诊断与发展靶向菌群治疗,我们或许不仅能更好地帮助 ABS 患者,也能借此进一步理解菌群在代谢健康中的位置与边界。
在这个由 100 万亿微生物组成的“超级器官”面前,我们对生命的理解或许才刚刚开始。
主要参考文献
Hsu CL, Shukla S, Freund L, Chou AC, Yang Y, Bruellman R, Raya Tonetti F, Cabré N, Mayo S, Lim HG, Magallan V, Cordell BJ, Lang S, Demir M, Stärkel P, Llorente C, Palsson BO, Mandyam C, Boland BS, Hohmann E, Schnabl B. Gut microbial ethanol metabolism contributes to auto-brewery syndrome in an observational cohort. Nat Microbiol. 2026 Jan 8.
Dinis-Oliveira RJ. The Auto-Brewery Syndrome: A Perfect Metabolic “Storm” with Clinical and Forensic Implications. J Clin Med. 2021 Oct 10;10(20):4637.
Malik F, Wickremesinghe P, Saverimuttu J. Case report and literature review of auto-brewery syndrome: probably an underdiagnosed medical condition. BMJ Open Gastroenterol. 2019 Aug 5;6(1):e000325.
Xue G, Feng J, Zhang R, Du B, Sun Y, Liu S, Yan C, Liu X, Du S, Feng Y, Cui J, Gan L, Zhao H, Fan Z, Cui X, Xu Z, Fu T, Li C, Huang L, Zhang T, Wang J, Yang R, Yuan J. Three Klebsiella species as potential pathobionts generating endogenous ethanol in a clinical cohort of patients with auto-brewery syndrome: a case control study. EBioMedicine. 2023 May;91:104560.
Tameez Ud Din A, Alam F, Tameez-Ud-Din A, Chaudhary FMD. Auto-Brewery Syndrome: A Clinical Dilemma. Cureus. 2020 Oct 16;12(10):e10983.

谷禾健康
Akkermansia muciniphila(AKK菌)因其独特的黏蛋白降解能力和与宿主健康的复杂关系,目前已成为微生物学和医学研究的前沿热点。谷禾此前已对该菌进行过系统性介绍。
肠道重要菌属——Akkermansia Muciniphila,它如何保护肠道健康
本文将继续深入探讨AKK菌的研究前沿。
2004年,当这株微小的厌氧菌首次从人类粪便中被分离出来时,没人预料到它会凭借独特的黏蛋白降解能力(当然目前又陆续发现了几个黏蛋白降解菌),在此后近二十年间掀起一波又一波的研究热潮。
更引人关注的是AKK菌呈现出耐人寻味的双面性:它既是下一代益生菌的希望之星,与代谢健康、长寿紧密相连;又在某些特定情境下显现出促进疾病的潜在风险。这种复杂的功能属性,正是当前科研亟待厘清的关键议题。
三篇发表于顶级期刊的AKK菌研究为该领域带来了重要突破:
这三篇文献从不同维度构建了迄今为止最为全面和深入的AKK菌研究图景,涵盖了其基本生物学特性、复杂的基因组结构与菌株多样性、与宿主免疫系统的精密互作机制,以及在不同疾病模型中呈现的矛盾表型,系统解析了这些现象背后的科学逻辑。
因此,本文谷禾整合了这三篇文献的核心观点,以科研从业者的视角带领大家深入解构AKK菌的研究进展。我们希望与各位共同探讨一个关键问题:如何理性认知并合理应用AKK菌这个双面细菌,从而真正实现精准调控肠道健康的目标。
嗜黏蛋白阿克曼氏菌(Akkermansia muciniphila),简称为AKK菌,属于疣微菌门。这个名字是为了纪念荷兰微生物生态学家Antoon Akkermans博士,而“muciniphila”则源自拉丁语,意为偏爱黏蛋白。
注:Antoon Akkermans:荷兰瓦赫宁根大学(Wageningen University)微生物生态学家,对土壤和肠道微生物研究有重要贡献。
AKK菌的发现和命名具有里程碑意义
2004年,仅使用黏蛋白(mucin)作为唯一的碳源和氮源进行富集培养了该菌。AKK菌具备独特的酶系统,能够降解并利用黏蛋白作为其生存和生长的主要能量来源,这种自给自足的能力使其在竞争激烈的肠道环境中占据了稳定的一席之地。
其模式菌株为Akkermansia muciniphila MucT(亦写作Muc5),拥有多个菌株编号,如ATCC BAA-835、DSM 22959等,是目前研究中应用最广泛的菌株。
生态位
AKK菌在人体肠道中占据着一个非凡的生态位——肠道黏液层。这一层由宿主杯状细胞分泌的黏蛋白糖蛋白构成,是隔开肠道上皮细胞与肠腔内大量微生物的第一道物理和化学屏障。
从分布来看,AKK菌不仅存在于人类肠道,也广泛栖息于包括小鼠、牛、猪、兔等多种脊椎动物的胃肠道。
形态与基本特性
在显微镜下,AKK菌呈现为一种椭圆形、不运动、不产芽孢的革兰氏阴性严格厌氧菌。
AKK菌并非绝对厌氧
研究发现,在只有微量氧气的环境中,它不仅能存活,甚至可能生长得更好。这主要得益于其基因组编码的细胞色素 bd 氧化酶复合物等耐氧机制。
也正因为具备这种有限耐氧能力,AKK菌才能适应并定植于结肠黏液层——这里处在厌氧的肠腔与相对更富氧的上皮细胞之间,存在明显的氧梯度。
结构特征,驱动互作
AKK菌约2.66Mb的基因组,是其特殊生活方式的说明书。其中最引人注目的,就是黏蛋白利用基因簇(Mucin Utilization Loci, MULs)。
黏蛋白转运系统:把食物带回家再吃的策略
Grant等人的研究指出,这些基因编码了一套精密的黏蛋白转运系统,能将黏蛋白大分子吞入菌体内部一个叫黏蛋白体的特殊结构中进行降解。这种偏自私的代谢策略,有点像把食物带回家再吃,从而最大限度地减少了与肠道中其他微生物的营养竞争,进而巩固自身的生存优势。
编辑
此外,它的细胞表面也布满了武器。电镜观察到的菌毛样结构,特别是重要的外膜蛋白Amuc_1100,不仅参与黏附,更是与宿主免疫系统直接对话的关键分子。
其细胞壁的肽聚糖层含有非乙酰化的葡糖胺残基,这在革兰氏阴性菌中相当罕见,使其能被宿主的NOD1和NOD2受体识别,从而触发免疫应答。这些独特的结构,都为它与宿主之间复杂的相互作用埋下了伏笔。
生态分布:影响因素和趋势
AKK菌在人群中的分布呈现出鲜明的特征
根据人类肠道微生物组图谱对来自20个国家3268名健康人的数据分析,AKK菌在约40.3%的健康供体中被检测到,平均相对丰度为1.24%。这个数字看似不高,但其分布的倾向性却极具信息量。
AKK菌的分布图谱
编辑
Grant et al., Nature Microbiology
a) 在健康人群中,其丰度和患病率存在地理和性别差异。
b) 在不同疾病状态下,其丰度呈现显著变化,例如在IBD中减少,而在帕金森病中富集。
c) 其相对丰度随生命周期动态变化,在百岁老人中再次出现高峰。
d) 进化树揭示了AKK菌属内部复杂的系统发育关系。
从图1中我们可以清晰地看到:
-地域与生活方式
工业化人群中的丰度更高,这暗示着饮食或生活方式可能是其丰度的重要调节因素。
-年龄
AKK菌在婴儿期迅速定植,成年后逐渐下降,但在百岁老人中却意外地再次富集,被认为是健康长寿的潜在标志物。Luo等人的文章也系统总结了这一现象,指出AKK菌在人类中的年龄依赖性定植模式与在小鼠中的模式恰好相反,这提醒我们在选择动物模型时需格外谨慎。
-疾病状态
这是最能体现其双面性的一点。在炎症性肠病(IBD)患者中,AKK菌丰度显著降低(UC患者中为9.9%,克罗恩病患者中为14.3%);然而,在帕金森病(90.3%)、某些癌症(如黑色素瘤,65.9%)和动脉粥样硬化(83.3%)患者中,其丰度却异常增高。这种看似矛盾的分布模式,强烈暗示AKK菌的角色并非简单的好或坏,而是深度依赖于宿主的病理生理背景。
Tips:看到这些分布数据,首先想到的不是简单的因果关系,而是一个复杂的生态反馈回路。例如,在IBD中,AKK菌的减少可能是肠道炎症环境恶化、黏液层破坏的结果,而非原因。反之,在帕金森病中,其增多也可能是一种代偿性反应,或是神经退行性疾病引发的肠道环境改变所致。将AKK菌丰度作为疾病的生物标志物时,我们必须高度审慎,通过系统证据加以判别:该因素究竟是事件升级的关键驱动(促进因素),是对事态的响应性干预(缓解因素),还是与结果无显著因果关联的伴随出现(旁观变量)。
所以构建真实世界的特定疾病样本人群数据才能更有助于解析复杂的肠道微生态,类似的菌还有如活泼瘤胃球菌和普雷沃氏菌等相互矛盾的研究。
三篇文献都不约而同地强调了菌株水平多样性的重要性。
长久以来,大多数研究都围绕着模式菌株MucT(ATCC BAA-835)展开。然而,随着基因组学技术的发展,AKK菌家族的内部复杂性逐渐浮出水面。基于全基因组测序,研究人员已将AKK菌划分为至少四个系统发育群(phylogroups, AmI-AmIV)。
这些不同发育群的菌株虽然16S rRNA基因序列高度相似(>;99%),但其全基因组的平均核苷酸同一性(ANI)却可能低于95%,这已经达到了物种划分的界限。这意味着,我们过去所称的“A. muciniphila”很可能是一个包含多个物种或亚种的复合体。这种基因组上的差异,直接导致了功能上的多样性:
代谢能力差异
Loannou等人提到,AmII发育群的菌株拥有合成维生素B12的能力,而AmI发育群则不具备,这直接影响了它们的代谢产物谱。不同菌株对人类母乳寡糖(HMOs)的利用效率也存在显著差异。
对宿主影响的差异
在DSS诱导的结肠炎小鼠模型中,研究人员发现不同的人源AKK菌株对肠道炎症的影响截然不同:一个菌株表现出保护作用,两个菌株没有效果,而第四个菌株甚至有加剧炎症的趋势。这为我们敲响了警钟:随意使用一种AKK菌株来治疗IBD,可能不仅无效,甚至有害。
抗生素抗性差异
模式菌株MucT携带多种抗生素抗性基因(如blaA, dfrA, sul, tetM, van)。但从健康人中分离的菌株,其抗性谱各不相同。考虑到AKK菌具有通过水平基因转移获取新基因的能力,将一个未经充分安全性评估的活菌株作为益生菌推向市场,存在传播抗生素抗性的潜在风险。
对于产业界而言,开发AKK菌产品时,可能要对菌株进行全面的功能和安全性评估(包括代谢能力、免疫调节特性、抗生素抗性谱、基因转移能力等)是不可或缺的关键步骤。
AKK菌能在竞争激烈的肠道环境中占据一席之地,并与宿主展开如此复杂的对话,其背后是一套精妙的分子机制。这不仅是它生存的智慧,也是我们理解其双面性的钥匙。
黏蛋白降解
AKK菌对黏蛋白的降解,远非简单的啃食。它拥有一整套工具箱——即种类繁多的碳水化合物活性酶(CAZymes)。这个过程极具策略性:
-外层突破
首先,通过表面的唾液酸酶和岩藻糖苷酶,切除黏蛋白聚糖链最外层的唾液酸和岩藻糖残基。
-核心瓦解
接着,动用半乳糖苷酶、己糖胺酶和硫酸酯酶等,逐步分解聚糖核心结构。
-内部消化
降解产物通过糖转运系统高效内化。AKK菌虽不具备拟杆菌式的典型PULs/MULs基因簇,但其膜转运蛋白能有效摄取降解后的单糖和寡糖,进行发酵,最终产生乙酸、丙酸等短链脂肪酸(SCFAs)。
注:
编辑
其中,丙酸的产生尤为重要。它不仅是其他肠道菌的能量来源,还能通过激活G蛋白偶联受体GPR41和GPR43,刺激肠道L细胞分泌胰高血糖素样肽-1(GLP-1),从而参与调节宿主的血糖稳态和食欲。这部分解释了AKK菌在代谢性疾病中的有益作用。
与免疫系统的多渠道对话
AKK菌与宿主免疫系统的互作,是一场多层次、多渠道的复杂对话。Grant等人的文章中这个信息量大的图,为我们生动地展示了这一过程。
AKK菌与宿主结肠黏膜的相互作用机制示意图
编辑
Grant et al., Nature Microbiology
有几个关键的互作途径:
编辑
图源:doi.org/10.1038/s41579-024-01106-1
AKK菌并非孤立地存在于肠道中,而是作为复杂微生物网络的一部分,与其他细菌、古菌甚至病毒发生着密切的相互作用。这些相互作用包括协同合作、竞争排斥和营养交换(交叉喂养),共同决定了肠道微生态的结构、功能以及对宿主健康的影响。
交叉喂养:生态系统中的资源分享者
尽管AKK菌被描述为自私的黏蛋白降解者,因为它倾向于将黏蛋白内化代谢,但这一过程实际上为其他微生物创造了丰富的资源。
为产丁酸菌提供底物
AKK菌降解黏蛋白释放的单糖(如岩藻糖)和产生的代谢产物(如乙酸盐、丙酸盐),可以被许多重要的产丁酸菌利用。
例如,在共培养实验中,AKK菌能支持产丁酸菌如Anaerostipes caccae、Anaerobutyricum hallii、Faecalibacterium prausnitzii、Roseburia等细菌的生长,并促进有益代谢物丁酸盐的产生。丁酸盐是结肠上皮细胞的主要能源,具有强大的抗炎和维持肠道屏障完整性的功能。
编辑
图源:doi.org/10.1038/s41579-024-01106-1
双向营养交换
AKK菌与其他细菌的相互作用可以是双向的。例如,它与A. hallii之间存在互惠共生关系:AKK菌为A. hallii提供黏蛋白降解产物和乙酸用于丁酸合成,而A. hallii产生的丁酸等代谢物有助于维持适宜AKK菌生长的肠道微环境。
某些研究还提示可能存在更直接的代谢物交换,如维生素或辅因子的共享,但具体机制仍在探索中。
协同作用:共同抵御疾病
A. muciniphila 可能需要其他微生物的存在来预防某些疾病。
1+1>2:微生物协同互作的抗病潜力
例如,在结直肠癌(CRC)小鼠模型 Apc 突变小鼠中,单独定植 A. muciniphila 或幽门螺杆菌导致肿瘤负担增加,而两者共定植则减少了肠道肿瘤数量。
A. muciniphila Muc 和狄氏副拟杆菌(Parabacteroides distasonis)表现出协同的抗结肠炎关系。
在癫痫 Kcna1⁻/⁻ 小鼠中,A. muciniphila Muc和 狄氏副拟杆菌的联合给药通过降低细菌交叉代谢产生的γ-谷氨酰转肽酶活性,在控制饮食条件下预防了癫痫发作。
这些发现强调了一个核心观点:微生物之间的相互作用网络,比单一菌株的存在与否更能决定疾病的结局。
编辑
图源:doi.org/10.1038/s41579-024-01106-1
不单单是AKK菌,而应评估菌群的整体效应
这种复杂的互作关系提示我们在应用层面需要更宏观的视野。
通过菌株脱除试验发现, A. muciniphila 的引入,与抗炎共生菌普拉梭菌(F. prausnitzii)丰度降低相关(在疾病诱导之前),对柠檬酸杆菌(C. rodentium)感染的易感性部分归因于这种细菌网络的变化。这意味着,即使 AKK 菌无法长期定植,它作为过客仍可能重塑肠道菌群的功能结构。
因此,未来评估益生菌应用时,不能仅盯着 AKK 菌本身,而应将其视为生态系统的一个扰动因子,充分重视个体间菌群基线的差异,深入评估其对肠道微生态结构和功能的长期重塑作用,以及宿主原有菌群对其行为的反向调控。
竞争与拮抗
在肠道有限的生态位中,竞争是不可避免的。
与黏液降解菌的竞争
在以MUC2为唯一碳源的体外共培养体系中,当AKK菌与其他的黏液降解菌如普通拟杆菌(Bacteroides vulgatus)、活泼瘤胃球菌(Ruminococcus gnavus)等一起培养时,AKK菌的生长会受到抑制,而其他细菌的生长则被促进。这表明在黏液降解这一功能上存在激烈的竞争。
对其他菌群的负向影响
在某些情况下,AKK菌的存在可能对其他有益菌产生负面影响。
例如,在一个定义的微生物群落中,AKK菌的存在与抗炎共生菌Faecalibacterium prausnitzii丰度的降低相关。但这种相关性并不一定代表因果关系——可能是:环境条件改变导致两者同时变化,或其他因素的间接影响特定疾病状态的反映(如炎症性肠病)。
与普雷沃氏菌的负相关
在人类肠型(enterotype)研究中,AKK菌通常在以Ruminococcus属为主的肠型中富集,而与以普雷沃氏菌为主的肠型呈负相关,这反映的是菌群组成的自然差异,不同肠型由遗传、饮食、地理位置等多因素决定,不是AKK菌排斥普雷沃氏菌,而是不同的生态位和代谢特征。
目前最核心、也最令人困惑的部分:AKK菌在不同疾病背景下的双面角色。三篇文献都花费了大量篇幅,尽力为我们揭示了这种环境依赖性。
AKK菌与结肠炎:保护、致病与宿主互作的复杂博弈
关于AKK菌与结肠炎的关系,研究结论充满了矛盾。
保护作用
在常规的DSS化学诱导结肠炎模型中,灌胃AKK菌或其蛋白Amuc_1100,通常能观察到保护效果。其机制包括上调紧密连接蛋白(如ZO-1、Occludin)以修复屏障、减轻内质网应激、促进Treg细胞应答等。Luo等人的综述也证实,活菌能够增加紧密连接蛋白的表达。
致病作用
然而,一旦宿主背景改变,情况就可能逆转。在遗传易感的Il10-/-小鼠(一种自发性结肠炎模型)中,单独定植AKK菌反而会加剧炎症,导致黏液层变薄和促炎细胞因子上调。这表明,在宿主免疫调节能力受损(如缺乏关键的抗炎因子IL-10)的情况下,AKK菌的黏蛋白降解活性可能弊大于利。
宿主蛋白的策反
Grant等人的研究揭示了一个更为精妙的机制。在溃疡性结肠炎患者中,一种名为Intelectin-1(ITLN1)的宿主蛋白会过度表达。这种蛋白会特异性地结合AKK菌,将其拉到更靠近上皮细胞的位置。这种亲密接触在炎症背景下,反而加剧了免疫反应和组织损伤。
感染:启动防御,还是放大风险?
在面对外来病原体入侵时,AKK菌的角色同样摇摆不定。
有益面:抗感染
AKK菌的某些组分或代谢物显示出抗感染的潜力。例如,其产生的三肽RKH能通过阻断TLR4信号通路,保护小鼠免于致死性脓毒症。Amuc_1100蛋白则能预防沙门氏菌引起的肝损伤。
有害面:缺膳食纤维就吃粘液层,病原体侵入
AKK菌的关键功能——降解黏蛋白,在特定条件下会为病原体开门。
在一项设计精巧的无菌小鼠实验中,研究者发现,缺乏膳食纤维的饮食会导致肠道中的AKK菌饥不择食,转而大量消耗黏液层。这层被削弱的物理屏障,使得致病菌Citrobacter rodentium能够轻易入侵,导致致命感染。
更具说服力的是,当研究者从这个菌群中移除AKK菌后,即使在无纤维饮食下,小鼠也能免于感染。这证明了AKK菌在这种情境下的内鬼角色。有趣的是,一旦恢复富含纤维的饮食,AKK菌的存在反而与较低的病原体载量相关,显示出保护作用。
编辑
Tips:同一种菌,两副面孔:有膳食纤维时,AKK菌是肠道卫士;没有纤维时,它却会啃食肠道黏膜。这个营养开关,让它在天使与魔鬼之间切换。这个发现具有极其重要的实践意义。告诉我们,补充AKK菌益生菌的同时,如果忽略了饮食管理(特别是保证充足的膳食纤维摄入),可能不仅无法获益,甚至可能损害肠道屏障,增加感染风险。这为精准营养与精准菌群干预的结合提供了强有力的理论依据。
癌症:抗肿瘤反应与促肿瘤微环境的动态博弈
在癌症领域,尤其是结直肠癌,AKK菌的形象同样复杂。
促癌风险
部分研究在结直肠癌患者的肿瘤组织和相应的小鼠模型中,都观察到了AKK菌的富集。有观点认为,它通过降解黏液屏障,可能为肿瘤的发生发展创造了条件。
抑癌潜力
然而,另一些研究则得出了相反的结论。AKK菌的EVs、乙酰转移酶Amuc_2172等组分,在小鼠模型中显示出抑制肿瘤生长的效果。
在免疫治疗中的助攻
AKK菌在癌症研究中最高光的时刻,莫过于其在免疫检查点抑制剂(如PD-1抗体)治疗中的作用。
多项研究一致发现,对PD-1治疗有反应的癌症患者(包括肺癌、肾癌等),其肠道中AKK菌的基线水平显著更高。更关键的是,在小鼠模型中,将无反应者粪菌移植给小鼠后,再补充AKK菌,能够重新恢复小鼠对PD-1治疗的敏感性。这表明AKK菌可能通过调节全身免疫状态,增强了抗肿瘤免疫应答,从而成为免疫治疗的增效剂。
代谢与神经系统疾病:更偏向有益的角色
相较于在炎症和感染中的摇摆不定,AKK菌在代谢性疾病中的有益作用,是目前证据最为一致、也最具转化潜力的领域。无论是动物模型还是初步的人体研究,补充活菌或巴氏杀菌的AKK菌,都被证明能够改善胰岛素敏感性、降低胆固醇、减少脂肪堆积。其机制与促进GLP-1分泌、调节脂肪酸代谢、减轻低度炎症等密切相关。
在神经系统疾病中,AKK菌的关联性再次变得复杂。它在帕金森病和多发性硬化症患者中常常富集,体外实验也显示它可能诱导α-突触核蛋白聚集或促炎反应。然而,也有研究将多发性硬化症患者中AKK菌的增多与较低的残疾程度联系起来,提示这可能是一种有益的代偿反应。这种矛盾性再次凸显了菌株差异和宿主背景的重要性。
面对AKK菌如此复杂,我们该如何从科学研究走向临床应用?有幸的是三篇文献为我们提供了一些思考和前瞻性的指导。
编辑
Grant et al., Nature Microbiology
影响AKK菌双重性的因素及潜在治疗策略。AKK菌的最终效应受到菌株差异、饮食、微生物互作和宿主状态的共同调节。理解这些变量,有助于我们选择最优的治疗应用方案,例如是使用活菌、灭活菌,还是分离的生物活性组分。
精准应用的策略选择
未来的AKK菌疗法,绝非一招鲜,吃遍天,而应是量体裁衣的精准策略。
活菌 vs. 灭活菌
巴氏杀菌的AKK菌是一个极具吸引力的选择。它在欧盟已被批准为新食品原料,安全性更高,避免了活菌定植和基因转移的风险。
多项研究证实,灭活菌依然保留了大部分有益的代谢调节功能,这可能归功于其热稳定的细胞壁成分(如Amuc_1100)。
整体 vs. 组分
直接使用分离的生物活性组分,是更为精准和安全的策略。例如,将Amuc_1100蛋白作为药物开发,或利用其EVs作为治疗载体。
Ioannou 等人的综述强调了 AKK 菌作为“酶学底盘”的巨大潜力。例如利用AKK菌的糖苷酶来改造血细胞表面的ABO血型抗原,展示了从基础研究到生物技术应用的转化潜力。
补充 vs. 内源调节
除了直接补充AKK菌,通过饮食干预(如补充富含多酚的食物、膳食纤维)来扶持宿主内源AKK菌的生长,也是一种温和而有效的方法。
关 键 挑 战
要把 AKK从研究热点真正推进到可精准应用的干预手段,还需要把菌群检测作为贯穿研发—临床—产业化全链条的基础设施,融入以下关键环节:
菌株筛选与鉴定
建立一个标准化的 AKK 菌株功能评价体系,涵盖其代谢谱、免疫调节能力、安全性等多个维度,是实现精准应用的前提。与此同时,需要配套规范化的菌群检测(如 16S/宏基因组与定量检测),用于界定不同菌株在不同微生态背景中的适配性与可重复性,避免同名不同效。
情境依赖性的机制阐明
需要更深入地研究,在不同宿主遗传背景、饮食模式和共存微生物群落的影响下,AKK 菌的功能会发生怎样的改变。这需要更复杂的动物模型(如人源化小鼠模型)和多组学技术的结合;其中,纵向菌群检测是识别“谁在场、谁在协同/拮抗、何时发生生态位迁移”的关键手段,也是将机制与真实个体差异对齐的必要条件。
安全性评估
必须对活菌制剂的抗生素抗性传播风险进行严格评估。开发不含抗性基因的工程菌株,或优先使用灭活菌及组分,是未来的方向。同时,应将菌群检测用于追踪干预后菌群结构扰动、耐药基因负荷变化以及潜在机会致病菌扩增等风险信号,实现从前期评估到使用中/使用后监测的闭环。
递送与生产工艺
作为严格厌氧菌,如何实现 AKK 菌的大规模、低成本培养,并开发出能保护其在通过胃肠道时保持活性的口服制剂,是产业化面临的技术瓶颈。
在这一过程中,菌群检测同样不可或缺:一方面用于生产端的质量控制(纯度、污染菌与批间一致性),另一方面用于应用端的效果评估与分层(基线菌群与 AKK 定植/丰度变化、关键功能菌群响应),从而把工艺参数—活性保持—体内生态学结果真正连接起来,提升可复制性与可监管性。
编辑
图源:doi.org/10.1038/s41579-024-01106-1
通过对这三篇力作的梳理,我们对Akkermansia muciniphila的认知,从一个模糊的有益菌形象,变得立体、丰满,也更加敬畏。它不再是一个简单的标签,而是一个充满动态和变数的生命体。
毒理学有一句名言:剂量决定毒性。在微生物学领域,我们或许可以引申为:情境决定属性。对于AKK菌而言,这个情境包含了我们吃下的每一口食物,我们基因中编码的每一个蛋白,以及我们肠道中与之共存的亿万菌群。
作为科研和从业工作者,我们的核心任务在于阐明这些情境依赖性效应的内在机制。未来研究需要超越简单的相关性分析,通过严谨的实验设计和先进的技术手段,明确AKK菌发挥特定生物学效应的必要条件、充分条件及其剂量-效应关系,解析宿主遗传背景、肠道微生态结构、代谢状态等因素对其功能的调控作用等。
探索之路,道阻且长,但充满希望。与各位同仁共勉。
主要参考文献
Grant ET, Monzel E, Desai MS. Navigating the duality of Akkermansia muciniphila. Nat Microbiol. 2026 Jan;11(1):20-30.
Ioannou A, Berkhout MD, Geerlings SY, Belzer C. Akkermansia muciniphila: biology, microbial ecology, host interactions and therapeutic potential. Nat Rev Microbiol. 2025 Mar;23(3):162-177.
Luo Y, Lan C, Li H, Ouyang Q, Kong F, Wu A, Ren Z, Tian G, Cai J, Yu B, He J, Wright AG. Rational consideration of Akkermansia muciniphila targeting intestinal health: advantages and challenges. NPJ Biofilms Microbiomes. 2022 Oct 17;8(1):81.
谷禾健康
Hungatella属隶属于厚壁菌门梭菌目梭菌科,是一种具运动性、严格厌氧、能形成内生孢子的杆菌,代表种包括 Hungatella hathewayi 和 Hungatella effluvii。是人类肠道中常见的细菌之一,谷禾菌群数据库显示,约90%的人群肠道中可检出该细菌。
这类细菌的不同菌株对革兰氏染色表现存在变异,例如Hungatella effluvii染色呈革兰氏阳性,而Hungatella hathewayi染色则为革兰氏阴性。提示同属细菌之间在细胞壁结构和染色特性上并不完全一致,菌株间差异较大。
它是人类肠道“重要”共生菌之一,具有较强的糖类和多糖(特别是糖胺聚糖,GAG)降解能力,能产生乙酸、丙酸等短链脂肪酸,同时在牛磺酸和氧化三甲胺(TMAO)等代谢通路中扮演重要角色。还有研究发现其与某些肿瘤化疗药物的敏感性相关,但证据仍属初步,需进一步验证因果性。
从目前的人群与动物研究来看,Hungatella在多种疾病中的丰度变化呈“双相”特点:一方面,在部分炎症性肠病(溃疡性结肠炎)和脑血管疾病(未破裂颅内动脉瘤)中表现出潜在保护作用;另一方面,在一些肿瘤、心血管及精神神经疾病中则与风险升高、预后不良或慢性炎症相关。
Hungatella是属于厚壁菌门、梭菌目、梭菌科下的一个属。Hungatella是人类肠道微生物群的成员之一,并具有重要的临床意义。
1
细胞形态
Hungatella属是一种能运动、严格厌氧、内生孢子(孢子形状为卵形至椭圆形,位于中央或近末端)的杆状细菌,长度为2-5μm,宽度为0.7-1.5μm。部分菌体还可产生粘稠胶囊。
固体琼脂培养基生长的Hungatella
编辑
doi: 10.3201/eid1011.040006.
▸ 革兰氏染色可变,取决于具体菌株
这类细菌的革兰氏染色结果因菌株而异,例如Hungatella effluvii染色呈革兰氏阳性,而Hungatella hathewayi染色则为革兰氏阴性。
一般而言,Hungatella菌落呈圆形,颜色为奶油色至灰白色,边缘稍不规则,凸起。通常,它们看起来有光泽且不透明。
注:37∘C 培养3天后,H.hathewayi 在哥伦比亚羊血琼脂培养基上形成灰白色、不透明、凸圆形、有光泽、边缘稍不规则的菌落。然而,在RCA培养基上,30°C 孵育24小时后,H.effluvii 形成奶油色、圆形、凸面和光滑的菌落,且边缘完整。
H.hathewayi经48小时培养后的菌落形态
编辑
doi: 10.3201/eid1011.040006.
2
营养和生长条件
▸ 生长环境与人体环境相近
菌株生长温度范围为10–45∘C(最佳温度为30–37∘C)、pH为5.0至8.5(最佳pH7.0)。生长可以发生在0–2.0%(w/v)NaCl 条件下,最佳条件是 0.5% NaCl,但在 3% (w/v) NaCl 条件下没有观察到生长。上述条件与人体内环境相近,说明 Hungatella可能适宜在人体肠道内定植与生长。厌氧培养的菌株通常需要24-72小时才能在RCA上形成菌落。
3
生态分布
Hungatella属分布广泛,构成正常人类微生物群的一部分,视为常见肠道成员之一。H.hathewayi(Clostridium hathewayi)最初从无胃肠道疾病的健康人粪便中分离并报道,随后也在术后血流感染患者中被检出。
编辑
另一种H.effluvii 菌株 DSM 24995T 是从用于处理益生菌制造业废发酵培养基的厌氧消化器获得的废水样品中分离出来的。
4
主要代谢与功能
▸ 糖类和多糖代谢
Hungatella属成员为糖分解菌,可发酵多种碳水化合物,包括苦杏仁苷、阿拉伯糖、纤维二糖、半乳糖、葡萄糖、乳糖、甘露糖、松三糖、蜜二糖、棉子糖、鼠李糖、核糖、水杨苷、山梨糖醇、淀粉、蔗糖和海藻糖,但不能将硝酸盐还原为亚硝酸盐。
葡萄糖发酵的主要终产物为乙酸盐和丙酸盐,并少量产生CO₂和H₂。
注:最新的功能组学研究表明,H.hathewayi在糖胺聚糖(GAG)代谢上具有非常突出的能力。
此外,Hungatella的过氧化氢酶、氧化酶和吲哚试验均为阴性,甲基红试验为阳性。其对七叶苷、淀粉和尿素的利用具有物种特异性,而所有已研究菌株均不水解明胶和酪蛋白。
▸ 与牛磺酸和氧化三甲胺含量相关
宏基因组关联研究及多个中国队列和相关动物模型的证据显示,Hungatella hathewayi 与血液中牛磺酸水平相关。但该关联尚不足以证明因果性,其在牛磺酸调控中的作用机制仍需进一步纵向研究和干预性试验加以证实。
而在另一项研究中,H.Hatheway产生氧化三甲胺(TMAO)的能力,与包括抑郁症在内的某些神经系统疾病有关。
▸ 可能会代谢5‑氟尿嘧啶
在结直肠癌队列中,H.hathewayi的肿瘤组织丰度与5-FU疗效下降及总体生存率下降呈相关关系。机制方面,研究提示可能通过CDX2/B-catenin通路的调控影响药物敏感性,但尚需因果性证据与纵向研究来证实。
5
耐药性
Hungatella属对青霉素、克林达霉素和莫西沙星的耐药性存在差异,但所有分离株均对美洛培南、β‑内酰胺/β‑内酰胺酶抑制剂及甲硝唑敏感。还发现利福昔明可抑制Hungatella hathewayi的生长。
▸ 抗生素体外敏感和实际临床药敏
•甲硝唑:迄今唯一在所有报道病例中均表现为敏感的药物。MIC 通常极低(如 0.032 μg/mL),无耐药报道,是治疗经验性厌氧覆盖首选。
•克林霉素:7/9例报道中Hungatella对其敏感。
•碳青霉烯类:约6例中表现为敏感,临床多作为重症败血症经验用药。
•β‑内酰胺/β‑内酰胺酶抑制剂:阿莫西林/克拉维酸、哌拉西林‑他唑巴坦在部分病例中敏感(4–5 例)。
•青霉素:多数早期H.hathewayi 菌株对其敏感,但不作为唯一经验药物。
•万古霉素:个别株报道为敏感,但Hungatella通常为厌氧 G+ 芽孢杆菌谱系,用万古霉素根除有限。
▸ 用药建议(基于有限病例):
•经验治疗严重感染:甲硝唑±第三/四代头孢或哌拉西林‑他唑巴坦或碳青霉烯。
•一旦确定为Hungatella:保持/调整为含甲硝唑方案通常可靠,再根据E‑test/MIC可逐步去除不必要的广谱药。
6
Hungatella属的重要物种
▸ Hungatella effluvii
细胞具运动性,杆状,长约2.0–4.0μm、宽1.0–1.5μm,革兰氏阳性。在pH6.0–8.0范围内可生长,最适pH为 7.0;适生温度为10–45℃,以30℃ 为最适。
菌株 UB-B.2ᵀ 的过氧化氢酶、氧化酶和吲哚试验为阴性,甲基红和脲酶试验为阳性;七叶苷、酪蛋白、淀粉和明胶水解阴性,不能还原硝酸盐,柠檬酸盐利用及 H₂S 产生均为阴性。
其对 L-吡咯烷酰芳香酰胺酶、脲酶和 β-吡喃半乳糖苷酶呈阳性,而对亮氨酸芳香酰胺酶、苯丙氨酸芳香酰胺酶、L-脯氨酸芳香酰胺酶、酪氨酸芳香酰胺酶、α-阿拉伯糖苷酶、β-甘露糖苷酶、七叶苷水解、β-D-岩藻糖苷酶、α-L-岩藻糖苷酶和碱性磷酸酶均为阴性。可利用半乳糖、蔗糖、葡萄糖、麦芽糖和甘露糖产酸。葡萄糖发酵的主要终产物为乙酸和丙酸。
DNA G+C 含量为 51.4 mol%。
遗传分析显示,Hungatella effluvii 与 Clostridium hathewayi(DSM 13479ᵀ)在 16S rRNA 基因水平的相似度为 97.84%,但全基因组亲缘性仅为 38.4%。在表型上,二者在七叶苷、淀粉和尿素水解等多项特征上存在显著差异。
▸ Hungatella hathewayi
细胞为严格厌氧、革兰氏阴性、具运动性的杆菌,宽0.7–1.0μm、长2.0–5.0μm,常成链排列,最长可达30个细胞。
生长pH范围为5.0–8.5,最适约7.0;温度范围15–45℃,最适约 37 ℃。菌株在含0–2.5%NaCl的培养基中生长最佳。
过氧化氢酶、氧化酶、吲哚、硝酸盐还原及 VP 试验均为阴性。葡萄糖发酵的主要终产物为乙酸、乙醇、H₂和CO₂。
能利用苦杏仁苷、阿拉伯糖、纤维二糖、果糖、半乳糖、葡萄糖、乳糖、麦芽糖、甘露糖、蜜二糖、松三糖、棉子糖、鼠李糖、核糖、水杨苷、山梨糖醇、淀粉、蔗糖、海藻糖和木糖产酸;不能利用麦芽三糖、糖原、肌醇、菊粉或甘露醇产酸。七叶苷和淀粉水解阳性,而酪蛋白、尿素、纤维素、明胶不被水解。
DNA G+C 含量为 50.7–50.9 mol%。
7
与人体及其他菌群的互作
▸ 与宿主的互作与潜在致病机制
(1)中性或有益方面
•作为肠道共生菌,可能通过产SCFA,维持结肠上皮能量与黏液层完整性;
•外泌GAG裂解酶,利用黏液层及 ECM 中的 GAG,为自身及共生菌提供碳源;在一定范围内维持黏膜更新与免疫稳态。
•牛磺酸产生:可能通过影响胆盐/抗氧化/血管平衡,对远端器官(如脑血管)产生系统性效应。
(2)有害或潜在有害方面
•当肠道屏障受损(阑尾炎、缺血、肿瘤、术后创面等),Hungatella 可由肠腔进入血流/腹腔,引起:菌血症、脓毒症、肝脓肿、胆囊炎、坏死性筋膜炎等;
•在合并其他菌(如 E. coli、Prevotella、Streptococcus constellatus)的多菌种感染中,则常与严重组织坏死、长期住院相关。
▸ 与其他肠道菌群的互作
•与Bacteroides的互补/竞争:
H.hathewayi拥有与B.thetaiotaomicron接近甚至更强的GAG降解能力,但使用的是完全不同的输运与调控网络(PTS/ABC vs SusC/D)。意味着在利用宿主黏液多糖这一生态位上,Hungatella可能与 Bacteroides形成功能冗余或在特定饮食/炎症条件下出现竞争/互补替代。
•向其他菌群“输出”寡糖与代谢中间体:
将大分子GAG裂解为寡糖/二糖,这些小分子自身能摄取利用,也可被缺乏初级外泌酶的共生菌进一步利用——这在微生态学上类似“初级降解者+次级利用者”的食物网结构。
1
影响炎症性肠病的风险
编辑
研究发现,Hungatella、Acidaminococcaceae及另外15个菌群是多种克罗恩病(CD)和溃疡性结肠炎(UC)亚型的保护因素,而Terrisporobacter、Anaerostipes及其他23个菌群与不同CD和UC亚型的风险增加相关。
▸ Hungatella有助于降低溃疡性结肠炎风险
分析显示,有9个肠道菌群对溃疡性结肠炎各亚型及肠外表现具有保护作用。其中,Lachnospiraceae 科 ND3007 组可降低溃疡性直肠乙状结肠炎风险;Ruminococcaceae UCG011和Hungatella属可降低溃疡性全结肠炎风险;Lachnospiraceae 科 ND3007 组和 Hungatella 属共同降低左侧溃疡性结肠炎风险。Prevotellaceae 科、Eubacterium fissicatena 属和 Ruminococcus gnavus 组则降低合并原发性硬化性胆管炎(PSC)的溃疡性结肠炎患者的风险。
▸ 与其他益生菌组合使用作为结肠炎的保护因子
Hungatella属和Lactobacillus属还可降低克罗恩病(CD)相关关节病变风险,而Barnesiella属则增加其发生风险。
在对比溃疡性直肠乙状结肠炎与左侧结肠炎时,Lachnospiraceae 科 ND3007 组在两者中均为保护因子,而 Hungatella 仅与左侧结肠炎相关,据此推测 Hungatella 可能参与降结肠炎症的起始。实验研究显示,克罗恩病术后吻合口处 Hungatella 丰度升高,提示其可能参与常见术后并发症的发生。
若肠道菌群在术后感染的发病机制中发挥促成作用,则针对肠道微生物群的靶向治疗或预防干预有望降低克罗恩病患者术后并发症风险。
2
丰度过高可能增加结直肠癌风险
编辑
近几十年来,早发结直肠癌(yCRC)发病率不断升高,但其肠道微生物特征仍知之甚少。既往研究多聚焦于老年发病型CRC(oCRC),尚不清楚老年患者的微生物特征能否外推至年轻患者。为此,有研究整合了迄今规模最大的 yCRC 肠道宏基因组数据,涵盖两个独立队列,发现CRC相关微生物组。
▸ H.hathewayi在结直肠癌患者中富集
分析显示,如Clostridium symbiosum、Peptostreptococcus stomatis、Parvimonas micra和Hungatella hathewayi在老年和年轻患者中均显著富集。并且在老年发病结直肠癌(oCRC)和早发结直肠癌(yCRC)中,观察到具核梭杆菌(Fusobacterium nucleatum)、脆弱拟杆菌(Bacteroides fragilis)和大肠杆菌(Escherichia coli)的菌株水平模式相似。
几乎所有与oCRC相关的宏基因组通路在年轻患者中均有方向一致的变化。值得注意的是,与对照组相比,oCRC和yCRC的相关毒力因子(fadA、bft)均为富集。此外,基于微生物组的分类模型在老年和年轻发病患者中对结肠癌状态的预测准确性相似,强调了不同年龄组微生物特征的一致性。
▸ H.hathewayi影响5‑氟尿嘧啶代谢进而干扰治疗效果
此外,刚才前文中也有讲到过的一项研究发现H.hathewayi在结直肠癌组织中的丰度显著增加,其高水平与整体生存率降低相关。
并且H.hathewayi处理明显减弱了5-氟尿嘧啶(5-FU)对结直肠癌(CRC)细胞系HCT116和HT29的增殖抑制和凋亡诱导作用,并增强了耐药细胞系 HCT116/5‑FU 和 HT29/5‑FU 对 5‑FU 的耐药性。机制上,H.hathewayi下调CDX2表达并促进 β‑catenin 在细胞核内的积累。CDX2 过表达可逆转H.hathewayi介导的细胞生长增强和对 HCT116/5‑FU、HT29/5‑FU 细胞凋亡的抑制,同时抑制 β‑catenin 的表达及其核内富集。
综上所述,结直肠癌组织中H.hathewayi高丰度不仅与较差生存预后相关,还可通过调控 CDX2/β‑catenin 信号通路影响结直肠癌细胞对5‑FU的耐药性。
3
未破裂动脉瘤患者中丰度较低
未破裂颅内动脉瘤(UIA)是一种危及生命的脑血管疾病。在两组中国UIA患者及接受人类供体粪便移植的对照个体和小鼠中进行了病例对照宏基因组全域关联研究。
注:非破裂颅内动脉瘤的全球患病率为3.2%,中国的社区调查表明中国成年人非破裂颅内动脉瘤患病率高达7%。颅内动脉瘤是一种极为凶险的脑血管疾病,一旦破裂其早期致死率为40-50%。
▸ 颅内动脉瘤患者中H.hathewayi减少,且与牛磺酸正相关
研究团队发现一群Clostridium属的物种包括C.bartlettii, C.nexile和C.bolteae在颅内动脉瘤组显著富集。然而,C.hathewayi(在2014年被更名为Hungatella hathewayi)却是唯一一个在颅内动脉瘤肠道中丰度降低的Clostridium属成员。
UIA患者的肠道微生物改变
编辑
doi: 10.1038/s41467-020-16990-3.
具体而言,在未破裂颅内动脉瘤(UIA)患者中Hungatella hathewayi 的丰度显著下降,并与人及小鼠体内的牛磺酸水平呈正相关。灌胃补充H.hathewayi可使小鼠血清牛磺酸水平恢复正常,并保护其免于颅内动脉瘤的形成与破裂;补充牛磺酸同样可逆转颅内动脉瘤的进展。上述结果提示,H.hathewayi相关的牛磺酸耗竭可能是UIAs发病机制中的关键因素。
此外,与单纯移植动脉瘤患者粪便的小鼠相比,补充H.hathewayi可显著降低颅内动脉瘤的总体发生率。Kaplan–Meier分析进一步证实,H.hathewayi 补充降低了动脉瘤破裂风险。同时,H.hathewayi 还能抑制脑血管炎症反应、细胞外基质重塑、MMP‑9 活性以及脑血管平滑肌细胞凋亡。
综上,H.hathewayi参与牛磺酸合成并提高血浆牛磺酸水平,从而降低颅内动脉瘤的发生率和破裂率。
4
在一些精神疾病中丰度较高
▸ 慢性精神分裂症患者中Hungatella丰度增加
编辑
研究共纳入115例处于阳性和紊乱症状缓解期的精神分裂症患者及119名对照者,评估了32项外周血标志物。
研究发现:在慢性精神分裂症患者中,Hungatella 的丰度显著增加,相比健康对照组其相对丰度更高。患者的肠道菌群多样性整体下降,而 Hungatella 等部分优势菌属呈富集状态。
差异丰度属与免疫炎症标志物、肠毒素、脂质谱成分和胰岛素抵抗水平的变化相关。此外,还观察到差异丰度属与认知障碍、阴性症状严重程度增加以及社交功能恶化存在若干相关性。甲烷短杆菌属(Methanobrevibacter)丰度与阴性症状、认知和社会功能水平的相关性似乎受白细胞介素-6和RANTES水平介导。
而Hungatella与注意力表现的相关性则受肠毒素水平介导。研究结果表明,精神分裂症患者肠道微生物群组成的改变与临床表现、肠道通透性、亚临床炎症、脂质谱改变以及葡萄糖稳态受损相关。亚临床炎症和肠道通透性受损可能介导了肠道微生物群改变与精神病理症状和认知障碍之间的关联。
▸ 产后抑郁的女性中H.hathewayi的含量较高
编辑
在另一项研究中,发现产后抑郁症状女性中 Hungatella hathewayi含量较高。在不同组学和横断面分析中观察到多种菌种丰度差异:中度且加重抑郁症状组中 Hungatella hathewayi丰度较高(FDR<0.25);高度且减轻抑郁症状组中 Bacteroides clarus 丰度较高(FDR<0.25)。
在妊娠晚期,与“低且稳定”组相比,“爱丁堡产后抑郁量表(EPDS)高且下降”组中 Bacteroides clarus、B. faecis 和 B. xylanisolvens 的相对丰度较高;而“EPDS 中且上升”组中 Hungatella hathewayi、Lachnospira SGB5076 和 Streptococcus thermophilus 的相对丰度较高。
既往研究也发现,H.hathewayi在非妊娠重度抑郁障碍人群中水平升高。其可能机制与产生三甲胺-N-氧化物(TMAO)的能力有关;TMAO 与包括抑郁在内的多种神经系统疾病有关,循环 TMAO 可激活促炎信号通路,从而诱发神经炎症。
5
冠状动脉病变中丰度升高
▸ 川崎病急性期合并冠状动脉病变儿童中富集
编辑
在川崎病急性期合并冠状动脉病变(AKDCAL)的儿童中,LEfSe 分析显示Hungatella显著富集,而在无冠状动脉病变的 AKDNCAL 组未见此现象。
注:川崎病(KD)是一种急性全身性免疫血管炎,主要影响中小动脉,常见于儿科患者。
虽然总体α多样性在川崎病急性期合并冠状动脉病变与无冠状动脉病变川崎病间无显著差异,但Hungatella、Burkholderia-Caballeronia-Paraburkholderia和Clostridium_innocuum富集,被鉴定为与冠状动脉病变(CALs)相关的特征菌属。
6
其他疾病中的丰度变化
⑴丰度增加可能与婴儿湿疹风险升高相关
在一项香港队列研究中,肠道Hungatella hathewayi丰度随时间增加与1岁前发生湿疹的风险升高显著相关。
⑵有时可能引起菌血症或严重感染
Hungatella 在多数情况下为肠道共栖菌,但在特定易感人群或肠道屏障受损的情况下,也可能导致菌血症或其他严重感染。
H.effluvii和H.hathewayi偶见引起败血症、坏死性筋膜炎或阑尾炎相关菌血症等重症感染。并且比较基因组学提示Hungatella属内部遗传差异大,不同“种/亚种”在毒力和耐药基因上差异明显。
⑶伴便秘的帕金森病患者中增多
与健康对照相比,伴有便秘的帕金森病(PD)患者粪便中Hungatella显著增多,同时伴随Collinsella升高、Lachnospira和Fusicatenibacter降低,提示整体向“促炎型菌群”转变。
反映便秘严重程度的 Wexner 评分与 Hungatella 丰度呈正相关,即Hungatella越高,便秘越重。
在机制层面,目前证据支持其作为参与PD相关肠道菌群失调和便秘,可能通过影响 SCFA 平衡、肠道炎症与肠‑脑轴功能发挥作用,是帕金森病合并便秘中值得关注的潜在干预靶点。
⑷乳腺癌患者中Hungatella丰度增加
乳腺癌(BCa)患者微生物多样性减少,但从BCa患者采集的粪便样本中,三种特定微生物属——Acidaminococus、Tyzzerella和Hungatella的丰度得到了富集。
⑸新冠后遗症/康复患者中增加
在新冠后遗症或康复期人群中,有研究发现,其肠道中Hungatella的总体丰度持续处于升高状态。进一步的分析显示,这种Hungatella丰度的增加与多种炎症因子水平呈显著正相关关系,尤其是与 IL‑8、IL‑1β 等经典促炎细胞因子相关联,因此提示 Hungatella 的升高可能反映或参与了机体长期存在的慢性炎症过程,与新冠感染后的持续性炎症状态具有一定联系。
综合目前多学科证据,Hungatella 在疾病中的作用大致通过以下几点体现:
1.黏膜与多糖代谢:强糖胺聚糖降解能力一方面为自身及其他菌群提供碳源,支持黏膜更新和生态食物网;另一方面,若失衡或过度活跃,可能削弱肠黏液屏障,增加内毒素和炎症因子易位,驱动全身慢性炎症。
2.小分子代谢(SCFA、牛磺酸、TMAO 等):
-通过产乙酸/丙酸等 SCFA 参与能量代谢和局部抗炎;
-通过促进牛磺酸合成或维持其水平,对脑血管等远端器官产生保护作用(如预防颅内动脉瘤);
-通过产生氧化三甲胺(TMAO),参与动脉粥样硬化和神经精神疾病相关的促炎与代谢异常。
3.免疫与炎症信号:与多种炎性细胞因子(IL‑6、IL‑8、IL‑1β 等)水平相关,可能通过诱导肠炎和系统性低度炎症,连接肠道微生态与代谢综合征、心血管疾病及精神神经障碍。
4.表观遗传与肿瘤信号通路:在结直肠癌中,通过调控 DNA 甲基转移酶活性,影响抑癌基因甲基化,以及通过 CDX2/β‑catenin 通路干预细胞增殖和 5‑FU 化疗敏感性,表现出典型的“促肿瘤菌”特征。
5.肠‑脑/肠‑血管轴:在帕金森病、抑郁、精神分裂症、川崎病和颅内动脉瘤等疾病中的一系列关联,提示 Hungatella不仅是肠腔共栖者,更可能通过代谢物与炎症通路参与中枢神经和血管系统的病理过程。
总体而言,Hungatella 是一个功能高度复杂且菌株异质性很大的肠道菌属:
•在某些场景中(如未破裂颅内动脉瘤、部分溃疡性结肠炎亚型),它可能具有保护性作用;
•在另一些场景中(如结直肠癌、乳腺癌、部分心血管和精神神经疾病、婴儿湿疹、新冠后遗症),其丰度升高则更多与风险增加、预后不良或慢性炎症相关;
•而在免疫低下或屏障破坏条件下,还可作为机会致病菌引发严重感染。
现有研究多为关联性和部分机制探索,因物种鉴定精度、株间差异及干预研究仍有限,Hungatella 在各类疾病中的真正“因果角色”和可干预潜力仍需更多纵向、功能和临床试验加以澄清。
注:本账号内容仅供学习和交流,不构成任何形式的医疗建议。
主要参考文献
Rawat PS, Li Y, Zhang W, Meng X, Liu W. Hungatella hathewayi, an Efficient Glycosaminoglycan-Degrading Firmicutes from Human Gut and Its Chondroitin ABC Exolyase with High Activity and Broad Substrate Specificity. Appl Environ Microbiol. 2022 Nov 22;88(22):e0154622.
Yang Q, Tang W, Ren J, Li M, Zhao C. Unveiling the gut-heart potential connection: microbiota’s role in kawasaki disease and coronary artery lesions. Front Cell Infect Microbiol. 2025 May 29;15:1560083.
https://doi.org/10.1002/9781118960608.gbm01640Digital Object Identifier (DOI)
Li F, Yu C, Zhao Q, Wang Z, Wang Z, Chang Y, Xu Z, Han X, Li H, Liu Y, Hu S, Chang S, Tang T, Li Y. Exploring the intestinal ecosystem: from gut microbiota to associations with subtypes of inflammatory bowel disease. Front Cell Infect Microbiol. 2024 Jan 4;13:1304858.
Qin Y, Tong X, Mei WJ, Cheng Y, Zou Y, Han K, Yu J, Jie Z, Zhang T, Zhu S, Jin X, Wang J, Yang H, Xu X, Zhong H, Xiao L, Ding PR. Consistent signatures in the human gut microbiome of old- and young-onset colorectal cancer. Nat Commun. 2024 Apr 22;15(1):3396.
Huang Z, Wang C, Huang Q, Yan Z, Yin Z. Hungatella hathewayi impairs the sensitivity of colorectal cancer cells to 5-FU through decreasing CDX2 expression. Hum Cell. 2023 Nov;36(6):2055-2065.
Li H, Xu H, Li Y, Jiang Y, Hu Y, Liu T, Tian X, Zhao X, Zhu Y, Wang S, Zhang C, Ge J, Wang X, Wen H, Bai C, Sun Y, Song L, Zhang Y, Hui R, Cai J, Chen J. Alterations of gut microbiota contribute to the progression of unruptured intracranial aneurysms. Nat Commun. 2020 Jun 25;11(1):3218.
Hieta J, Benchraka C, Pärnänen K, Houttu N, Mokkala K, Lotankar M, Kataja EL, Lahti L, Laitinen K. Perinatal depressive and anxiety symptoms are associated with gut microbiota in pregnant women with overweight and obesity. Brain Behav Immun Health. 2025 Jun 19;47:101042.
Misiak B, Pawlak E, Rembacz K, Kotas M, Żebrowska-Różańska P, Kujawa D, Łaczmański Ł, Piotrowski P, Bielawski T, Samochowiec J, Samochowiec A, Karpiński P. Associations of gut microbiota alterations with clinical, metabolic, and immune-inflammatory characteristics of chronic schizophrenia. J Psychiatr Res. 2024 Mar;171:152-160.
Elsayed S, Zhang K. Human infection caused by Clostridium hathewayi. Emerg Infect Dis. 2004 Nov;10(11):1950-2.
谷禾健康
肠道微生物组作为人体“第二基因组”,在癌症的发生、发展及治疗反应中扮演着日益重要的角色。这些微生物不仅在局部和全身层面调节免疫应答,还深刻影响着炎症、免疫监视和耐受性。
平衡良好的微生物群是有效免疫功能的基石,但由饮食,环境因素,生活方式,感染或药物驱动的干扰可能会促进慢性炎症并损害抗肿瘤免疫力。越来越多的证据表明,这些干扰不仅有助于肿瘤的发生和发展,而且还对治疗产生耐药性。
微生物及其代谢产物能够通过局部及全身性机制,直接影响药物代谢、塑造免疫微环境,并改变肠道、全身乃至肿瘤微环境(TME)中的转录和表观遗传程序。
大量新兴数据支持,微生物组作为预后和预测性生物标志物、癌症防治中可调控因素以及治疗靶点的巨大潜力。以微生物组为靶点的干预措施(如粪菌移植、饮食调整、益生元和益生菌)可作为传统癌症疗法的辅助手段,增强疗效并降低毒性。
本文探讨了微生物组驱动癌症发生发展的多维机制,包括基因毒性、慢性炎症和表观遗传调控。详细分析微生物组如何调节化疗、放疗及免疫治疗的疗效与不良反应,还系统梳理了当前基于微生物组的癌症治疗策略,如粪菌移植、饮食干预、益生菌和后生元等,并总结了相关临床试验的进展。
历史上,微生物与癌症的关联常采用科赫假设(Koch’s postulates)等框架,将疾病归因于具有确定潜伏期的单一病原体。然而,现在人们认识到,微生物作为复杂的群落发挥作用,对宿主产生集体效应,而不仅仅是孤立的病原体。这些效应包括调节炎症、免疫、代谢和表观遗传,从而深刻影响致癌过程。
微生物驱动的致癌机制
微生物群通过多种复杂的机制促进肿瘤的启动和进展,主要包括直接的基因毒性效应、诱导慢性炎症和肠道屏障功能障碍,以及对宿主细胞的表观遗传调控。这些机制相互交织,共同构成了一个促癌的微环境。
肠道微生物组可通过诱导基因组损伤、表观遗传调控或破坏肠道屏障功能参与癌症的启动和/或进展。这些机制共同作用,促进了从正常上皮到恶性肿瘤的转化。
病原菌的致癌策略:信号传导与毒素的双重威胁
定植于上皮表面的病原菌或机会性细菌可通过直接的宿主细胞信号传导、微生物间通讯或分泌毒素发挥致癌作用。其中,细菌基因毒素是研究最为深入的直接致癌因素之一。这些毒素包括tilimycin、indolimines、tilivalline、colibactin和脆弱拟杆菌毒素(Bacteroides fragilis toxin, BFT)等。
编辑
-Colibactin
在这些毒素中,Colibactin的机制研究最为透彻。Colibactin由存在于变形菌门(如特定大肠杆菌菌株)中的聚酮合酶(pks)基因岛编码。这种基因毒素能够诱导DNA双链断裂和烷基化,从而引发基因组不稳定性,促进癌症的发生。
编辑
图源:doi.org/10.1038/s41579-025-01268-6
例如,一项研究发现,在结直肠癌中,由产colibactin的大肠杆菌(pks+ E. coli)引起的突变特征与早发性CRC相关。此外,产BFT的脆弱拟杆菌(ETBF)能够协同pks+ E. coli,其产生的BFT和黏液溶解活性使得colibactin能够穿透黏液层,到达上皮细胞,从而加剧DNA损伤。
微生物代谢物:潜在的DNA损伤元凶?
除了直接的基因毒素,微生物代谢物也可能具有DNA损伤效应。例如,有研究指出微生物将初级胆汁酸转化为次级胆汁酸,后者过量可通过诱导DNA损伤、调节炎症通路和抑制抗肿瘤免疫反应(包括抑制在癌症免疫监视中至关重要的细胞毒性CD8+ T细胞)来促进结直肠肿瘤的发生。
同样,肠道微生物将管腔中的乙醇转化为乙醛,或将某些亚硝胺类致癌物(如N-丁基-N-(4-羟丁基)亚硝胺)转化为更具活性的形式,这些过程均可导致基因组损伤。
雌激素与微生物:远程影响
此外,微生物的β-葡萄糖醛酸酶(GUS)能够重新激活在肝脏中被结合失活的雌激素,提高循环雌激素水平,这可能对乳腺癌等雌激素敏感性肿瘤的发生构成远距离影响。
慢性炎症与肠道屏障功能障碍
慢性微生物感染是癌症发生的重要驱动力
例如,幽门螺杆菌(Helicobacter pylori)感染通过诱导持续性胃部炎症,促进胃癌的发生,而根除幽门螺杆菌已被证实可以预防胃癌。同样,伤寒沙门氏菌(Salmonella typhi)与胆囊癌的发生相关。
编辑
图源:doi.org/10.1038/s41579-025-01268-6
微生物组失衡促癌变
即使在没有明显感染的情况下,微生物组失衡也能通过调节局部炎症反应促进癌变。
例如,通常存在于口腔的具核梭杆菌(Fusobacterium nucleatum)在特定条件下可定植于肠道,并在CRC肿瘤中诱导肿瘤微环境的炎症。
炎症与癌症之间的复杂信号网络
微生物诱导的炎症部分通过模式识别受体(如Toll样受体,TLRs)信号和炎症小体激活介导,导致细胞因子和活性氧(ROS)的产生,这些分子刺激上皮增殖、免疫逃逸以及癌细胞在远端部位的播散和植入。
肠道炎症与肠道屏障功能障碍密切相关
屏障功能受损,即黏液层和上皮细胞无法在管腔微生物群落与宿主上皮之间提供足够的空间,增加了促癌微生物和分泌性基因毒素造成损害的易感性。受损的屏障完整性促进了微生物产物(如脂多糖,LPS)和代谢物(如胆汁酸)的易位,进入体循环。这些因素可驱动远端组织的全身性炎症或免疫抑制,可能导致远端上皮和非上皮癌症及转移的发生。
编辑
保护性微生物代谢物的作用
一些微生物代谢物,如膳食纤维在结肠中发酵产生的短链脂肪酸(SCFAs),通过促进紧密连接形成和刺激黏液产生,对肠道屏障具有保护作用。
表观遗传调控
肠道微生物组是一个强大的表观遗传调节器,能够通过代谢、炎症和饮食-微生物相互作用的组合来塑造基因表达和肿瘤行为。
图源:doi.org/10.1038/s41579-025-01268-6
丁酸盐的双重角色:从能量源到表观遗传调控
最典型的微生物驱动的表观遗传调控机制是SCFA中的丁酸盐对组蛋白去乙酰化酶(HDAC)的抑制。在健康的上皮细胞中,丁酸盐是支持增殖和分化的能量来源。然而,随着肿瘤细胞代谢转向有氧糖酵解(瓦博格效应),丁酸盐不再被有效消耗。其积累导致过度HDAC抑制,改变染色质构象,促进抑癌基因的转录激活,从而抑制癌变。
膳食成分与微生物的影响
其他微生物或膳食成分也能影响表观基因组。例如,叶酸、亚硝胺和致肥胖饮食在小鼠模型中影响DNA甲基化和组蛋白乙酰化,促进肠道肿瘤发生。
产毒脆弱拟杆菌的致癌机制
有趣的是,在小鼠模型中,产肠毒素的脆弱拟杆菌(ETBF)定植通过破坏DNA修复机制促进结直肠肿瘤发生,并诱导广泛的CpG岛高甲基化,这是肿瘤组织中表观遗传变化的标志。
不同类型的癌症与特定的微生物组特征相关联,这些特征不仅存在于胃肠道肿瘤中,也见于乳腺癌、前列腺癌等远端肿瘤,揭示了微生物组在肿瘤生态系统中的广泛影响。
结 直 肠 癌
结直肠癌(CRC)的发展与多种微生物、其代谢物和结肠上皮之间的相互作用密切相关。在动物模型和细胞培养研究中,特定微生物如ETBF(产肠毒素的脆弱拟杆菌)、具核梭杆菌(F. nucleatum)和携带pks基因岛的大肠杆菌(pks+ E. coli)被证明有助于肿瘤发生。
将结直肠癌患者的粪便微生物群移植到无菌小鼠体内,会导致细胞增殖增加、息肉数量增多、异型增生更严重以及炎症标志物水平升高,这强烈表明肠道微生物组失衡可能是结直肠癌发展的致病因素。
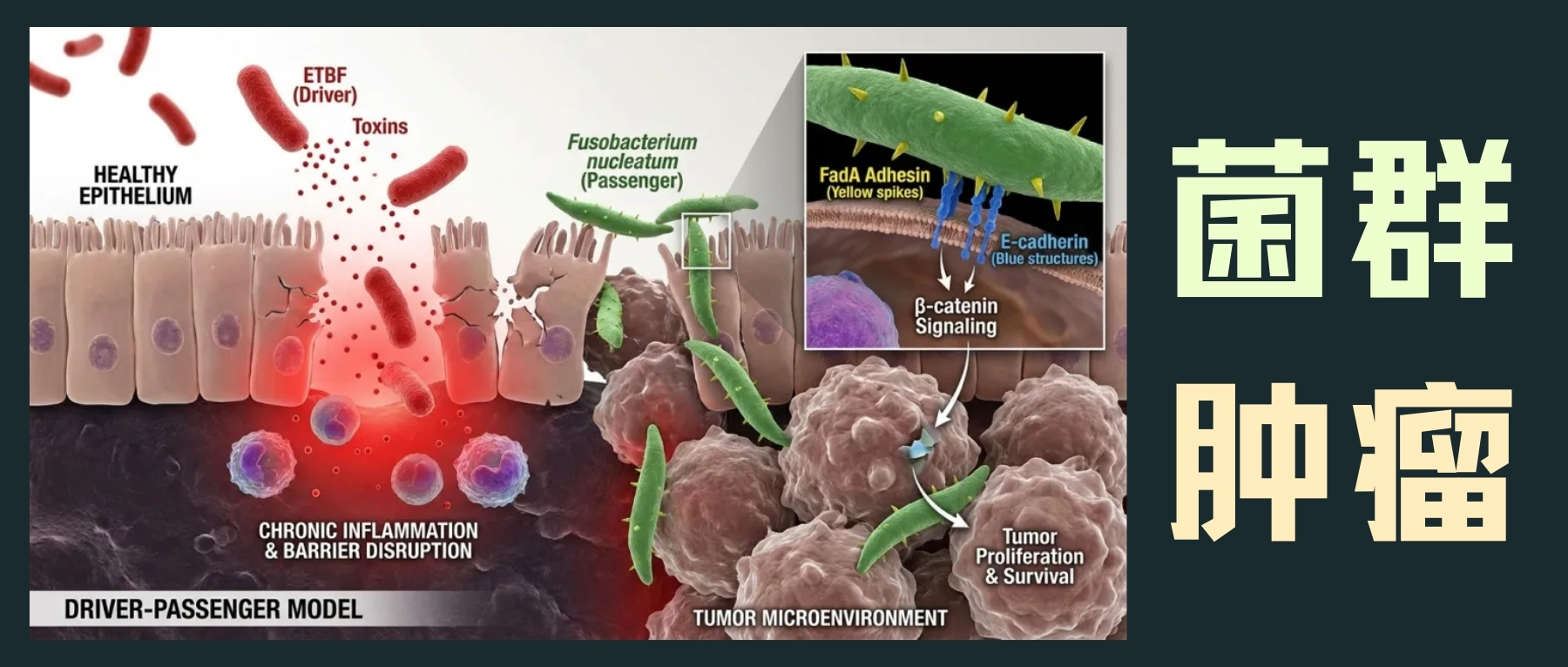
驱动-乘客模型:微生物的协同作用
“驱动-乘客”模型提出,像ETBF这样的驱动菌通过引发慢性炎症和破坏上皮屏障,创造一个允许性环境,让像具核梭杆菌这样的共生乘客菌得以定植并加剧肿瘤进展。具核梭杆菌通过其细胞表面的FadA黏附素与上皮E-钙黏蛋白直接相互作用,激活β-连环蛋白信号通路,促进肿瘤细胞增殖和存活。
编辑
具核梭杆菌与化疗耐药性
此外,具核梭杆菌还通过刺激肿瘤细胞中的自噬相关蛋白,与化疗药物的细胞毒性作用相抗衡,从而与化疗耐药性相关。临床上,结直肠癌肿瘤组织中的具核梭杆菌DNA水平较高与结直肠癌特异性死亡率增加相关,表明其可作为潜在的预后生物标志物。
胃 癌
在胃癌中,幽门螺杆菌(H. pylori)感染是公认的关键致病因素。它通过其毒力因子(如CagA和VacA,在谷禾16S Plus检测实践中,也会检出个别受检者携带 CagA和VacA毒力基因)引发炎症和免疫反应,进而可能有诱发癌症风险。然而,随着胃癌进展,胃内环境发生萎缩,幽门螺杆菌数量反而减少,而其他非幽门螺杆菌物种(如乳酸杆菌属、链球菌属)开始增多。
肝 细 胞 癌
在肝细胞癌(HCC)中,肠-肝轴起着关键作用。肠道屏障功能受损导致细菌及其产物(如LPS)易位至肝脏,诱发炎症、纤维化和肝细胞损伤,最终促进癌变。
肝细胞癌患者的肠道微生物组通常表现为产SCFA的细菌多形拟杆菌(Bacteroides thetaiotaomicron)减少,而促炎细菌(如大肠杆菌)增多。
胰 腺 导 管 腺 癌
胰腺导管腺癌(PDAC)同样与肠道微生物组失调相关。肠道屏障受损允许某些肠道微生物(如变形菌门)易位至胰腺,在局部创造一个免疫抑制的微环境,促进肿瘤进展。此外,微生物对胆汁酸的代谢异常也被认为是PDAC的风险因素之一。
激素依赖性癌症
肠道微生物组在激素代谢中发挥着重要作用,从而影响激素依赖性肿瘤的进展。所谓的雌激素菌基因组(estrobolome)是指能够代谢雌激素的细菌基因集合。肠道中的细菌β-葡萄糖醛酸酶可以解偶联在肝脏中结合的雌激素,使其重新被吸收入血,提高循环中活性雌激素的水平,从而促进乳腺癌等雌激素驱动肿瘤的生长。
在乳腺癌患者中,具有β-葡萄糖醛酸酶活性的菌种(如梭菌属)丰度增加可能与更严重的临床分期有关。同样,在前列腺癌中,肠道菌群能够从头合成雄激素或将皮质醇代谢物转化为雄激素,促进对内分泌治疗的抵抗。
癌症的多步发生模型认为,遗传和表观遗传的序贯改变是恶性转化的驱动力。新兴证据表明,微生物组的失调可能是这一模型中一个额外的、相互关联的因素。
生命早期的微生物组发育
微生物组的发育始于生命早期,受分娩方式、喂养习惯和饮食转变的影响。在这一关键窗口期的干扰可能导致微生物多样性永久性降低和对免疫与代谢稳态至关重要的关键菌种的缺失。
生活方式对微生物组的影响
在整个生命周期中,累积的损害(包括抗生素滥用、慢性压力和低纤维、超加工饮食)会进一步侵蚀微生物的多样性和功能。这种微生物恢复力的逐步丧失可能会损害上皮完整性、增加炎症、削弱代谢信号和免疫监视。
多个研究和科学家提出,微生物组的这种纵向退化可能会降低癌症启动和进展的阈值,与经典致癌过程中的逐步遗传事件相平行。
微生物组健康随生命周期的逐步退化可能促进癌症发展
编辑
图源:doi.org/10.1038/s41579-025-01268-6
图中显示了肠道微生物组健康和恢复力的两种对比轨迹
每次急性干扰后恢复不完全,导致微生物组健康和恢复力逐步退化。
持续的压力源可能将个体推向恶化轨迹,直至达到癌症发展概率更高的阈值。
癌症治疗反应的变异性受到多种宿主和肿瘤相关因素的影响,而肠道微生物组是导致这种变异性的一个重要贡献者。
微生物组主要通过两大机制调节癌症治疗结果:
一是通过微生物对治疗药物的生物转化以及药物对微生物组的反作用;
二是通过微生物介导的宿主免疫反应调节。
微生物组与化疗、放疗及靶向治疗
肠道微生物组编码着庞大的基因库,能够代谢多种肿瘤学和非肿瘤学药物,从而影响药物的生物利用度、疗效和毒性。
微生物介导的影响癌症治疗结果的机制
编辑
图源:doi.org/10.1038/s41579-025-01268-6
药物的微生物生物转化
一个典型的例子是化疗药物5-氟尿嘧啶(5-FU)及其口服前药卡培他滨。某些大肠杆菌和厌氧棒状菌属(Anaerostipes)表达细菌二氢嘧啶脱氢酶(DPD),能将5-FU转化为其非活性形式,从而可能降低其疗效。
类似地,用于治疗胰腺癌的吉西他滨可被某些肿瘤相关γ-变形菌纲细菌表达的胞苷脱氨酶(CDA)脱氨而失活,导致耐药。
微生物也能激活药物
拓扑异构酶I抑制剂伊立替康在肝脏中被代谢为活性形式SN-38,随后经葡萄糖醛酸化为无活性的SN-38G,通过胆汁排入肠道。肠道中的细菌β-葡萄糖醛酸酶(GUS)能将SN-38G重新激活为SN-38,后者会引起严重的上皮损伤、黏膜炎和腹泻等剂量限制性毒性。虽然理论上肿瘤内的再激活可能增强疗效,但肠道内的再激活主要导致毒性增加。
微生物影响疗效与毒性
除了直接代谢药物,微生物组还能通过调节免疫系统间接影响化疗效果。例如,化疗药物环磷酰胺能促进特定肠道细菌(如Enterococcus hirae)易位至次级淋巴器官,诱导产生有助于其治疗效果的TH17细胞(T辅助17细胞)反应。
同样,奥沙利铂的疗效被发现依赖于肠道微生物组,肠道菌群通过激活免疫系统来增强抗肿瘤效果。
放疗与微生物组之间的双向互动
一方面,放疗(尤其是盆腔放疗)会显著改变肠道微生物组的组成,导致多样性下降,并可能增加机会性病原菌。
另一方面,微生物组的组成可以预测患者对放疗的反应和毒性。例如,放疗前肠道菌群多样性较低与晚期放射性肠病相关。在小鼠模型中,粪菌移植(FMT)被证明可以保护机体免受辐射引起的毒性。
靶向治疗
对于靶向治疗,研究尚处早期。一项针对转移性CRC的研究发现,对抗EGFR(西妥昔单抗)和抗VEGF(贝伐珠单抗)治疗的疾病进展组患者,其肠道微生物组中具核梭杆菌的丰度显著高于部分缓解组。
在HER2阳性乳腺癌中,抗生素使用或移植来自抗生素处理供体的小鼠粪菌会削弱曲妥珠单抗的抗肿瘤效果。
微生物组与免疫治疗
免疫检查点抑制剂(ICIs)通过阻断T细胞上的抑制性信号(如PD-1、CTLA-4),重新激活细胞毒性CD8+ T细胞以攻击肿瘤,彻底改变了癌症治疗。然而,只有部分患者对ICIs有反应,而肠道微生物组是决定疗效的关键因素之一。
►▷ 微生物组如何调节免疫反应与治疗效果
多项早期研究独立证实,肠道微生物组的组成与抗PD-1/PD-L1治疗的反应相关。
尽管与疗效相关的具体菌种在不同研究中有所差异(如Akkermansia、Bifidobacterium、Ruminococcus),但其免疫调节机制常常趋同于增强CD8+ T细胞的数量或活性。
例如,在临床前模型中,口服双歧杆菌可增强树突状细胞(DC)的激活和CD8+ T细胞的启动,从而增强对PD-L1阻断的反应。同样,Akkermansia muciniphila通过促进CD8+ T细胞招募至肿瘤,来增强对PD-1抑制的反应。
这些发现表明,与ICI疗效相关的菌种可能共享一种能力,即在肠系膜淋巴结或肠黏膜中诱导依赖于DC的CD8+ T细胞启动,随后通过趋化因子介导浸润至TME。
肠道菌群的影响:U型效应
值得注意的是,微生物组对ICI疗效的影响可能具有“U型”或“倒U型”特征。
编辑
例如,一项针对晚期非小细胞肺癌(NSCLC)患者的研究发现,肠道中A. muciniphila的相对丰度处于中等水平(而非低或高水平)的患者,其临床反应最佳。这提示微生物的影响并非简单的“越多越好”,而是需要一个平衡的生态系统。
►▷ 微生物代谢物:抗肿瘤免疫的关键因素
微生物的功能冗余性可能比分类学组成更为关键,即不同的微生物群落可能产生相似的代谢或免疫输出。多种微生物代谢物被证明在调节免疫治疗反应中发挥核心作用。
肠道微生物代谢物与免疫
编辑
图源:doi.org/10.1038/s41579-025-01268-6
该图描绘了肠道微生物组与免疫系统之间的相互作用,说明了膳食成分(如纤维)如何影响SCFAs、胆汁酸和色氨酸衍生物的产生。这些代谢物调节多种免疫细胞,并通过调节炎症细胞因子和增强癌细胞凋亡来促进抗肿瘤免疫和ICI治疗效果。
编辑
图源:doi.org/10.1038/s42255-025-01287-w
代谢物的作用是情境依赖的
例如,丁酸盐在某些情况下能增强细胞毒性CD8+ T细胞功能,支持抗肿瘤免疫,但在其他条件下也可能促进免疫抑制性的Treg细胞扩增。
同样,具核梭杆菌在不同研究中显示出截然相反的作用:一项研究表明其产生的丁酸能敏化MSS型CRC对ICIs的反应,而另一项研究则发现其产生的琥珀酸会抑制免疫信号通路,导致耐药。
这凸显了菌株、肿瘤微环境和代谢产物背景的复杂性。
►▷ 肠道菌群预测免疫相关不良事件
ICIs激活的免疫系统也可能攻击正常组织,导致免疫相关不良事件(irAEs)。多项报告将基线肠道微生物组组成与irAEs的发生风险联系起来。
然而,由于患者群体异质性、队列规模小以及irAEs定义不一致等原因,目前尚未发现一致的预测性微生物标志物。一些研究提示,特定的菌群(如拟杆菌门丰度较高)可能与结肠炎风险增加有关,但仍需更多机制性研究来阐明因果关系。
鉴于微生物组与癌症疗效和毒性的密切联系,操纵微生物组成员为增强治疗效果提供了新的契机。一系列策略正在研究中,包括饮食干预、益生菌/益生元、粪菌移植(FMT)以及更前沿的工程菌和后生元等。
这些干预措施旨在通过重塑微生物组生态系统,来调节药物代谢、增强抗肿瘤免疫或减轻治疗毒性。
编辑
图源:doi.org/10.1038/s42255-025-01287-w
粪菌移植 (FMT)
粪菌移植是目前研究最深入的微生物组干预手段之一。两项开创性的I期临床试验表明,将来自对ICI有反应的黑色素瘤患者的粪便微生物群移植给耐药患者,可以使一部分患者转为应答者,并伴随着CD8+ T细胞活化增强或TME中免疫细胞浸润的有利变化。
最近,一项在晚期黑色素瘤患者中进行的 I期试验进一步证实,接受健康供体FMT后,患者对一线抗PD-1治疗的反应得到改善,其粪便样本能够在小鼠模型中恢复抗PD-1疗效。
此外,FMT也被用于治疗难治性ICI相关性结肠炎,初步研究显示其能有效缓解症状,并伴随免疫细胞谱的改变。目前,大量临床试验正在探索FMT联合ICI在不同癌种(如肾细胞癌、NSCLC)中的应用。
饮食、益生元与益生菌
饮食干预是一种极具性价比的策略
一项观察性研究发现,高纤维饮食(不使用益生菌)与改善的ICI结局相关。
BE GONE试验(NCT02843425)等临床研究正在评估高纤维食物(如豆类)对微生物组和临床结局的影响,初步结果显示其能增加有益菌(如Faecalibacterium)并改善炎症标志物。
注:zhuBE GONE试验(NCT02843425)是一项由MD安德森癌症中心开展的临床研究,旨在探索食用 canned navy beans对肠道微生物组的影响,以及这种饮食干预是否能减轻肥胖对结直肠癌风险的负面影响。
传统益生菌
益生菌的应用也备受关注。一些传统益生菌(如某些乳杆菌和双歧杆菌)被发现能通过产生特定的代谢物(如I3A、吲哚-3-羧酸)或直接作用来增强抗肿瘤免疫。
下一代益生菌 和 工程菌
例如,A. muciniphila被多项研究证实与ICI的良好反应相关,其巴氏杀菌形式(一种后生元)或其膜蛋白Amuc_1100在临床前模型中显示出抗肿瘤效果。
工程化的大肠杆菌Nissle 1917菌株被改造用于在肿瘤局部递送免疫刺激分子(如将氨转化为L-精氨酸)或免疫检查点抑制剂纳米抗体,从而在不产生全身毒性的情况下增强抗肿瘤免疫。
微生物组干预策略面临的挑战
– 机制不确定性
许多干预措施(如高纤维饮食、传统益生菌)缺乏对特定菌种或功能如何调节疗效的深入机制理解,导致策略普适化,而非个体化。
– 情境依赖性和个体差异
微生物组的效果受宿主遗传、肿瘤类型、治疗方案和生活方式等多种因素影响,使得干预效果难以一概而论。
– 缺乏个体化生物标志物
目前缺少可靠的生物标志物来指导患者分层,或确定应靶向微生物的哪个方面(如药物代谢、屏障功能或免疫调节)。
– 临床试验设计的局限性
癌症患者常伴有免疫抑制、恶病质或胃肠道毒性,难以依从严格的饮食方案或口服疗法。此外,抗生素的普遍使用也干扰了微生物组的评估和干预。
除了作为治疗靶点,微生物组在癌症的早期检测和风险分层方面也展现出巨大潜力。
早期检测与风险分层
肠道微生物组已被探索作为早期癌症检测的工具,尤其是在结直肠癌领域。多项研究通过比较癌症患者与健康对照的微生物组谱,试图识别可用于诊断的微生物标志物。
谷禾健康通过上万例的结直肠癌和结直肠息肉患者及健康对照构建的肠癌识别模型,在样本量、病例类型覆盖与真实世界差异性方面显著优于多数以“数十到数百例”为主的研究型队列。更大的队列意味着模型能够学习到更稳定、可重复的微生物特征组合,降低因地域、饮食、年龄结构、用药史等因素带来的偶然偏差,从而在实际应用中具备更强的泛化能力与可识别性。
同时,许多研究仅做“癌症 vs 健康”的二分类;谷禾模型纳入“结直肠息肉”等癌前人群,更贴合早筛需求,能在症状出现前识别风险,并捕捉从健康到息肉/腺瘤再到癌变的连续变化信号,从而支持早期风险分层与更有针对性的随访与干预。
谷禾结直肠癌模型示意图
编辑
<来源:谷禾肠道菌群检测数据库>
但是不管是谷禾的模型还是科学研究的结果直接引用于临床诊断还需要有一段路走。
首先,横断面研究难以区分因果关系和相关性,且易受混杂因素(如并存病、用药)的干扰。
其次,与现有筛查手段(如 FIT/粪便免疫化学、结肠镜、影像与血液指标)进行对照评估,确定其增益价值与适用场景;并建立标准化的采样、测序/检测、质控与解读体系,控制抗生素、饮食、肠道准备等混杂因素影响。
伴 随 诊 断
微生物组作为伴随诊断工具的前景可能更为广阔,可用于指导治疗决策、预测疗效或毒性风险。例如,通过检测治疗前的肠道或肿瘤微生物组,可以识别出对特定疗法(如ICI)反应可能性较高的患者,或发生irAEs风险较高的患者。
基于对瘤胃球菌(Ruminococcus)等丰度的检测,可以对接受ICI治疗的患者进行分层。理想的伴随诊断应植根于机制理解,而不仅仅是关联数据,从而指导个性化干预,例如决定是否需要将FMT作为一线辅助治疗,或是否需要先通过菌群移植恢复患者对膳食纤维的降解能力再进行饮食干预,而不是盲目的粪菌移植。
人工智能(AI)有望通过实现高级数据整合、模式识别和预测建模,彻底改变微生物组研究。
编辑
图源:doi.org/10.1038/s41579-025-01268-6
面对微生物组多组学数据维度灾难(即特征数量远超样本数量)的挑战,AI提供了解决方案。例如,通过降维、合成数据增强或迁移学习等策略,可以提高模型的稳定性和泛化能力。
新兴的AI方法,如变分贝叶斯神经网络和卷积神经网络,能够整合配对的微生物组和代谢组数据,或将多组学数据转换为图像格式进行分析,从而发掘深层生物学关联。
编辑
数字孪生是另一个令人兴奋的方向。这是一个动态的、虚拟的患者复制品,整合了静态数据(如基因信息)和实时数据(如临床记录、组学数据),以模拟、监测和预测健康结果。将微生物组数据纳入数字孪生模型,可以模拟其与宿主和治疗的复杂相互作用,支持假设驱动的干预措施和个性化治疗调整。
为不同生命阶段(如婴儿期和成年期)开发量身定制的数字孪生模型,将为整个生命周期的微生物组靶向干预提供宝贵的指导。
当前临床试验概览
近年来,旨在利用微生物组改善癌症治疗的临床试验数量激增。
FMT试验主要集中于免疫治疗效果良好的癌种(如黑色素瘤、RCC、NSCLC),旨在逆转或改善对ICIs的耐药性(如NCT05286294, NCT04988841)。其他试验则探索FMT在肝癌(NCT05690048)、胰腺癌(NCT06393400)等其他癌种中的应用,或用于减轻治疗相关的毒副反应,如免疫相关性结肠炎(NCT04038619)。
关于益生元、益生菌和后生元的临床试验同样蓬勃发展,多数旨在增强抗癌免疫治疗的效果(如NCT05303493, NCT06250335)。
A.muciniphila(NCT05865730)和C.butyricum等下一代益生菌的专项试验正在进行中。
这些研究不仅关注疗效,也探索其在围手术期管理、预防癌前病变复发以及减轻治疗副作用方面的潜力。
越来越多的证据表明,肠道微生物组是癌症治疗结果的关键调节器,其通过免疫调节、药物代谢和维持上皮屏障完整性等多种机制发挥作用。这一认知推动了新型诊断和治疗策略的开发,但临床转化仍受制于机制不确定性、缺乏个性化生物标志物以及在癌症人群中实施的挑战。
微生物组研究揭示,宿主与微生物群作为一个整合系统运作,治疗结果源于宿主-微生物-药物三者间的动态相互作用。
功能冗余、微生物可塑性和强烈的情境依赖性使得因果推断变得困难,也可能解释了为何微生物标志物在不同研究中难以重复。当前的许多干预措施基于普适性策略而非深刻的机制理解,而个体差异进一步加大了个性化治疗的复杂性。
为了推动该领域的发展,未来的研究必须转向由AI赋能、以机制为导向的精准干预,这需要以可靠的伴随生物标志物为支撑,并通过大规模、纵向的实用性临床试验证实。至关重要的是,临床试验必须考虑癌症患者的特殊限制,并在真实世界环境中关注有临床意义的功能性终点。随着微生物组科学日益融入肿瘤学,它有望通过将机制精度与临床相关性相结合,重新定义癌症治疗,实现更有效、更耐受的疗法。
注:本账号内容仅供参考和学术探讨,不构成医疗诊断或治疗建议。任何健康问题,请务必咨询专业医生。
主要参考文献
Hajjar R, Mars RAT, Kashyap PC. Harnessing the microbiome for cancer therapy. Nat Rev Microbiol. 2026 Jan 5.
Nobels A, van Marcke C, Jordan BF, Van Hul M, Cani PD. The gut microbiome and cancer: from tumorigenesis to therapy. Nat Metab. 2025 May;7(5):895-917.
de Vos, W. M., Tilg, H., Van Hul, M. & Cani, P. D. Gut microbiome and health: mechanistic insights. Gut 71, 1020–1032 (2022).
Domzaridou, E. et al. The impact of oral antibiotics prior to cancer diagnosis on overall patient survival: findings from an English population-based cohort study. Curr. Oncol. 30, 8434–8443 (2023).
Yuan, L. et al. The influence of gut microbiota dysbiosis to the efficacy of 5-fluorouracil treatment on colorectal cancer. Biomed. Pharmacother. 108, 184–193 (2018).
Routy, B. et al. Gut microbiome influences efficacy of PD-1-based immunotherapy against epithelial tumors. Science 359, 91–97 (2018).
Elkrief, A. et al. Antibiotics are associated with worse outcomes in lung cancer patients treated with chemotherapy and immunotherapy. NPJ Precis. Oncol. 8, 143 (2024).
Ransohoff, J. D. et al. Antimicrobial exposure is associated with decreased survival in triple-negative breast cancer. Nat. Commun. 14, 2053 (2023).
Cong, J. et al. Bile acids modified by the intestinal microbiota promote colorectal cancer growth by suppressing CD8+ T cell effector functions. Immunity 57, 876–889.e11 (2024).
Tintelnot, J. et al. Microbiota-derived 3-IAA influences chemotherapy efficacy in pancreatic cancer. Nature 615, 168–174 (2023).
Bender, M. J. et al. Dietary tryptophan metabolite released by intratumoral Lactobacillus reuteri facilitates immune checkpoint inhibitor treatment. Cell 186, 1846–1862.e26 (2023).
Salminen, S. et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 18, 649–667 (2021).
谷禾健康
普通拟杆菌(Bacteroides vulgatus/Phocaeicola vulgatus)是人类结肠中最常见的革兰阴性厌氧杆菌之一,在超过90%的人群中都能够检出。其属于拟杆菌属(Bacteroides),是肠道“核心菌”,既有益生潜力,也在某些条件下与疾病相关。
它具有大量多糖利用位点(PULs),能降解膳食多糖和黏液糖蛋白,产生短链脂肪酸(如乙酸、丙酸、丁酸)和琥珀酸等有机酸。基因组学显示其具有丰富的碳水化合物转运与代谢系统。研究发现,其会产生一种叫做泛酸盐(维生素B5)的物质,影响肠道中的GLP-1和肝脏的FGF21的释放,这两种激素会影响葡萄糖代谢并具有神经保护特性。
B.vulgatus产生的代谢物(如N-乙酰氨基葡萄糖)可以促进另一个肠道有益细菌Akkermansia muciniphila(简称Akk菌)的生长。在高脂饮食导致Akk菌数量减少的情况下,B.vulgatus的施用可以恢复其丰度。
研究发现,普通拟杆菌(B.vulgatus)在肥胖、心血管疾病、2型糖尿病、肠道炎症、内皮功能障碍患者中丰度降低;而在自闭症和甲状腺炎患者中发现其丰度升高。
总的来说,普通拟杆菌(Bacteroides vulgatus) 算是一种有益的肠道共生菌,但其效果依赖于特定菌株的水平特异性。其在心血管代谢病、糖尿病、骨质疏松、结肠炎及感染防御等领域中显示出显著的保护潜力,同时在特定菌株或蛋白酶过度表达、黏膜屏障破坏时又可参与炎症和严重感染。临床应用需基于明确的安全性和作用机制的精准菌株,而不是泛指“B.vulgatus”补充。
普通拟杆菌(Bacteroides vulgatus/Phocaeicola vulgatus)是一种厌氧、革兰氏阴性杆菌。它是人类肠道微生物组中最常见的细菌之一,该菌不仅对维持肠道健康至关重要,还具备多种益生潜力,并与多种疾病(如炎症性肠病和心血管疾病)相关。
分类信息
门:拟杆菌门
纲:拟杆菌纲
目:拟杆菌目
科:拟杆菌科
属:拟杆菌属
物种:普通拟杆菌(Bacteroides vulgatus)
普通拟杆菌(Bacteroides vulgatus)属于拟杆菌门(Bacteroidota)、是Bacteroides/Phocaeicola 属的重要种。
普通拟杆菌(Bacteroides vulgatus)又称Phocaeicola vulgatus,2019年起基于全基因组系统发育分析的研究将若干原属 Bacteroides 的物种重划入新属或其它属,Phocaeicola 作为新属或被恢复的属用于容纳包括 Bacteroides vulgatus 在内的若干物种。
形态与生理
细胞形态:Bacteroides vulgatus为杆状菌,专性厌氧,且不具芽胞形成能力。
抗原性:该菌有多个抗原类型,可能影响其致病性和免疫应答。
• 重要结构与分子特征
脂多糖(LPS):其LPS为低酰化、低磷酸化结构,以四、五酰型为主,与大肠杆菌六酰型脂多糖相比,引发更弱的TLR4促炎反应,具有明显免疫调节/低炎性特征。
蛋白酶与其他酶:具多种蛋白酶和糖苷水解酶,如 GH123 家族的 N-乙酰-β-半乳糖胺苷酶,参与黏膜糖链利用;部分菌株分泌的蛋白酶与溃疡性结肠炎严重度相关。
生态位
Bacteroides vulgatus是人类肠道的常驻共生菌,主要栖息在结肠中,谷禾数据库中显示其人群检出率高达98%。并能通过分解复杂的植物碳水化合物和膳食纤维来产生短链脂肪酸等代谢物。
编辑
代谢能力
Bacteroides vulgatus具有大量多糖利用位点(PULs),能降解膳食多糖和黏液糖蛋白,产生短链脂肪酸(如乙酸、丙酸、丁酸)和琥珀酸等有机酸。基因组学显示其具有丰富的碳水化合物转运与代谢系统。
研究还发现,它会产生一种叫做泛酸盐(维生素B5)的物质,帮助身体释放两种重要的激素。肠道中的GLP-1和肝脏的FGF21,这些激素会调节对糖分的渴望。
耐药性
Bacteroides vulgatus通常对广泛使用的抗生素(如青霉素、头孢菌素和氨基糖苷类)耐药,但对甲硝唑、碳青霉烯类以及某些β-内酰胺和β-内酰胺酶抑制剂仍然敏感。其耐药性可能因感染部位而异。
与其他菌的相互作用
• 促进Akk菌的生长
韩国一项双胞胎队列(n=582)研究表明,B.vulgatus可以促进另一个肠道有益细菌Akkermansia muciniphila(简称Akk菌)的生长。
编辑
研究发现,B.vulgatus通过代谢物的产生,尤其是在黏蛋白降解过程中,促进了Akkermansia muciniphila的丰度。B.vulgatus产生的代谢物(如N-乙酰氨基葡萄糖)对Akk菌的生长至关重要。在高脂饮食导致Akk菌数量减少的情况下,B.vulgatus的施用可以恢复其丰度。
噬菌体互作:被温和噬菌体感染后,可通过抑制感受蛋白 TspO 改变转录组并抑制胆汁酸脱缀合,进而影响宿主胆汁酸信号与表型。
与其他菌的互作:
增强:
Akkermansia muciniphila;
Bacteroidales;
Bacteroides;
Odoribacter;
Peptococcaceae.
抑制:
Bifidobacterium
Coriobacteriales
Adlercreutzia
Collinsella
Porphyromonas
Prevotella
Clostridium
Clostridiales incertae sedis
Clostridiales Family XIII. Incertae Sedis
Blautia
Coprococcus
Dorea
Lachnospiraceae
Ruminococcaceae
Ruminococcus
Dialister
Campylobacteraceae
Erysipelotrichaceae
普通拟杆菌(Bacteroides vulgatus)是人类肠道微生物组中一种重要的厌氧细菌,对维持整体健康至关重要。作为常驻菌群的成员,它参与多种关键生理功能,包括营养物质的代谢、维持肠道微环境的平衡,以及通过产生短链脂肪酸来支持肠道黏膜的完整性。
此外,B.vulgatus还能调节免疫反应,有助于抗炎效应,从而帮助防止各种疾病的发生。这种细菌的健康益处不仅限于肠道,研究表明它与心血管健康、代谢平衡及免疫系统的良好运作密切相关。
1
改善血糖控制并减少糖分渴望
2025年1月,江南大学研究团队在《Nature Microbiology》(IF:20.5)发表重要研究成果:游离脂肪酸受体4(Ffar4)通过肠道菌群调控糖类偏好的分子机制。
编辑
• B.vulgatus丰度下降减少GLP-1的分泌
研究表明,Ffar4表达下调与高糖饮食行为相关,其缺失会导致肠道普通拟杆菌(Bacteroides vulgatus)丰度降低及代谢产物泛酸盐(维生素B5)减少。泛酸盐通过促进GLP-1分泌,激活肝脏FGF21释放,进而作用于下丘脑神经环路,抑制糖类摄入欲望并调节能量代谢平衡。该发现揭示了全新的肠-肝-脑轴调控网络,为代谢性疾病的干预提供了潜在治疗靶点。
GLP-1通过让身体在需要时释放胰岛素,减缓食物离开胃的速度,帮助身体。这能保持血糖稳定,帮助控制食欲。FGF21是另一种激素,能增强身体对胰岛素的敏感度,帮助你燃烧更多能量,并减少对含糖食物的渴望。它们共同创造了一个强大的系统,帮助保持新陈代谢和大脑的正常运作。
2
帮助恢复Akk菌,减轻肥胖
刚才前文就有提到,B.vulgatus通过产生的代谢物(如N-乙酰氨基葡萄糖),促进了Akkermansia muciniphila的丰度。
编辑
You HJ,et al.Gastroenterology.2023
• 改善了Akk菌的耗竭,减轻了肥胖体重
研究发现,Bacteroides vulgatus 与 Akkermansia muciniphila 在瘦受试者(BMI<25)中存在互作关系,但在肥胖群体(BMI≥25)中这种互作消失。补充普通拟杆菌(Bacteroides vulgatus SNUG 40005)显著改善了高脂饮食小鼠的肥胖、胰岛素抵抗和肠道通透性,并通过产生代谢物N-乙酰葡萄糖胺提升了肥胖小鼠中Akkermansia的丰度。
研究者从健康成年人肠道中分离出 Bacteroides vulgatus SNUG40005,并将其灌胃给高脂饮食的肥胖小鼠,结果发现该菌株能显著降低小鼠体重、改善糖耐量并减少血液中脂多糖水平。
// 结论
综合以上结果,Bacteroides vulgatus 通过调节肠道微生物的相互作用,特别是促进Akkermansia muciniphila 的生长,可能对肥胖和代谢健康产生积极影响。这些发现为发展新型治疗方法提供了依据,尤其在管理肥胖和相关代谢疾病方面。
3
抑制动脉粥样硬化
冠状动脉疾病(CAD)患者的肠道微生物谱分析显示,与无冠状动脉风险因素的对照组相比,Bacteroides vulgatus和Bacteroides dorei的丰度显著降低。证实Bacteroides vulgatus和Bacteroides dorei通过降低血液内毒素水平改善动脉粥样硬化小鼠体的症状。
• B.vulgatus与脂多糖水平负相关,影响心血管健康
并且在CAD患者中,B.vulgatus和B.dorei的丰度与粪便脂多糖水平呈负相关,暗示这两种细菌可能通过调控脂多糖的产生来影响动脉粥样硬化的进展。
总之,Bacteroides vulgatus 在调节肠道微生物群、抑制炎症反应及维护心血管健康方面发挥着重要作用。进一步的研究可能帮助开发基于微生物群的策略,用于防治心血管疾病。
4
改善脂质代谢并调节高血脂
Bacteroides vulgatus Bv46 是一种常见的肠道细菌,其在高脂血症(高血脂)中的潜在健康效益受到研究。该研究通过高脂饮食诱导大鼠模型,评估了B.vulgatus Bv46 的补充对体重、脂质代谢、炎症标志物及肝脏病理的影响。
编辑
• 关键发现:
改善脂质代谢:补充B.vulgatus Bv46 显著降低了高脂饮食大鼠的体重增加、食物效率以及肝脏指数,改善了血清脂质状况,包括降低总胆固醇(TC)、甘油三酯(TG)和低密度脂蛋白胆固醇(LDL-C),并提高了高密度脂蛋白胆固醇(HDL-C)的水平。
调节炎症反应:补充后,血清中的促炎细胞因子(如 IL-1β、IL-6 和 TNF-α)显著降低,表明B.vulgatus Bv46 能够减轻全身炎症反应。
肠道微生物的改变:该菌株的补充导致肠道微生物群落组成的显著变化,主要表现为Firmicutes与Bacteroidetes 的比率降低,Bacteroides 和 Parabacteroides 属的丰度增加,这种变化与健康个体的微生物群落特征相似。
短链脂肪酸的产生:补充B.vulgatus Bv46促进了肠道内短链脂肪酸(SCFAs),特别是丙酸和丁酸的产生,这些代谢产物对抗炎和代谢健康具有重要作用。
胆汁酸代谢:研究发现,B.vulgatus Bv46 产生的胆盐水解酶(BSH)有助于胆汁酸的去结合,促进了胆汁酸的转化和排出,从而改善了胆固醇代谢。
代谢组学影响:血清代谢物分析显示,补充B.vulgatus Bv46 能够增强氨基酸、甘油磷脂的代谢,并对脂质代谢产生正面影响。
// 结论
Bacteroides vulgatus Bv46 通过多种机制(包括改善血脂谱、减轻全身性炎症、调节肠道微生物群、增强短链脂肪酸产生和胆汁酸代谢)证明了其在高脂血症干预中的潜力。这些发现不仅支持了B.vulgatus在宿主代谢调节中的作用,也为高脂血症的治疗提供了新的潜在策略。
5
减轻肠道炎症
• B.vulgatus有助于减轻结肠炎症状和病理损伤
有研究探讨了B.vulgatus在小鼠模型中对结肠炎的保护作用及其通过调节肠道微生物群和免疫反应的机制。
编辑
研究发现,B.vulgatus的补充显著减轻了小鼠的结肠炎症状,包括减轻体重下降、改善粪便形态以及减少结肠病理损伤。这表明该菌株在缓解肠道炎症方面具有效果。
此外,B.vulgatus的使用还导致结肠中促炎细胞因子TNF-α、IL-1β和IL-6的表达下降,并可能通过短链脂肪酸(SCFAs)释放实现抗炎效果。
• B.vulgatus可能作为治疗溃疡性结肠炎的靶点
编辑
另一项溃疡性结肠炎(UC)患者的多组学分析研究发表于《Nature Microbiology》,研究发现Bacteroides vulgatus来源的蛋白酶是破坏UC患者肠道屏障的一个原因,体外和体内实验均证实抑制该蛋白酶可降低小鼠结肠炎的发生。Bacteroides vulgatus分泌的蛋白酶可做为治疗UC的潜在靶点。
• 保护急性肠道损伤
还有研究探讨了不同Bacteroides vulgatus菌株对脂多糖(LPS)诱导的急性肠道损伤的保护作用。
从健康的人类粪便样本中分离出七个B.vulgatus菌株,分别命名为FTJS7K1、FTJS5K1、FSDTA11B14、FJSWX62K35、FSDLZ51K1、FBJS10K3和FGSZY37K4。
编辑
研究发现,B.vulgatus FTJS7K1的施药能保护肠道上皮的完整性。B.vulgatus FTJS7K1还抵消了LPS诱导的结肠组织细胞因子mRNA表达变化,以及肠系膜淋巴结中调节T细胞比例的变化。
与LPS组相比,FTJS7K1 组的B.vulgatus增加了乳杆菌、阿克曼氏菌和双歧杆菌的丰度,而 Faecalibaculum 的丰度则下降。此外,B.vulgatus FTJS7K1 组的粪便样本中短链脂肪酸(SCFAs)浓度显著增加。基因组分析表明,FTJS7K1 的保护作用可能通过与防御机制和代谢相关的特定基因介导(例如 SCFA 的分泌)。
6
在骨骼健康中存在矛盾作用
有研究探讨了Bacteroides vulgatus在骨骼健康中的作用,但结果存在矛盾。例如一项研究分析了该菌株在去除卵巢小鼠模型中改善结肠微生物群失调和减轻骨质流失的潜力。
• B.vulgatus在去除卵巢小鼠中减轻骨质流失
研究发现补充B.vulgatus显著减轻了去卵巢小鼠的椎体骨质流失。这一发现表明,该菌株可能在雌激素缺乏引起的骨量减少中发挥保护作用。
B.vulgatus的施用还改善了去卵巢小鼠的结肠微生物群组成,增加了有益菌的丰度,特别是 Lactobacillus 和 Bifidobacterium 属;这些变化与骨质改善的效果相吻合。
• 下调血清戊酸、骨吸收增加
然而,在另外一项研究中发现:B.vulgatus的丰度增加会影响血清戊酸(VA)的水平,而这与骨密度呈正相关。喂食B.vulgatus的卵巢切除小鼠骨吸收增加,骨微结构变差,而喂食VA的小鼠则骨吸收减少,骨微结构改善。
总体而言,Bacteroides vulgatus对骨骼健康的影响并不是简单的有益或有害,而是取决于宿主的生理状态(如激素水平)。在雌激素缺乏的情况下,B.vulgatus的增多可能导致骨密度降低,而在改善肠道健康及抑制炎症的情况下,可能展现出一定的保护作用。因此,针对不同人群或临床情况,应谨慎评估B.vulgatus的治疗潜力。
B.vulgatus丰度过低与疾病
结合上文内容,总体来看,现有研究表明,B.vulgatus的丰度在这些疾病中普遍偏低,且这种低丰度可能与具体的健康问题存在潜在的联系。我们在下方的表格中进行了总结:
编辑
B.vulgatus丰度过高与疾病
尽管研究发现 Bacteroides vulgatus 具有多种代谢或健康方面的益处,但同时也观察到其在某些疾病状态下,其丰度异常增高。
①自闭症患者中丰度较高
编辑
研究发现,Bacteroides vulgatus在自闭症儿童中的丰度显著高于健康对照组,暗示其可能与自闭症症状的发展相关。这种细菌可以产生像丙酸这样的SCFA,在高浓度下可能具有神经毒性并影响行为。
②甲状腺炎患者中增加
编辑
在 Hashimoto 甲状腺炎 (HT) 的患者群体中,B.vulgatus 的丰度显著增加,这与肠道微生物群的失衡及免疫反应激活可能相关。
与其他细菌的对比:HT 患者的肠道微生物组相比于健康对照组,显示出整体的细菌多样性和丰度改变,且B.vulgatus在特定条件下的增多可能与其他有害菌(如Escherichia-Shigella)的共存有关。
③可能是婴儿乳糜泻的风险因素
过高水平的 B. vulgatus 还被认为是婴儿乳糜泻的重要风险因素,这意味着当这种细菌在婴儿的肠道中丰度较高时,患上乳糜泻的可能性会增加。
饮食因素
B.vulgatus的种群规模对所摄入的食物高度敏感,因此饮食是影响其丰度的最重要外部因素。
•高纤维饮食:富含植物纤维和抗性淀粉的饮食能够促进B.vulgatus的生长。全谷物、豆类、蔬菜及坚果等食物能够为这种细菌提供营养,帮助其发酵并产生有益的短链脂肪酸(SCFAs),这些代谢产物对维持肠道健康和代谢调节至关重要。洋葱、大蒜、香蕉和芦笋等食物也被发现能够促进B.vulgatus丰度增加。
•发酵食品:酸奶、开菲尔、酸菜、泡菜等发酵食品也有助于支持B.vulgatus的健康生长。
•富含多酚的食物:食用富含多酚的食物,如浆果、绿茶、黑巧克力和红酒。
•减少加工食品摄入:高加工食品通常纤维含量低,会对有益细菌产生负面影响,导致肠道菌群不平衡,进而抑制 B. vulgatus 的生长。
生活方式因素
•日常锻炼:规律的身体活动有助于增强肠道微生物的多样性,从而支持B.vulgatus的丰度。
•压力管理:持续的心理压力可能对肠道健康产生负面影响。采用减压措施(如冥想、瑜伽和正念练习)有利于维护肠道微生物的平衡。
•健康的睡眠模式:保持规律的睡眠,以支持肠道微生物群的平衡。
药物与补充剂
•益生菌:考虑直接补充益生菌以增加肠道中的B.vulgatus。
•避免不必要的抗生素:仅在必要时使用抗生素,以避免破坏肠道微生物群。
主要参考文献:
quez-Baeza Y, Sauceda C, Daniel N, Gerner RR, Batachari LE, Malfavon M, Zhu Q, Weldon K, Humphrey G, Carrillo-Terrazas M, Goldasich LD, Bryant M, Raffatellu M, Quinn RA, Gewirtz AT, Chassaing B, Chu H, Sandborn WJ, Dorrestein PC, Knight R, Gonzalez DJ. Multi-omics analyses of the ulcerative colitis gut microbiome link Bacteroides vulgatus proteases with disease severity. Nat Microbiol. 2022 Feb;7(2):262-276.
Yoshida N, Emoto T, Yamashita T, Watanabe H, Hayashi T, Tabata T, Hoshi N, Hatano N, Ozawa G, Sasaki N, Mizoguchi T, Amin HZ, Hirota Y, Ogawa W, Yamada T, Hirata KI. Bacteroides vulgatus and Bacteroides dorei Reduce Gut Microbial Lipopolysaccharide Production and Inhibit Atherosclerosis. Circulation. 2018 Nov 27;138(22):2486-2498.
Wang C, Xiao Y, Yu L, Tian F, Zhao J, Zhang H, Chen W, Zhai Q. Protective effects of different Bacteroides vulgatus strains against lipopolysaccharide-induced acute intestinal injury, and their underlying functional genes. J Adv Res. 2021 Jun 15;36:27-37.
Xu M, Lan R, Qiao L, Lin X, Hu D, Zhang S, Yang J, Zhou J, Ren Z, Li X, Liu G, Liu L, Xu J. Bacteroides vulgatus Ameliorates Lipid Metabolic Disorders and Modulates Gut Microbial Composition in Hyperlipidemic Rats. Microbiol Spectr. 2023 Feb 14;11(1):e0251722.
Liu L, Xu M, Lan R, Hu D, Li X, Qiao L, Zhang S, Lin X, Yang J, Ren Z, Xu J. Bacteroides vulgatus attenuates experimental mice colitis through modulating gut microbiota and immune responses. Front Immunol. 2022 Nov 30;13:1036196.
Yuan S, Shen J. Bacteroides vulgatus diminishes colonic microbiota dysbiosis ameliorating lumbar bone loss in ovariectomized mice. Bone. 2021 Jan;142:115710.
Li S, Wang C, Zhang C, Luo Y, Cheng Q, Yu L, Sun Z. Evaluation of the Effects of Different Bacteroides vulgatus Strains against DSS-Induced Colitis. J Immunol Res. 2021 May 29;2021:9117805.
Lin X, Xiao HM, Liu HM, Lv WQ, Greenbaum J, Gong R, Zhang Q, Chen YC, Peng C, Xu XJ, Pan DY, Chen Z, Li ZF, Zhou R, Wang XF, Lu JM, Ao ZX, Song YQ, Zhang YH, Su KJ, Meng XH, Ge CL, Lv FY, Luo Z, Shi XM, Zhao Q, Guo BY, Yi NJ, Shen H, Papasian CJ, Shen J, Deng HW. Gut microbiota impacts bone via Bacteroides vulgatus-valeric acid-related pathways. Nat Commun. 2023 Oct 27;14(1):6853.

谷禾健康
《植物性饮食》
植物性饮食能提供丰富的膳食纤维、多酚和其他生物活性化合物,适当和健康的植物性饮食能够促进有益肠道细菌的生长,增加菌群多样性和短链脂肪酸(SCFAs)的产生,有助于改善肠道屏障功能、减少炎症并降低慢性疾病风险。
然而,值得注意的是,植物性饮食也可能通过生食和未加工食品将环境微生物引入肠道,其中一些微生物可能是机会致病菌或携带抗生素抗性基因,这对健康构成潜在威胁。
此外,健康与不健康植物性饮食的区别也很关键。富含全谷物、水果、蔬菜和豆类的饮食与更有利的肠道菌群组成和代谢结果相关,而富含精制碳水化合物和加工食品的植物性饮食可能产生不利健康的效果。
本文一起了解下植物性饮食对肠道菌群的影响和塑造,尤其在多样性方面,以便更好地理解和实践植物性饮食,在享受植物性饮食带来的健康益处的同时,避免可能的健康风险。
植物性饮食主要由植物来源的食物组成。根据对动物产品的限制程度,可分为不同类型:
– 纯素饮食 (vegan)
仅包含植物性高纤维食物,如水果、蔬菜、谷物、豆类、坚果和种子,完全排除肉类、家禽、鱼类、乳制品、蛋类及所有动物副产品。
– 素食饮食 (vegetarian)
排除肉类和肉类衍生食品,但可能包含鸡蛋或乳制品等动物产品。
– 弹性素食 (flexitarian)
主要以植物性食物为主,但偶尔包含少量肉类或鱼类。
植物性饮食增加多样性和有益菌
研究表明,富含多种植物性食物的饮食始终与更高的肠道微生物多样性相关。总体而言,从植物中引入有益细菌,加上植物纤维,可能培养出更具弹性和功能多样性的肠道生态系统,最终可能导致人类更好的健康结果。
肠道微生物通过发酵促进消化
因为生活在结肠中的肠道微生物通过发酵胃和肠道酶无法降解的食物物质,对消化做出重要贡献。富含植物性食物的饮食能显著改善有益肠道菌群,直接影响人类健康。
最近的研究结果强调,植物性饮食富含益生元,如膳食纤维(包括抗性淀粉),它们作为有益肠道微生物的能量来源。
特定细菌发酵膳食纤维生成SCFAs
膳食纤维是一类植物性碳水化合物,包括非淀粉多糖、木质素和抗性淀粉,它们能逃脱人类上消化道内源性酶的消化。只有特定的肠道菌群,如F. prausnitzii、Roseburia、双歧杆菌和普雷沃氏菌,能在厌氧条件下发酵膳食纤维,产生SCFAs(如乙酸、丙酸和丁酸)作为主要代谢产物。
编辑
一项系统综述显示,纯素和素食饮食通过增加有益细菌同时减少有害细菌,在重塑肠道菌群方面发挥关键作用。在食用植物性饮食的个体中,观察到F. prausnitzii、Roseburia spp.、Coprococcus spp.和Ruminococcus spp.的水平增加,这些细菌都能产生SCFAs,促进更好的肠道健康、减少炎症和改善新陈代谢。
研究还发现普雷沃氏菌属的丰度增加,在某些情况下还有脆弱拟杆菌(Bacteroides fragilis),它们在纤维消化和免疫调节中起关键作用。但是要注意,不合理的植物性饮食也会使得普雷沃氏菌属或有些情况下的脆弱拟杆菌过度增殖,影响菌群平衡和进一步的消化免疫状况。
短链脂肪酸:肠道菌群的黄金代谢物
短链脂肪酸(SCFAs)已被广泛研究为关键的肠道菌群代谢物。人类由于缺乏必要的酶而无法发酵某些碳水化合物,而肠道菌群则能合成诸如丙酸-CoA转移酶和丙醛脱水酶等酶,将这些碳水化合物转化为SCFAs。
胃肠道中的主要SCFAs是乙酸、丙酸、丁酸(占总量的≥95%):
乙酸
通过Wood-Ljungdahl途径和乙酰-CoA途径合成,相关细菌包括乳酸杆菌属、双歧杆菌属、 Akkermansia muciniphila、拟杆菌属、普雷沃氏菌属、瘤胃球菌属、链球菌属等。
丙酸
由Phascolarctobacterium succinatutens、拟杆菌属、Dialister、Megasphaera elsdenii、韦荣球菌属、AKK、Coprococcus catus、Roseburia inulinivorans、Ruminococcus obeum、沙门氏菌属等通过琥珀酸、丙烯酸和丙二醇途径生成。
丁酸
主要由Faecalibacterium prausnitzii、Roseburia、Eubacterium rectale、Clostridium leptum、Eubacterium hallii、Coprococcus eutactus、Anaerostipes caccae、C. catus等通过丁酰-CoA:乙酸CoA转移酶和磷酸丁酰转移酶/丁酸激酶途径产生。
短链脂肪酸在维持肠道屏障完整性、调节免疫反应和支持代谢健康方面发挥着关键作用,因此与纯素和素食饮食相关的独特肠道菌群特征有助于产生抗炎作用和改善代谢结果。
并非所有植物性食物都同样健康
一些植物性食物可能增加健康风险。为解决这些问题,研究人员创建了:
另一项研究发现,较高的hPDI与Bacteroides cellulosilyticus和Eubacterium eligens的丰度增加相关,这两种细菌都有助于积极的代谢健康。
相反,较高的hPDI显示Ruminococcus torques和Ruminococcus gnavus的水平降低,这些细菌种类已知会引起炎症反应和不利的代谢结果。
高hPDI饮食促进有益微生物,降低代谢风险
高hPDI还与肠道微生物途径的富集相关,这些途径参与支链氨基酸(包括L-异亮氨酸和L-缬氨酸)的生物合成,以及丙酮酸发酵生成异丁醇,这些都与较低的代谢风险评分相关。相反,低hPDI与嘌呤碱基降解和脂质生物合成相关的途径相关,这与不太有利的代谢特征相关。
这些结果表明,坚持健康的植物性饮食能促进有益的肠道菌群和代谢健康,而不健康的植物性饮食可能产生相反的影响。
植物性饮食与肠道菌群的关系
一项大规模调查研究了杂食者、素食者和纯素者的肠道菌群特征,分析了来自5个独立全球队列的21,561人的数据。
研究表明,植物性饮食驱动肠道菌群组成,有利于参与纤维发酵和SCFAs合成的微生物物种。与杂食者相比,纯素饮食的人总体肠道微生物多样性较低,但有益物种(如毛螺菌科、Butyricicoccus spp.和Roseburia hominis)的较高,这些物种有助于减少炎症和增强肠道健康。
研究局限与未来方向
然而,该研究也存在局限性:参与者被分为杂食者、素食者或纯素者时,没有考虑饮食质量,例如全食物纯素与加工纯素的区别。尽管使用了健康植物性饮食指数(hPDI),但它没有考虑纤维摄入量的差异,而纤维摄入量可能对肠道菌群产生不同影响,并可能为积极的代谢健康提供机制见解。
大多数关于植物性饮食对肠道菌群影响的研究持续时间较短,从几天到13个月不等。虽然短期饮食干预可以快速改变微生物组成,但它们可能无法提供长期稳定性和更广泛健康影响的完整图景。未来需要更多长期研究来探索植物性饮食对肠道菌群的持久影响。
多酚:植物中的抗氧化战士
植物性饮食富含多种植物生物活性化合物,包括膳食纤维、多酚、类胡萝卜素、维生素和生物活性肽,这些化合物选择性地促进有益肠道细菌的生长和活性,从而有助于宿主健康。
据观察,富含叶菜类蔬菜、浆果、豆类、小扁豆和燕麦等植物性食物的饮食支持共生细菌的生长并提高SCFAs水平。
编辑
多酚是一类广泛而多样的天然化合物,大量存在于植物中,以其强大的抗氧化和抗炎特性为特征。它们可大致分为类黄酮和非类黄酮(酚酸和芪类)。
大多数膳食多酚在小肠中不被吸收,而是在结肠中进行微生物代谢,产生生物活性代谢物如尿石素,这些代谢物随后可被吸收到体内,并通过影响代谢、免疫甚至神经通路(通常通过复杂的肠-肝-脑轴)发挥显著的健康益处。
这些化合物似乎能增加人类肠道中益生菌如乳酸杆菌和双歧杆菌的丰度。据报道,来自浆果和蔓越莓等水果的多酚对梭状芽胞杆菌和大肠杆菌等致病菌具有抗菌活性,同时富集有益菌。
一些动物和人类研究表明,多酚可能将厚壁菌/拟杆菌比率转向更健康的平衡,这种平衡在肥胖和代谢紊乱中常常被破坏。通过增加SCFA水平和调节胆汁酸途径,多酚可以改善肠道屏障完整性,减少炎症,并支持代谢健康。
类胡萝卜素:未被充分探索的肠道调节剂
膳食纤维和多酚是研究最广泛的用于调节肠道菌群和促进肠道健康的生物活性化合物。相比之下,其他生物活性化合物如类胡萝卜素(β-胡萝卜素、番茄红素、虾青素、叶黄素、玉米黄质和岩藻黄质)在这方面的研究仍不充分。
类胡萝卜素具有抗炎和抗氧化特性,研究表明它们可能促进有益肠道细菌,增强肠道屏障功能,减少炎症。然而,大多数证据来自动物研究,剂量、干预持续时间各不相同,且对SCFAs产生的评估有限。
这突出了需要精心设计的人类干预研究,以更好地了解类胡萝卜素对肠道健康的影响。未来的研究应侧重于标准化的人体试验和综合多组学方法,以阐明特定类胡萝卜素和其他生物活性化合物影响肠道微生物多样性、屏障功能和宿主健康结果的机制。
不同植物性食物对肠道菌群的独特影响
不同的植物性食物含有独特的生物活性化合物组合,因此对肠道菌群有不同的影响:
-浆果类
富含花青素等多酚,促进双歧杆菌和乳酸杆菌生长。
-全谷物
含有丰富的膳食纤维和抗性淀粉,促进产丁酸菌如Faecalibacterium prausnitzii和Roseburia spp.
-豆类
含有棉子糖家族低聚糖,促进双歧杆菌增殖
-坚果和种子
富含不饱和脂肪酸、膳食纤维和多酚,增加 Akkermansia muciniphila等有益菌
-十字花科蔬菜
部分研究显示有调节拟杆菌丰度的潜力,含有硫代葡萄糖苷,经肠道菌群代谢为具有抗癌活性的异硫氰酸酯。
因此,多样化的植物性饮食能够提供更广泛的生物活性化合物,支持更丰富和多样化的肠道菌群,从而带来更多健康益处。
除了提供营养价值和塑造肠道菌群的生物活性次生代谢物外,植物性饮食还作为影响微生物多样性的来源。最近的研究首次提供了水果和蔬菜微生物群可能传播到人类肠道的证据。
在这项研究中发现的人类肠道与水果/蔬菜之间的共享微生物包括假单胞菌(Pseudomonas)、鞘氨醇单胞菌(Sphingomonas)、气单胞菌(Aeromicrobium)和泛菌(Pantoea)。
尽管这些细菌中的一些具有促进植物生长的特性,但它们可能在人类肠道中引起感染,特别是在免疫功能低下的个体中。
通过植物性饮食将土壤微生物群转移到人类肠道
编辑
图示显示微生物从土壤转移到植物,最后植物性饮食促进植物微生物群直接转移到人类肠道。
另外一项采用物种水平鉴定的跨国队列研究发现,食用植物性饮食的个体,特别是纯素者,肠道中通常与土壤和植物相关的微生物(包括Enterobacter hormaechei、Citrobacter freundii、Raoultella ornithinolytica和肺炎克雷伯菌)的较高。这是由于摄入更多的生的和最低限度加工的植物性食物,这些食物携带环境微生物。
编辑
在农业环境中,这些细菌通过溶解磷等营养物质、固氮、产生植物激素(包括吲哚-3-乙酸)以及作为植物病原体的生物防治剂,发挥着至关重要的作用。另一方面,它们在人类中可能是共生菌或机会致病菌,引起从尿路感染到败血症的广泛感染。这些细菌还携带抗菌素耐药基因,导致对抗生素的强烈耐药性。
下面总结了植物微生物在植物中的作用
和对人类的可能潜在风险(仅供参考)
编辑
我们食物中的微生物群落可能受到不同食品加工方法和更广泛的农业实践的影响。微生物从植物到人类的传播表明,植物及其土壤环境可以作为这些细菌的储存库。食用未加工或最低限度加工的植物性饮食可能是将共生菌或机会致病菌转移给人类的潜在载体。烹饪可能是使食物中天然存在或通过农业实践引入的抗菌化合物变性的途径。
小鼠模型研究:生食与熟食的微生物差异
一项对小鼠模型的研究表明,小鼠食用生的或熟的植物性饮食后,肠道菌群存在显著差异。根据饮食的不同,小鼠的肠道细菌负荷有所不同;食用生块茎的组比食用熟块茎的组细菌负荷更低。这种微生物负荷的下降与微生物细胞损伤增加和微生物活性降低有关。
人体研究
一项短期人体研究检查了在生食和熟食植物性饮食之间切换如何影响肠道菌群组成。结果表明,食用熟食后48小时内,肠道菌群发生了显著变化。食用生食和熟食的个体之间细菌类群丰度的差异突出了食品加工对肠道菌群结构和功能的影响。
农业实践通过减少整体多样性和选择性地支持化学/抗生素抗性细菌物种,改变土壤微生物组成。农业环境中的抗生素耐药性主要由肥料、废水等的应用驱动。
肥料作为抗生素耐药细菌的储存库
农业中广泛使用肥料是抗生素耐药细菌及其抗生素抗性基因的主要储存库。根际、内生圈和叶际内的微生物群落密切相关,形成一个统一的生态系统,对人类食物链至关重要。
抗生素耐药细菌在植物中的传播及其对人类的影响
抗生素耐药细菌可以侵入植物根部,进入茎和叶组织,或与内生菌交换抗性基因,促进植物内抗生素抗性的扩展。
人类通过食用可食用植物获得这些抗性基因。长期摄入这些抗性基因可能与人类致病性增加有关,因为这些基因可能通过水平基因转移被肠道的一些机会致病菌获得。然而,对与植物相关的抗生素耐药组及其向人类转移的理解仍处于初级阶段。
这些发现强调了土壤、植物和人类肠道中微生物群落的相互联系。因此,维持土壤健康对于健康的植物至关重要,而健康的植物又有助于平衡的肠道菌群。
多篇研究警告说,缺乏适当饮食规划的植物性饮食可能导致关键营养素(如维生素B12、维生素D、铁、钙、锌、二十碳五烯酸(EPA)和二十二碳六烯酸(DHA))的缺乏,可能损害肠道和整体健康。
考虑到遗传因素和微生物多样性的差异,即将进行的研究应阐明植物性饮食对个体肠道和代谢健康的影响。
营养建议
对于遵循植物性饮食的人,建议关注以下营养素的摄入:
-维生素B12
主要存在于动物性食物中,植物性饮食者应考虑强化食品或补充剂
-铁
植物性来源的铁(非血红素铁)吸收率较低,建议与富含维生素C的食物一起食用以提高吸收率
-钙
可从强化植物奶、豆腐(用钙盐制作)、绿叶蔬菜中获取
-Omega-3脂肪酸
可从亚麻籽、奇亚籽、核桃中获取ALA,以及其他富含DHA/EPA等食物
-锌
全谷物、豆类、坚果是主要来源,发酵食品可提高锌的生物利用率
编辑
✦ 如何开始并坚持健康的植物性饮食?
对于想要尝试植物性饮食的人,建议循序渐进:
-从餐盘开始
将餐盘的一半分配给蔬菜和水果,四分之一给全谷物,四分之一给植物蛋白来源
-增加多样性
尝试不同种类的植物性食物,每周至少尝试一种新的水果、蔬菜或全谷物
-注重全食物
优先选择完整的、未加工的食物,减少精制食品和添加糖
-合理搭配
结合不同植物蛋白来源(如豆类和谷物)以获得完整的氨基酸谱
-适当烹饪
虽然生食有其益处,但适当烹饪可以减少潜在病原体风险,同时提高某些营养素的生物利用率
✦ 平衡植物性饮食与食品安全
为了最大限度地发挥植物性饮食的益处同时减少风险:
-清洗生食
彻底清洗所有生水果和蔬菜,去除表面的土壤和微生物
-适当烹饪
对可能含有较多环境微生物的食物(如芽菜、生豆类)进行适当烹饪
-注意高风险人群
免疫功能低下者应格外小心生食,考虑咨询医生或营养师
-多样化来源
从不同来源购买农产品,减少单一来源的风险
-考虑有机选择
在可能的情况下,选择有机种植的农产品,减少农药暴露
编辑
✦ 个性化植物性饮食策略
每个人的肠道菌群都是独特的,因此对植物性饮食的反应也可能不同。建议:
通过这些策略,我们可以充分利用植物性饮食对肠道菌群和整体健康的益处,同时最大限度地减少潜在风险,踏上个性化的健康之旅。
主要参考文献
Anand R, Sahil R, Jain M, Maurya GK, Kharat AS. Plant-Based Diet as a Precursor to Human Gut Diversity. J Nutr. 2025 Nov 26:101251.
Fackelmann G, Manghi P, Carlino N, Heidrich V, et al., Gut microbiome signatures of vegan, vegetarian and omnivore diets and associated health outcomes across 21,561 individuals. Nat Microbiol. 2025 Jan;10(1):41-52.
Ramaboli MC, Ocvirk S, Khan Mirzaei M, et AL., Diet changes due to urbanization in South Africa are linked to microbiome and metabolome signatures of Westernization and colorectal cancer. Nat Commun. 2024 Apr 20;15(1):3379.

谷禾健康
精神分裂症(SCZ)是一种严重的持续性精神障碍,表现为阳性症状(如妄想和幻觉)、负性症状(如动力丧失和社交退缩)以及认知症状(包括工作记忆和认知灵活性缺失)。此外,大多数精神分裂症患者伴有显著的睡眠障碍(SD),常出现入睡困难、睡眠维持问题、睡眠结构紊乱和昼夜节律失调。
睡眠障碍这种共病会加重精神症状,导致更频繁的偏执意念、幻觉和思维紊乱,以及更高程度的抑郁和焦虑。还显著影响治疗依从性和长期预后。
精神分裂症的发病机制涉及遗传、环境、免疫和神经发育等多种因素的相互作用。与此同时,现有研究表明,肠道微生物群通过微生物群-肠道-大脑轴调节大脑功能,影响神经递质的代谢和免疫炎症反应,因此可能在精神分裂症和睡眠障碍的发生和发展中发挥重要作用。
精神分裂症患者常表现出产丁酸细菌减少、产乳酸菌增加,以及与谷氨酸和γ-氨基丁酸(GABA)代谢相关的细菌增加。特别是,在精神分裂症患者中,这种肠道微生物群破坏可能通过影响神经递质平衡、促进神经炎症和干扰昼夜节律等机制,促成精神症状和睡眠问题的发展。
本文整合了现有研究,首先介绍了精神分裂症(SCZ)和睡眠障碍(SD)患者的肠道微生物群特征。探讨了SCZ和SD中关键微生物群及其代谢物的共同影响,以及肠道微生物群通过神经免疫、内分泌和神经递质通路的作用机制。最后,提出了针对肠道微生物群的干预策略在改善精神分裂症伴随SD中的潜在应用。
精神分裂症(SCZ)是一种严重的精神障碍,表现为阳性症状(超出正常范围的行为和思维)如妄想和幻觉;负性症状(缺乏正常的情感和行为)如动力丧失和社交退缩;以及认知症状,包括工作记忆和认知灵活性的缺失。
★ 许多精神分裂症患者伴有睡眠障碍
除了典型的精神症状外,睡眠障碍(SD)是精神分裂症最常见的共病之一,表现为失眠、睡眠片段化、慢波睡眠减少和昼夜节律紊乱。
研究显示,约80%的精神病患者至少患有一种类型的SD。睡眠质量的恶化会加重精神症状,导致更频繁的偏执意念、幻觉和思维紊乱,以及更高程度的抑郁和焦虑。研究发现,睡眠质量受损和昼夜节律紊乱的精神分裂症(SCZ)患者在负性症状评估上得分更高,认知功能障碍更为明显。
此外,SD不仅加重精神分裂症的精神病症状,精神分裂症的核心症状也会破坏睡眠生理。多导睡眠图研究表明,更严重的阳性症状与短的快速眼动(REM)潜伏期、长的入睡潜伏期和低睡眠效率相关,而明显的负性症状与非快速眼动(NREM)睡眠中的慢波振幅降低和REM起始潜伏期缩短有关。
这些发现表明精神分裂症与睡眠障碍之间存在双向病理循环,睡眠障碍不仅是精神分裂症的常见共病,也是影响其症状表达和疾病进展的关键因素。
1
流行病学特征
研究表明,精神分裂症(SCZ)患者的睡眠障碍(SD)患病率显著高于普通人群,影响其生活质量和疾病预后。
精神分裂症患者快速眼动睡眠时间缩短
通过睡眠脑电图(EEG)观察精神分裂症患者与健康对照组的睡眠结构差异发现,精神分裂症患者的快速眼动(REM)潜伏期缩短,REM密度增加,并且REM睡眠比例与症状严重程度密切相关。在非快速眼动(NREM)睡眠期间,精神分裂症患者呈现慢波睡眠减少、纺锤体波密度和振幅下降,以及持续时间缩短。
不同精神分裂症患者睡眠障碍也表现出差异
同时,不同精神分裂症患者组间的睡眠障碍表现也存在显著差异,例如,住院患者在睡眠时间、活动水平和生物钟稳定性方面较门诊患者表现更加严重。这表明精神分裂症临床症状的严重程度不仅取决于病理机制,还受到患者生活环境和治疗状态等多种因素的影响。
2
睡眠障碍对精神分裂症的影响
夜间睡眠不足会进一步导致白天功能障碍和生活质量下降。大量临床研究证实,睡眠障碍与精神分裂症患者核心症状组之间存在显著且特异的关联模式。
睡眠障碍会加重精神分裂症症状并影响治疗效果
睡眠障碍不仅加重精神分裂症的阳性症状(如幻觉和妄想),还会加重负性症状(如情感冷漠和社交退缩),进而影响患者的认知功能。此外,患有睡眠障碍的精神分裂症患者表现出治疗依从性差、静坐不能和攻击性言语。
睡眠障碍的精神分裂症患者自杀行为风险增加
在一项为期八年的纵向研究中,结合Kaplan–Meier生存分析和对数秩检验,发现患有精神分裂症谱系障碍且频繁失眠的患者,其累计自杀行为风险显著增加,证实睡眠障碍会增加这些患者的自杀风险。这些结果显示睡眠问题对精神分裂症患者产生负面影响,强调了加强临床关注精神分裂症患者睡眠的必要性。
3
现有治疗的局限性
目前,治疗合并睡眠障碍(SD)的精神分裂症(SCZ)患者主要依赖传统抗精神病药物,特别是非典型抗精神病药物如氯氮平、奥氮平、喹硫平、利培酮、齐拉西酮和帕利立酮。
这些药物有助于提高精神分裂症患者的总睡眠时间和睡眠效率。然而,它们在改善SD方面的疗效有限,并伴有如体重增加和代谢综合征等副作用,部分患者在接受治疗后仍出现睡眠质量差和睡眠结构障碍。
药物疗法存在副作用,生物行为疗法逐渐兴起
尽管药物治疗是主要干预手段,但患者对药物依赖和副作用的担忧使得部分人不愿长期使用。此外,认知行为疗法(CBT)在改善持续妄想或幻觉患者的睡眠上已显示一定效果,但其临床应用仍需更多研究支持。这表明当前治疗精神分裂症合并睡眠障碍的方法面临显著局限,需探索新的治疗靶点和方法。
近年来,褪黑素的研究显示其在调节睡眠-觉醒节律、改善睡眠质量和缓解精神分裂症症状方面有效。同时,生物反馈疗法及其他非药物干预(如光疗和睡眠卫生教育)也被认为是有效的辅助治疗方式,可以改善患者睡眠质量。
总体而言,现有精神分裂症与睡眠障碍的联合治疗方法在疗效和耐受性上存在明显局限,迫切需要通过进一步研究和临床试验来探索新的治疗方案,以为患者提供更有效和安全的治疗选择。
近年来,研究发现肠道微生物群在调节精神健康和睡眠质量方面扮演着重要角色。数据显示,精神分裂症患者的肠道微生物群与健康个体存在显著差异,表现为多样性降低和特定微生物群落的失衡。与此同时,睡眠障碍的发生也与肠道微生物群的组成变化密切相关,这种相互作用可能通过微生物群-肠道-大脑轴影响患者的情绪、认知功能和睡眠模式。
1
精神分裂症患者的肠道菌群失调
多项研究发现,肠道微生物群与精神分裂症(SCZ)的病理机制、症状和认知功能密切相关。
关于精神分裂症患者肠道微生物群变化的研究
编辑
编辑
变形菌增加,拟杆菌减少,这会影响代谢健康
高通量测序结果表明,与健康对照组相比,精神分裂症患者的肠道微生物群丰富度指数(Chao)和多样性指数(Shannon)均较低,且特定微生物群的组成和丰度存在显著差异。曼-惠特尼U检验显示,精神分裂症患者的拟杆菌门(Bacteroidetes)显著减少,而变形菌显著增加。
Faecalibacterium等产生丁酸盐的抗炎菌减少
在属层面,与抗炎和神经保护作用相关的丁酸产生菌Faecalibacterium、Coprococcus和Bacteroides的丰度下降,而Prevotella和Collinsella丰度增加。短链脂肪酸(SCFAs)产生菌减少和促炎细菌增加可能与精神分裂症(SCZ)患者的病情严重程度有关。
微生物群的差异可能有助于判断疾病不同阶段
在不同疾病阶段的研究中,急性精神分裂症患者的嗜血杆菌属(Haemophilus)和Faecalibacterium数量减少,而缓解期患者则表现出巨单胞菌属(Megamonas)和Megasphaera的增加。
注:在这里小编推测嗜血杆菌属(Haemophilus)在肠道里变少可能更像“口源菌/黏膜相关菌”的信号,急性期患者的黏膜生态位与免疫/炎症微环境发生了系统性改变,而不是单纯少了一种好菌或坏菌,也可能是研究存在饮食,治疗等的混杂因素。
此外,口腔常驻细菌如Veillonella atypica、唾液链球菌(Streptococcus salivarius)和Bifidobacterium dentium在精神分裂症患者肠道微生物群中显著富集,这种病理状态可能削弱肠道屏障和免疫功能,为口腔细菌的肠道定殖提供有利条件。
一项涵盖细菌、真菌、古菌和病毒的综合多界微生物组分析发现,精神分裂症患者肠道中链球菌、脱硫弧菌(利用氨基酸或脂肪酸作为碳源)和Methanobrevibacter smithii等微生物数量增加,而丁酸盐产生菌的数量减少。
精神分裂症人群中吸烟率、牙周状况差异常见;如果口腔来源菌输入或口腔生态改变,使得口腔菌更容易通过吞咽等进入肠道并在那里存活、甚至黏附定殖肠道菌群组成更像“口腔来源的组合”,常见于炎症、屏障受损或环境改变的肠道。可以推测病源菌增加 + 产丁酸菌(如 Faecalibacterium)下降+ 屏障/炎症指标异常(如 LPS/DAO/zonulin、粪便钙卫蛋白、炎症因子),可能是未来辅助判别急性精神分裂症的重要标志组合。
2
与睡眠障碍相关的微生物群变化
在具有睡眠障碍(SD)的人类和动物模型中也观察到了微生物群变化。
关于睡眠障碍患者肠道微生物群变化的研究
编辑
微生物群的多样性与睡眠效率和睡眠时间正相关
对健康成年人睡眠的研究发现,微生物群的多样性与睡眠效率和总睡眠时间呈正相关,而与睡眠碎片化呈负相关。简而言之,肠道微生物群的多样性促进了更健康的睡眠。
此外,针对学龄前儿童的研究表明,夜间睡眠时间较长的孩子肠道微生物群结构与睡眠时间较短的孩子不同,且微生物群中的双歧杆菌和拟杆菌与睡眠时间延长、睡眠效率提高和夜间清醒时间缩短相关。
睡眠紊乱导致肠道菌群失调,普雷沃氏菌等增加
研究还表明,睡眠和昼夜节律紊乱会导致人类和动物模型中的肠道微生物群失调,表现为致病菌增加和有益菌减少。在人类中,睡眠不足和质量差常与普雷沃氏菌科(Prevotellaceae)和丹毒丝菌科(Erysipelotrichaceae)的增加及瘤胃球菌属(Ruminococcus)的减少有关。
在动物模型中,睡眠剥夺和破碎会导致毛螺菌科(Lachnospiraceae)和丹毒丝菌科(Erysipelotrichaceae)增加,而乳杆菌科(Lactobacillaceae)和双歧杆菌科(Bifidobacteriaceae)减少。
人类和小鼠的平行实验显示,不同物种的肠道微生物群存在昼夜节律振荡。此外,将人类在经历时差反应前后的肠道微生物群移植到无菌小鼠中,会导致微生物群结构改变,并出现代谢异常,如体重增加、血糖升高和体脂肪堆积。
不同类型的睡眠障碍患者显示出微生物群的特征差异
不同类型的睡眠障碍(SD)患者似乎表现出不同的肠道微生物群特征。慢性失眠患者主要表现为放线菌门数量增加,抗炎菌属Faecalibacterium含量减少,这一变化可能与长期低度炎症状态相关。
相比之下,急性失眠患者的肠道微生物群中,厚壁菌门增加而拟杆菌门减少,这一模式与急性24小时睡眠剥夺实验的发现高度相似,表明这种短期微生物失衡可能是对睡眠模式突然变化的压力反应。
此外,厚壁菌门数量增加,而拟杆菌门减少,导致(F/B)比例升高。关于急性睡眠时间表延迟的研究进一步证实,睡眠与觉醒周期的改变会增加F/B比。研究还报告指出,急性昼夜节律紊乱促进疾病相关的嘌呤代谢和丁酸生成乙酰辅酶A发酵途径,这些通路与宿主能量代谢和炎症反应相关。
注:这些代谢变化在恢复期内往往会逆转。
3
精神分裂症与睡眠障碍的微生物共性
基于研究发现,精神分裂症(SCZ)患者和睡眠障碍(SD)患者的肠道微生物多样性均降低,且特定微生物群组成发生类似变化。这种多样性的缺乏可能削弱肠道微生态系统的稳定性,增加疾病风险。
能代谢膳食纤维的菌种减少,而促炎菌增加
在微生物群组成方面,抗炎菌减少、促炎菌增加,这与疾病易感性密切相关。此外,结肠中的微生物如拟核菌、双歧杆菌和Faecalibacterium能发酵不易消化的碳水化合物和寡糖,从而合成短链脂肪酸(SCFAs)。这些属的减少显著影响丁酸盐、丙酸酯和乙酸盐的生产。
微生物群通过肠脑轴影响精神分裂症和睡眠障碍
与睡眠密切相关的特定微生物群在精神分裂症患者中数量减少。例如,双歧杆菌的减少与睡眠效率降低和睡眠碎片化有关,而Prevotella的增加可能干扰睡眠结构。
在关于睡眠与肠道微生物组组成的研究中,曼-惠特尼U测试显示,Ellagibacter isourolithinifaciens和Senegalimassilia faecalis在睡眠质量良好的患者中显著富集。在属层面,Senegalimassilia与患者的较好睡眠质量呈正相关。
这些微生物改变的特征表明,精神分裂症和睡眠障碍通过微生物群-肠道-大脑轴(MGBA)相互作用。这一过程涉及短链脂肪酸生成减少、全身炎症加剧和昼夜节律紊乱等机制,这些因素共同促进并加重两种疾病的共存和进展。
在肠道微生物群影响精神疾病的复杂病理网络中,宿主的遗传背景和环境因素共同构成了疾病易感性的基础。全基因组多效性分析揭示了胃肠疾病与精神疾病之间的广泛遗传相关性、共同的致病基因和通路,以及它们与肠道微生物群的遗传关联,证实了微生物群-肠道-大脑轴(MGBA)在这两类疾病的共同遗传基础中的关键作用。
多种环境因素,如地理位置、饮食和生活方式,动态调节肠道微生物群的丰富性和多样性,同时影响MGBA的功能稳态。MGBA作为宿主与微生物相互作用的调控枢纽,通过神经、内分泌和免疫系统之间的多层次双向通信网络,整合肠道微生态系统与中枢神经系统的功能耦合。
具体而言,肠道微生物群通过迷走神经直接调节中枢神经系统的活动,影响血脑屏障的通透性、神经炎症及通过细胞因子介导的免疫调节通路促进神经递质的合成和代谢。MGBA功能障碍可能导致病理变化,如下丘脑-垂体-肾上腺(HPA)轴异常激活及小胶质细胞持续激活,这些与精神疾病如精神分裂症(SCZ)和睡眠障碍(SD)的共病机制密切相关。
肠道微生物群及其代谢物通过肠-脑轴相互作用
编辑
1
神经递质调控
肠道菌群通过神经递质影响精神状态和睡眠
在精神分裂症(SCZ)和睡眠障碍(SD)研究中,神经递质的异常代谢被认为是影响症状的重要因素,而肠道微生物群及其代谢物在神经递质的产生和功能调控中发挥关键作用。如血清素(5-HT)、GABA和多巴胺(DA)的代谢异常可能妨碍SCZ的神经传导及SD的睡眠调节。
研究显示,肠道微生物群通过调控色氨酸代谢直接影响5-HT的生物合成。在松果体中,5-HT经历一系列酶转换,通过N-乙酰化转化为N-乙酰丝氨酸,并进一步O-甲基化合成褪黑素。该生物合成途径的完整性直接影响内源性褪黑素的生成,进而影响睡眠的开始和维持。
因此,肠道微生物群可以通过影响色氨酸的可用性来调控该代谢途径,从而通过特定微生物群增加色氨酸转化为5-HT而非犬尿氨酸途径,从而提高5-HT和褪黑素的水平。
乳杆菌和双歧杆菌通过GABA影响精神分裂症
研究表明,特定微生物群如乳杆菌和双歧杆菌不仅能促进色氨酸转化为血清素(5-HT),还可以直接合成GABA。作为中枢神经系统的主要抑制性神经递质,GABA与精神分裂症(SCZ)患者的焦虑和抑郁症状密切相关,其代谢异常对症状产生影响。通过与GABA受体结合,它有效降低神经兴奋性,延长慢波睡眠时间。
总之,神经递质的代谢在精神分裂症和睡眠障碍的病理机制中扮演重要角色。肠道微生物群通过影响神经递质的水平,不仅影响精神症状,还通过改变睡眠加重病情。因此,针对神经递质代谢的干预措施为治疗SCZ合并SD患者提供了新的思路与策略。
2
免疫炎症通路
肠道微生物群组成的变化会促进神经炎症,而这种炎症被认为是多种精神疾病(包括精神分裂症(SCZ)和睡眠障碍(SD))的重要病理机制。
IL-6和TNF-α等促炎细胞因子增加易导致精神分裂症恶化
肠道微生物群及其代谢物是调控小胶质细胞的成熟、形态和功能的关键分子。微生物群的变化可激活胶质细胞,促使IL-6和TNF-α等促炎细胞因子的释放,这影响神经元的存活与功能,破坏血-脑屏障的完整性,使外周炎症细胞因子和神经活性代谢物渗透中枢神经系统,最终诱发神经炎症反应。
针对SCZ患者的纵向研究显示,IL-6和TNF-α水平较高与脑源性神经营养因子(BDNF)水平下降及认知障碍相关。这些炎症标志物水平较高的SCZ患者更可能出现症状恶化。免疫系统在特定症状中也发挥作用,炎症刺激能改变健康个体腹侧纹状体区域的神经活动,导致动机减弱和奖赏处理缺陷,表明炎症与负性和认知症状之间的病理生理关系。
急性炎症升高也影响睡眠时间和深度
此外,睡眠障碍(SD)与炎症标志物如C反应蛋白(CRP)和IL-6的增加有关,急性炎症升高可改变睡眠的时间和深度。使用曼-惠特尼大学测试的统计分析发现,与对照组相比,接受72小时快速眼动(REM)睡眠剥夺的小鼠血浆内脂多糖浓度升高和TNF-α显著上调。
限制大鼠睡眠后,血-脑屏障的选择性过滤功能减弱,降低了阻断有害物质的能力,增加了神经炎症的风险。
通过调节肠道菌群的抗炎治疗有助于改善精神分裂症合并睡眠障碍
在临床干预中指出,使用认知行为疗法(CBT)治疗慢性和原发性失眠的老年患者后,炎症风险标志物减少,显示免疫炎症通路与睡眠障碍存在显著关联。
抗炎治疗可能是改善SCZ合并SD患者的重要策略。通过降低炎症通路则可减少小胶质细胞的激活和促炎细胞因子的产生,从而改善患者的整体症状和生活质量。
未来的研究应集中于探索特定的抗炎干预方法(如细胞因子拮抗剂或微生物组调控)对精神分裂症(SCZ)合并睡眠障碍(SD)患者的治疗效果及潜在神经保护机制。
3
神经内分泌调节
下丘脑-垂体-肾上腺轴(HPA)是神经内分泌系统的核心部分,在调节应激反应中发挥关键作用,其功能障碍已成为肠道微生物群介导睡眠障碍(SD)的重要机制之一。
压力下的高糖皮质激素导致认知障碍和焦虑增加
在压力条件下,下丘脑分泌的促肾上腺皮质激素释放因子(CRF)激活垂体前叶,释放促肾上腺皮质激素(ACTH),随后刺激肾上腺皮层合成糖皮质激素,如皮质醇。
长期过度暴露于糖皮质激素会影响大脑不同区域的神经元可塑性。这些结构重塑与认知功能障碍和动物实验中焦虑类行为增加密切相关,表明糖皮质激素介导的异常神经回路可能是慢性压力和精神障碍中认知缺陷和情感症状的重要病理基础。大量临床证据表明,HPA轴功能障碍在精神分裂症中普遍存在。
具体而言,精神分裂症患者的皮质醇觉醒反应(CAR)呈现明显异常模式,包括过度多动或钝化。这些患者的晨间基础皮质醇水平显著高于健康人群,而这种HPA轴功能障碍的模式具有疾病特异性,在高风险精神病人群中未观察到类似变化。
精神分裂症患者松果体受损影响褪黑素合成分泌
神经影像学研究显示精神分裂症(SCZ)患者的松果体体积减少、钙化加重,导致褪黑素合成和分泌功能受损。这种多层次内分泌障碍与SCZ临床症状的严重程度相关,并扰乱睡眠与觉醒周期,增加睡眠起始潜伏期和睡眠连续性受损情况。
可见,神经内分泌异常与睡眠障碍(SD)在SCZ中形成复杂的双向调控网络。睡眠不足激活下丘脑CRF神经元,导致HPA轴过度激活和皮质醇增加,而高皮质醇水平则抑制褪黑素合成,干扰睡眠调节,进一步加重SD。这一过程形成“内分泌障碍-睡眠问题-疾病恶化”的病理闭环,促进SCZ的进展和症状维持。
4
代谢物介导的调控
肠道微生物群生成的代谢物,如短链脂肪酸(SCFA)、色氨酸衍生物和胆汁酸,直接或间接参与中枢神经系统的调控,影响神经递质的平衡、维持免疫稳态,并调节昼夜节律。
丁酸盐有助于改善精神分裂症状和睡眠质量
菌群失调可破坏肠道屏障的完整性,增加肠道通透性,损害血-脑屏障的功能。这些双重屏障的破坏使得细菌代谢产物和促炎细胞因子等有害物质更易进入中枢神经系统,引发神经炎症反应。
丁酸盐作为关键代谢物,不仅对维持肠道黏膜的完整性至关重要,还能穿越血-脑屏障,通过激活迷走神经调节睡眠和清醒周期。动物研究发现,静脉注射丁酸盐显著延长了大鼠的非快速眼动睡眠(NREMS)时间。
此外,血清丁酸盐水平的升高与精神分裂症(SCZ)患者阳性和阴性综合征量表(PANSS)的阳性症状分数下降相关,暗示其潜在的神经保护作用。
血清素和胆汁酸水平下降可能是影响精神分裂症和睡眠障碍的重要机制
精神分裂症(SCZ)患者还表现出显著的色氨酸代谢障碍,研究发现其血浆色氨酸水平通常较低,这可能由于色氨酸向犬尿氨酸的转化增强和5-羟色胺(5-HT)合成途径的减少。这种转变导致神经递质5-HT和褪黑素的合成减少,由于这些物质在睡眠调节中起关键作用,其缺乏会加重精神分裂症患者的睡眠障碍(SD)。
代谢物分析显示,精神分裂症患者胆酸通路中的代谢物如糖果酸、牛磺酸脱氧胆酸和牛磺鹅脱氧胆酸的浓度显著降低,胆汁酸水平的下降通过影响抗炎信号通路,促进慢性低度炎症微环境的形成,从而增加神经退行性疾病和精神障碍的风险。研究还发现慢性失眠与特定胆酸结构和组成之间存在关联。
因此,代谢轴的异常不仅是精神分裂症的典型生物标志物,也是精神分裂症中睡眠障碍发病的关键机制之一。针对肠道微生物群及其代谢物的干预措施可为改善精神分裂症合并睡眠障碍患者提供新的治疗靶点。
5
昼夜节律紊乱
肠道微生物群与昼夜节律密切相关,其组成会随宿主生物钟的变化而波动,反之肠道微生物群的紊乱也会影响生物钟、肠道免疫功能和营养代谢。
肠道微生物群影响人体生物钟在精神疾病中发挥重要作用
肠道微生物群的组成和代谢物对宿主时钟基因(如CLOCK和Bmal1)的表达具有独特的调控作用。研究发现,微生物代谢物可以调节中枢神经系统和肝脏的生物节律,比如,短链脂肪酸(SCFAs)直接调控肝细胞中的时钟基因表达,影响宿主生物节律的稳定性。
肠道微生物群失衡在精神分裂症(SCZ)患者中较常见,这种失衡可能通过肠道-大脑轴影响宿主时钟基因的表达。分析首次发作SCZ患者的单核血细胞发现,与健康对照相比,患者的CLOCK、PER2和CRY1基因表达显著降低。而在慢性SCZ患者的成纤维细胞中,时钟基因CRY1和PER2的节律表达丧失。这些研究揭示了肠道微生物群通过代谢物与时钟基因的相互作用,对SCZ患者昼夜节律的稳定性至关重要。
时钟基因表达的变化不仅影响肠道微生物群的结构,还进一步影响核心昼夜节律输出通路的效率。研究表明,昼夜节律紊乱与肠道-大脑轴(MGBA)之间存在显著相互作用,MGBA在精神疾病的整个过程中持续影响患者的健康状况和临床症状。
昼夜节律调节神经内分泌活动影响精神分裂患者临床症状
结合本文前面的内容,昼夜节律通过调节神经内分泌活动和自主神经系统活动,调节睡眠、新陈代谢和免疫等生理过程。该输出通路的失效可能导致睡眠结构紊乱、压力激素节律失调,以及代谢和免疫反应的周期性紊乱,这些共同构成了精神分裂症患者常见的临床症状基础。与此同时,时钟基因表达的变化反过来会影响肠道微生物群的组成,显示出丰富度和多样性的减少,形成复杂的相互作用网络。
例如,研究显示宿主昼夜节律的核心激活基因(Bmal1)和抑制基因(Per1, Per2)共同调节肠道微生物群组成的昼夜振荡。这些时钟成分的基因敲除消除了这些节律波动,改变了小鼠的微生物群落结构。
深入研究肠道微生物群与其代谢物及时钟基因之间的关系,不仅有助于理解精神分裂症的发病机制,还将为基于肠道微生物群干预的未来治疗策略提供新思路。例如,通过调整饮食或补充益生菌以改善肠道微生物群的组成,调控时钟基因的表达以优化昼夜节律信号的传递效率,改善精神分裂症患者的临床症状。
小编总结
这组证据总体在说明:睡眠/昼夜节律紊乱本身就是一个能“驱动”肠道微生态改变的上游因素,而这种改变不仅是“相关”,还可能通过菌群把代谢风险传递出去。菌群不是静态的,它会随进食时间、胆汁酸分泌、肠蠕动、激素(如皮质醇/褪黑素)等出现日内波动,群落结构不稳定和功能(例如短链脂肪酸、胆汁酸转化、内毒素负荷)也会随之波动。
如果精神分裂症患者同时存在常见的睡眠/昼夜节律紊乱,那么观察到的某些肠道菌群特征(如特定门、科/属的增减,口源菌富集,以及产丁酸菌减少等)可能并非完全由疾病本身导致,而是共同受到多条路径的影响,包括:疾病相关应激、药物与生活方式,以及睡眠—生物钟失调这一通路。
与此同时,膳食纤维代谢菌减少提示菌群功能发生改变:尤其是短链脂肪酸(SCFAs)生成能力下降,可能削弱其对肠道屏障维护与免疫调节的作用;而促炎菌增加、抗炎菌减少则意味着机体更偏向促炎状态与肠道高通透,进而可能通过“肠—免疫—脑”轴影响中枢免疫激活与神经炎症。
在监测精神分裂症菌群时,睡眠状态可能是重要的混杂因素,也可能是可干预的上游靶点(改善睡眠可能间接改善菌群与代谢/炎症表型)。
尽管睡眠障碍与多种精神疾病的发生和发展密切相关,但针对精神分裂症(SCZ)合并发病的干预研究仍然不足。目前大多数治疗策略集中于症状控制,如通过抗精神病药物改善核心精神症状,而较少关注肠道微生物群失调机制。
因此,以下讨论提出针对肠道微生物生态的新治疗策略,包括特定益生菌、益生元、饮食干预、粪便微生物移植(FMT)和靶向代谢物治疗,旨在调节肠道-脑轴功能,以改善精神分裂症患者的睡眠问题及相关精神症状。
针对精神分裂症患者的微生物群干预方法
编辑
1
个性化补充益生菌
个体间肠道微生物群的差异会影响药物的疗效和毒性,因此个性化的肠道微生物群调控有助于改善药物反应。
通过检测精神分裂症(SCZ)患者的肠道微生物群,可以制定个性化益生菌方案,有效调节肠道微生物组成,从而改善症状。
长双歧杆菌等益生菌能改善精神状态
益生菌的疗效因菌株而异,不同菌株在调节炎症和压力方面表现出不同作用机制。例如,婴儿双歧杆菌(Bifidobacterium infantis 35624)已被证明能逆转HPA轴功能障碍,并与抗炎和促炎细胞因子比例的正常化相关,具有临床意义。
鼠李糖乳杆菌(Lactobacillus rhamnosus,JB-1)能降低压力诱发的皮质酮水平,缓解焦虑和抑郁行为。长双歧杆菌(Bifidobacterium longum NCC3001)通过MGBA内的迷走神经通路传递信号,使低度肠道炎症小鼠的焦虑行为和海马BDNF水平恢复正常。该菌株在人类试验中也显示抗抑郁效果,与多个情绪处理脑区活动变化相关。
益生菌有助于改善炎症相关、情绪状态及睡眠
在一项针对失眠患者的双盲研究中,参与者接受了植物乳杆菌(Lactobacillus plantarum PS128)或安慰剂。结果显示,PS128组在疲劳水平、脑电波活动和深度睡眠期间醒来次数方面有所改善,同时也缓解了焦虑和抑郁症状。这表明特定益生菌菌株可能通过微生物群-肠道-脑轴的机制(如GABA能系统调控)影响睡眠结构和情绪状态。
一项针对双相情感障碍和精神分裂症谱系障碍患者的双盲随机安慰剂对照试验表明,补充多株益生菌配方能改善与肠道通透性和炎症相关的生物标志物,并对认知功能产生积极影响。
这些研究结果揭示了益生菌补充剂在调节神经功能方面的潜在治疗价值,为进一步探索益生菌的干预机制提供了重要理论基础。
2
膳食益生元补充
益生元是一类能够抵抗宿主消化酶的功能性膳食成分,能选择性促进有益肠道微生物群的代谢活动,从而发挥生理调节功能。
菊粉等益生元改善精神分裂症的精神和行为症状
多项研究发现,半乳糖、菊糖型果聚糖及其合成制剂能够降低促炎因子(如高敏感性CRP、IL-6和TNF-α)的水平,表明特定的益生元干预可以调控炎症相关生物标志物的表达。菊粉作为可溶性膳食纤维,已被证实改善精神分裂症(SCZ)模型小鼠的精神和行为症状,同时增加有益菌数量并改善肠道通透性。
服用低聚果糖和低聚半乳糖的复合制剂也改善了SCZ模型小鼠的肠道功能。这些变化可能通过重塑肠道微生物群、降低促炎细胞因子水平以及增强肠道通透性,从而减少病原体与肠道黏液层的接触,为改善多种精神疾病的核心症状提供重要的病理生理基础。
益生元调节神经递质平衡改善精神状态及睡眠
另一项研究表明,短链半乳糖与长链果糖的联合干预显著促进了短链脂肪酸中乙酸和丁酸的生成。这些益生元成分还有效调控下丘脑和海马体中核心时钟基因BMAL1和CLOCK的表达。
这表明益生元通过多重靶点的协同效应发挥神经调节功能。特别是,益生元对时钟基因的调节可能通过恢复昼夜节律改善睡眠问题,并通过调节神经递质平衡潜在改善精神分裂症核心症状。
为促进益生元疗法在精神疾病临床实践中的应用,后续研究应明确不同益生元的应用价值、个体化剂量策略以及治疗的安全性和有效性评估。
3
饮食调整干预
特定饮食成分通过与肠道菌群互作影响睡眠质量
饮食调整是个性化治疗的重要部分,已有大量科学证据证明饮食与睡眠之间存在关系。健康饮食有助于改善睡眠质量,而加工食品和高糖食物则与睡眠质量较差相关。
特定饮食成分(如脂肪和蛋白质)及习惯(如牛肉、咖啡和干果摄入)通过与肠道微生物群相互作用影响睡眠。
生酮和高纤维饮食有助于改善睡眠质量和免疫
近年来,生酮饮食受到广泛关注,已被证明是改善伴有代谢异常的精神疾病患者心理和代谢健康的一种可行辅助治疗方法。根据匹兹堡睡眠质量指数(PSQI)评估,接受生酮饮食干预后,双相情感障碍和精神分裂症(SCZ)患者的主观睡眠质量有所改善。这表明,生酮饮食可以纳入精神疾病患者的综合治疗策略,有望在控制精神症状、调节代谢和改善睡眠方面带来多重益处。
此外,从高纤维饮食的角度来看,短链脂肪酸(SCFAs)是通过肠道共生细菌发酵膳食纤维产生的。当膳食纤维摄入不足时,SCFAs的生成水平会降低,这将对宿主的各种生理功能产生不利影响。增加膳食纤维摄入以促进SCFA的产生,不仅能增强血液和脑屏障的保护功能,维持肠道黏膜的完整,还能调节肠道微生物群的组成,从而协同促进免疫稳态的建立。
基于这一机制,这种饮食干预策略为改善精神障碍患者的睡眠提供了新思路。通过缓解肠道炎症状态并调节神经递质水平,它可以改善患者的精神症状和睡眠质量。
4
粪菌移植
粪菌移植(FMT)是一种以重建微生物群落为核心的生物治疗策略,通过将健康供体的粪便移植到患者的胃肠道,直接重组受体的肠道微生物群,促进微生物生态系统的平衡,从而实现临床治疗目标。
粪菌移植有助于调节情绪、行为和睡眠质量
作为一种突破性干预方法,粪菌移植(FMT)在动物模型和初步临床研究中展现出独特优势。研究发现,通过移植精神分裂症(SCZ)患者的肠道微生物群,被移植小鼠出现了类似SCZ的行为,包括多动、焦虑增加、社交互动受损和记忆缺陷。
注:这些小鼠还显示出周围和中枢神经系统中色氨酸代谢犬尿氨酸-犬尿酸通路的显著激活,前额叶皮层基底细胞的多巴胺能神经递质释放增强,以及海马体5-HT水平升高,同时谷氨酸能神经递质浓度下降,谷氨酰胺和GABA水平上调。
FMT在人体临床研究中展现出重要价值。比较接受健康供体FMT前后的粪便样本显示,α多样性显著增加,柯林斯氏菌属(Collinsella)和双歧杆菌的丰度也有所上升。FMT治疗还显著降低了患者的血液皮质醇水平,同时改善了睡眠和情绪相关评估指标。
作为潜在治疗干预,FMT通过调节肠道微生物群的组成和功能,促进神经递质及其前体物质的合成,从而发挥治疗作用。这些研究表明,FMT能够有效调节情绪和行为,促进多种精神疾病患者的症状缓解。
!
注意事项
尽管粪菌移植(FMT)展现了广泛的治疗前景,但其临床应用面临标准化不足、疗效不一及长期安全性验证的挑战。高通量测序技术可帮助识别健康稳定的供体。口服FMT胶囊相比传统灌肠方法更方便,患者依从性更高。多样化的给药途径,如结肠镜输注、鼻肠管插管或口服胶囊,有助于平衡疗效与患者依从性。建立安全监测系统可预防和控制感染风险,加强安全监管。
此外,在进行FMT治疗时,必须密切关注精神障碍患者的知情同意能力,确保他们能够在充分了解潜在风险和益处的基础上做出自主决策,并通过伦理审查委员会的批准进行标准化申请。尽管仍需更多研究验证FMT在精神分裂症(SCZ)和睡眠障碍(SD)治疗中的长期效果,但作为个性化治疗的一部分,FMT显示出广泛的应用前景。
5
靶向代谢物药物治疗
在精准医疗领域,靶向代谢物药物治疗正成为新研究方向。尽管传统微生物群移植在临床应用中取得了一定成效,但个体肠道微生物群的差异严重限制了其治疗效果的稳定性和重复性。因此,直接补充微生物代谢物或前体物质为解决这一问题提供了新思路。
靶向代谢物能够避免个体菌群差异,更精准治疗
这种策略能够绕过微生物群移植中的个体差异,直接作用于人体代谢网络,实现更精确的调控。短链脂肪酸(SCFAs)和色氨酸代谢在生理和病理过程中起着关键作用。
以吲哚胺2,3-二氧加氧酶(IDO)抑制剂为例,IDO是色氨酸代谢中的限速酶,其过度活化可导致色氨酸耗竭和一系列神经毒性代谢物的产生,进而引发异常免疫反应和神经功能障碍。IDO抑制剂通过抑制其活性,可以有效调节免疫反应,减少神经毒性代谢物的产生,展现出治疗多种疾病的潜力。
SCFAs和色氨酸代谢物与精神分裂症(SCZ)发病机制及睡眠调节密切相关。未来研究应进一步探讨代谢物的具体作用机制,推动其临床应用的发展,为合并睡眠障碍的SCZ患者提供更安全、更有效的治疗选择。
治疗策略总结
并非所有精神分裂症(SCZ)患者都适合微生物组靶向干预。潜在候选者的识别应基于具体特征,例如明显的胃肠道症状、抗精神病药物反应不良或显著副作用、异常睡眠结构与疾病活动的强相关性,以及肠道微生物组分析结果显示与健康对照有显著偏差。
根据临床表型和微生物特征进行对应治疗选择
目标群体应根据临床表型和微生物特征的组合进行选择。在推进微生态干预时,应明确不同策略的优先级,优先考虑低风险、非侵入性的方法。例如,补充特定益生菌和益生元可作为基础干预,而饮食模式调整可作为长期管理策略。
相比之下,高风险且难以预测的治疗如粪便微生物移植(FMT)应仅考虑用于难治症状、对传统疗法反应不足及严重肠道菌群失调的患者,并在充分知情同意和严格伦理监督下使用。
此外,实施时需认真考虑个体差异和潜在风险。益生菌和益生元干预应考虑菌株特异性效应和个体耐受性,饮食干预需关注患者的依从性和营养平衡。对于FMT,严格的供体筛查至关重要,并需密切监测感染、免疫和代谢不良反应,以及长期精神病学结局,同时建立动态的疗效-安全性评估体系。
本文分析了精神分裂症(SCZ)及其相关睡眠障碍患者的典型肠道微生物群特征。研究发现,这两种状况均表现出肠道微生物群α多样性下降、短链脂肪酸产生细菌减少以及促炎微生物群比例增加。由此可以推测,SCZ患者的肠道菌群失调可能是导致睡眠质量下降的重要因素,而睡眠质量恶化又通过反馈机制加重精神症状,形成恶性循环。
肠道菌群失调、中枢神经系统功能障碍和睡眠稳态紊乱在宿主体内形成自我延续的动态循环,导致精神病理表现和生理节律紊乱的周期性恶化。这些发现为理解精神分裂症患者共病睡眠障碍的机制提供了重要的理论框架。针对肠道微生物群以打破这一恶性循环,为这类患者提供了有前景的新颖治疗视角。
现有研究表明,治疗应采取综合策略。虽然非典型抗精神病药物能改善精神病症状,但对睡眠结构的复杂影响仍需深入评估。认知行为疗法同样对失眠有效,但需根据精神病症状进行调整。因此,针对肠道微生物群的干预策略正在向多层次和精准化方向发展,具体包括补充特定益生菌和益生元,结合饮食调整,通过多途径共同改善精神症状和睡眠质量。尽管粪便微生物移植(FMT)作为高级干预措施展现出潜力,但其效果仍需通过标准化研究来验证。实施时应重点关注胃肠道症状、药物反应不足、精神症状严重程度与睡眠节律紊乱以及肠道微生物群特征显著偏离健康标准的患者。整合临床表型与微生物特征的模型将为精准应用干预策略提供基础,最终推动微生物群靶向治疗的系统化及个性化发展。
我们每个人都在不断追求身心健康的道路上前行,理解肠道微生物群在我们的心理与生理健康中所扮演的角色,不仅能增进我们对自身健康的认识,更能激励我们在日常生活中关注饮食、生活方式等易被忽视的细节。希望未来的研究能够持续启发我们,推动科学领域的进步,为那些受到精神障碍困扰的人们带来新的希望和解决方案。通过这样的努力,我们将共同迈向一个更加健康和明亮的未来。
注:本账号发表的内容仅是用于信息的分享,在采取任何预防、治疗措施之前,请先咨询临床医生。
主要参考文献:
Huang, Z., Huang, Z., Du, Z., Gao, X., Jiang, Y., Zhou, Z., & Zhu, H. (2026). Role and mechanism of gut microbiota and metabolites in schizophrenia complicated with sleep disorder. Gut Microbes, 18(1). https://doi.org/10.1080/19490976.2025.2607817.
McCutcheon RA, Reis Marques T, Howes OD. Schizophrenia-An Overview. JAMA Psychiatry. 2020 Feb 1;77(2):201-210.
Meyer N, Faulkner SM, McCutcheon RA, Pillinger T, Dijk DJ, MacCabe JH. Sleep and Circadian Rhythm Disturbance in Remitted Schizophrenia and Bipolar Disorder: A Systematic Review and Meta-analysis. Schizophr Bull. 2020 Sep 21;46(5):1126-1143.
Reeve S, Sheaves B, Freeman D. Sleep Disorders in Early Psychosis: Incidence, Severity, and Association With Clinical Symptoms. Schizophr Bull. 2019 Mar 7;45(2):287-295.
McGuinness AJ, Davis JA, Dawson SL, Loughman A, Collier F, O’Hely M, Simpson CA, Green J, Marx W, Hair C, Guest G, Mohebbi M, Berk M, Stupart D, Watters D, Jacka FN. A systematic review of gut microbiota composition in observational studies of major depressive disorder, bipolar disorder and schizophrenia. Mol Psychiatry. 2022 Apr;27(4):1920-1935.
Mayeli A, LaGoy AD, Smagula SF, Wilson JD, Zarbo C, Rocchetti M, Starace F, Zamparini M, Casiraghi L, Calza S, Rota M, D’Agostino A, de Girolamo G; DiAPAson Consortium; Ferrarelli F. Shared and distinct abnormalities in sleep-wake patterns and their relationship with the negative symptoms of Schizophrenia Spectrum Disorder patients. Mol Psychiatry. 2023 May;28(5):2049-2057.
Penninx BWJH, Lange SMM. Metabolic syndrome in psychiatric patients: overview, mechanisms, and implications. Dialogues Clin Neurosci. 2018 Mar;20(1):63-73.
Zhu C, Zheng M, Ali U, Xia Q, Wang Z, Chenlong, Yao L, Chen Y, Yan J, Wang K, Chen J, Zhang X. Association Between Abundance of Haemophilus in the Gut Microbiota and Negative Symptoms of Schizophrenia. Front Psychiatry. 2021 Jul 30;12:685910

谷禾健康
植物性食物 Plant-based Diet
食物,尤其是植物性饮食,具有复杂的化学多样性。然而,肠道细菌大规模代谢植物营养素的活动在很大程度上仍是不完全清楚的。
近日,来自德国耶拿莱布尼茨天然产物研究与感染生物学研究所(Leibniz-HKI)微生物组的 Gianni Panagiotou团队联合上海交通大学医学院于发表在《Nature microbiology》整合并系统分析了包含酶反应和食物健康益处信息的多个数据库,以及3,068个全球公开的人类微生物组数据和来自可食用植物的775种植物营养素,发现植物营养素的转化与多种肠道微生物编码的酶相关。
通过结构预测与体外实验证实关键菌株(如 Eubacterium ramulus)对黄酮类化合物的特异代谢作用及其关键酶的催化功能。植物营养素的转化能力存在显著个体差异和地域特异性,五大洲人群共享630种核心转化通路,且疾病,年龄,肥胖等都会改变我们的代谢能力。
最终,这项研究告诉我们:我们吃下的植物性食物中含有数千种化学物质,其中许多需要经过你肠道中数万亿微生物的“加工处理”才能被人体有效利用。
同时该研究创新性地建立了“膳食化学—微生物酶—宿主健康”多层耦合框架,暗示在未来,营养师和医生可能会常规分析你的肠道微生物组,以推荐最适合你个人“肠道化工厂”的饮食方案,让我们真正迈向个性化营养的新时代。
“饮食决定健康”这一简单概念正被更复杂的理解所取代:我们的健康不仅取决于吃什么,还取决于我们的肠道微生物如何处理我们所吃的食物。”
就如刚发表在《Nature Microbiology》上的这项开创性研究,系统地探索了肠道微生物如何代谢植物性食物中的化学物质(称为植物营养素),以及这一过程如何影响我们的健康。来自德国、丹麦、中国等国家的科学家团队整合了全球3,068个人类肠道微生物组数据和1,300多种植物营养素信息,揭示了一个肠道惊人的”代谢宇宙”:我们肠道中约70%的微生物酶参与植物营养素的生物转化。
这意味着,当你咬下一口新鲜的草莓或清脆的西兰花时,你肠道中的微生物正在加班加点地将这些复杂化合物转化为对你身体有益的物质——但前提是拥有理想的微生物团队。
黄酮类化合物是植物性食物重要的次级代谢产物,隶属多酚类物质,广泛存在于多种膳食植物来源中,如水果、蔬菜、草药、种子、谷物及饮料等。其基本骨架通常由芳香环 A 与杂环 C 相连,并与另一芳香环相结合形成特征性结构。依据化学结构差异,黄酮类可进一步分为黄酮、黄酮醇、黄烷酮、黄烷醇、异黄酮和花青素等主要类型。
摄入后,部分黄酮类化合物可在小肠中经上皮酶 O-脱糖基化后被吸收;进入肠上皮细胞与肝脏后,吸收的黄酮类苷元进一步发生 I 相与 II 相代谢转化。然而,多数膳食黄酮类化合物(及其 I/II 相代谢产物)会到达大肠,并在肠道菌群作用下被进一步改造。大肠细菌酶可介导多种结构修饰反应,如脱羧、开环、还原、异构化和去甲基化等。
由于膳食黄酮多以糖苷形式存在,肠道细菌的 α-L-鼠李糖苷酶 可去除末端 α-L-鼠李糖,将其转化为葡萄糖苷;随后 β-葡萄糖苷酶进一步脱糖生成可吸收的苷元,苷元既可在大肠被吸收,也可继续代谢为多种开环产物。
肠道菌群介导的黄酮类代谢中,雌马酚(equol)生物合成途径是研究最为深入的代表之一,其核心在于一系列关键酶将异黄酮苷元逐步转化为雌马酚。
另一条较为清晰的途径是黄酮/黄酮醇的降解代谢,其中 Flavonifractor plautii 与 Eubacterium ramulus被认为是参与转化的活跃菌种;在这些菌种中已鉴定出多种相关酶,包括黄酮/黄酮醇还原酶(Flr)、查尔酮异构酶(Chi)、黄烷酮/黄烷醇裂解还原酶(Fcr)以及根皮苷水解酶(Phy)。
黄酮类化合物及其微生物代谢产物因具备抗炎、抗氧化与抗癌等生物活性而与多种健康获益相关,其保护作用已在结直肠癌(CRC)和炎症性肠病(IBD)等疾病研究中得到支持。
植物营养素与微生物酶的完美匹配
当想研究这个复杂关系时,首先面临的挑战是如何将植物性食物中的化学物质与肠道微生物产生的酶联系起来。研究团队人员从NutriChem 2.0数据库中获取了7,825种与可食用植物相关的低分子量植物营养素,并功将超过1,500种化合物与具有独特酶库编号的酶联系起来。经过精细筛选,最终确定了1,388种植物营养素与4,678种酶相关联。
通过分析全球3,068个健康人的肠道微生物组数据,研究团队发现这些植物营养素中有775种可被肠道微生物酶转化,涉及1,118种可食用植物。令人惊讶的是,约64%(1,226种)的这些酶仅存在于肠道微生物物种中,而在人类基因组中不存在。这意味着,如果没有肠道微生物的帮助,我们的身体将无法代谢这些植物性化合物。
广泛的代谢能力
约67%的肠道微生物酶可能参与植物营养素的生物转化
独特的微生物贡献
约64%的植物营养素转化酶仅存在于肠道微生物中,人类基因组中不存在
主要参与者
拟杆菌门和变形菌门,是最常见的植物营养素转化菌门
核心化学类别
萜类、黄酮类和生物碱是主要被转化的植物营养素类别
主要酶类别
氧化还原酶、转移酶和水解酶是参与转化的主要酶类
值得注意的是,达人群中肠道细菌物种的丰度和流行度与其基因组转化植物营养素的能力呈正相关。这意味着那些在我们肠道中更常见、数量更多的细菌,往往具有更强的代谢植物性化合物的能力。
研究团队特别关注了21种被认为是健康饮食选择的常见食物,发现它们中超过一半的已识别植物营养素与肠道微生物酶相关联。例如,黄酮类化合物根皮素的糖基化(由EC 2.4.1.4催化)和根皮素向3-(4-羟苯基)丙酸的转化(由查尔酮异构酶EC 5.5.1.6催化)都是已知的与健康相关的生物转化过程。
注:Parsnips 是英文里对 “欧洲防风草 的称呼,一种根茎类蔬菜,外形有点像白色胡萝卜。
这些发现表明,我们日常食用的许多健康食物,如蓝莓、绿茶、坚果等,其健康益处可能在很大程度上依赖于我们肠道微生物的代谢能力。换句话说,如果你肠道中缺乏特定的微生物或其产生的酶,即使食用”超级食物”也可能无法获得其全部健康益处。
随着益生菌产品的普及,许多人可能认为服用益生菌补充剂可以获得与健康肠道微生物相同的益处。但这项研究揭示了一个重要事实:常见益生菌在代谢植物营养素方面的能力远不如我们肠道中自然存在的某些细菌。
益生菌的短板 与 肠道细菌的特长
研究人员注释了59种益生菌的基因组,发现这些菌株中的酶可以潜在地转化775种与肠道微生物酶相关的植物营养素中的525种。然而,在186种与次级代谢相关途径的植物营养素中,只有116种可被益生菌和肠道微生物酶共同修饰,而70种只能被肠道微生物酶修饰。
更重要的是,能够修饰这些植物营养素的肠道细菌种类平均为158种,而能够做到这一点的益生菌种类平均仅为8种。这意味着,许多植物性食物中的生物活性化合物需要我们肠道中特定的”本土”细菌才能被有效代谢,而常见的益生菌补充剂可能无法提供这种能力。
研究还发现,共享和肠道限制性植物营养素在结构特征上存在差异。含有羧酸和羰基的植物营养素(如反式肉桂酸和柚皮素)特别富集在共享化学空间中,而肠道限制性植物营养素则富含苯环和双环基团。
注:
原文中 shared phytonutrients(共享):既能被肠道常驻菌也能被常见益生菌,预测/注释到可代谢的植物营养素。
gut‑restricted phytonutrients(肠道限制性):只被肠道常驻菌预测/注释到可代谢,而未在益生菌物种中预测到可代谢的植物营养素。
所以,肠道限制性在这里就是一个相对概念:相对于他们纳入分析的那批益生菌而言,这些化合物的代谢能力更“限制在肠道本土菌里”。
体外实验验证:Eubacterium ramulus的明星表现
为了验证这些发现,研究团队在实验室中测试了六种具有高生物转化潜力的肠道菌株对36种植物营养素的代谢能力。结果令人瞩目:Eubacterium ramulus(一种常见的肠道细菌)表现出最强的生物转化活性,能显著代谢12种底物中的11种。
黄色的代表Eubacterium ramulus 菌种
进一步研究发现,Eubacterium ramulus含有一种名为EC 5.5.1.6的酶,能够催化两种黄酮类化合物——butein和isoliquiritigenin的生物转化。当存在活的E. ramulus时,这些化合物的转化速度显著加快,并且只在活菌存在的情况下才能检测到下游产物。这一发现不仅验证了计算预测,还为理解特定细菌如何影响植物营养素的代谢提供了机制性见解。
研究结果对益生菌行业提出了挑战。许多商业益生菌可能无法提供我们肠道中自然存在的某些细菌所具有的代谢能力。同时也意味着保护我们原生肠道微生物组多样性的重要性。
如果你曾经疑惑为什么某些食物对你的朋友或家人有效却对你似乎没什么作用,这项研究可能给了答案。研究发现,植物营养素的生物转化存在显著的个体间和地理变异性,这解释了为什么相同的饮食对不同人会产生不同的健康效果。
个体差异:与α多样性(群落丰富度/均匀度)的关系
分析显示,在个体微生物组中注释的酶中,平均70%与植物营养素生物转化相关,而在次级代谢相关途径中这一比例高达90%。然而,植物营养素相关酶的比例与微生物组α多样性呈负相关,这表明植物营养素生物转化是肠道微生物群的普遍特性。但对于参与次级代谢的酶来说,这种相关性是正的,表明肠道微生物转化次级代谢相关植物营养素的能力存在差异。
个体差异很大,但洲际总体很相似
个体间植物营养素转化范围差异明显:每人可被转化的植物营养素数量在264–620之间(覆盖5个地理区域)。
植物营养素相关的微生物组独特性在不同个体之间显著高于同一个体内部。
但把人按“洲/地区”汇总后,整体“植物营养素与食物生物转化相关的酶学机器”在各地区高度相似:5个地区共同关联到的植物营养素有630种;
只有少数显示强地域特异性(如亚洲10种、大洋洲2种)。
例如黄烷醇类(taxifolin)及其相关酶 EC 1.1.1.219 只在亚洲个体中被注释到,可能与当地食物/药用植物相关。
地理差异:祖籍可能影响你的代谢能力
地理差异的有趣发现所有地区的肠道微生物组都能代谢630种常见植物营养素;然而,也有一些植物营养素表现出高度的地理特异性。
例如,黄酮类化合物taxifolin(存在于荔枝和柘树等食物中,后者是东亚的传统药物)及其相关酶EC 1.1.1.219仅在亚洲个体中被注释到。这种地理特异性可能反映了长期饮食传统与肠道微生物组之间的共同进化。
某些植物营养素的代谢能力具有地理特异性;且不同洲的酶谱仍显著不同,反映饮食差异。非洲和亚洲与其他地区的差异尤为显著,这反映了区域饮食的巨大差异。酶的变化与特定化学类别(包括黄酮类和脂肪酸相关化合物)显著相关。
为了探索饮食如何塑造这些模式,研究人员使用了已有的连续饮食信息和配对的宏基因组数据。Procrustes分析显示,可食用植物饮食记录与这些可食用植物1,665种植物营养素相关的酶丰度之间存在显著一致性,表明食用植物的摄入与微生物组的植物营养素生物转化潜力相关。
更有趣的是,对美国出生的个体和居住在泰国、刚移民到美国或已在美国生活至少20年的泰国人的数据集进行的相似性分析发现,尽管饮食习惯的适应相对温和,但泰国参与者的食用植物相关酶组成随着时间推移变得与美国出生的参与者显著相似。这表明,我们的肠道微生物组及其代谢能力可以通过改变饮食来调节,尽管这可能需要相当长的时间。
研究还发现,植物营养素相关酶的比例与年龄呈负相关,而植物营养素相关微生物组的独特性与年龄呈正相关。这表明,随着年龄增长,我们的肠道微生物组可能失去一些代谢植物营养素的能力,同时变得更加个性化。此外,511种酶与BMI显著相关,随着BMI增加,植物营养素相关酶的数量减少,这可能部分解释了为什么肥胖人群对某些植物性食物的反应不同。
研发发现
疾病状态会显著改变肠道微生物群对“具有健康益处的植物食物/其植物营养素”的生物转化潜力;这些与植物营养素相关的微生物酶特征还能用于区分健康人与患者。
研究设计
对象/数据:公共肠道宏基因组病例-对照队列,覆盖三类疾病:
方法主线
把肠道细菌酶(EC/酶学分类,且做了species-stratified分层归因到物种)与其可作用的具有健康益处的植物营养素空间关联,评估不同疾病中这些酶的丰度变化,并进一步用机器学习做判别。
主要发现1:与植物营养素相关的酶在疾病中大量失衡
IBD:炎症性肠病(Inflammatory Bowel Disease)
CRC:结直肠癌(Colorectal Cancer)
NAFLD:非酒精性脂肪性肝病(Non-Alcoholic Fatty Liver Disease)
识别到的植物营养素相关酶数量:
IBD:608
CRC:1,038
NAFLD:517
其中在患者 vs 对照中显著差异丰度
IBD:59.7%(363/608)
CRC:49.9%(518/1,038)
NAFLD:22.2%(115/517)
说明IBD 与 CRC 中,这类能处理健康植物营养素的微生物酶谱改变更广泛;NAFLD 相对较少但仍存在系统性改变。
主要发现 2:对具体健康食物的生物转化潜力改变程度不一
按每种有益食物对应的植物营养素相关 EC来看,平均有显著改变的比例:
IBD:63.0%
CRC:33.6%
NAFLD:25.3%
并且存在显著异质性:不同疾病之间差异很大;
同一疾病内部,不同食物差异也很大;
有些食物对应的酶中超过 50%都显著改变。
意味着不是“整体代谢能力统一下降/上升”,而是呈现“食物/营养素通路特异”的重塑。
主要发现 3:饮食相关酶特征可用于疾病判别,且优于非饮食相关酶
随机森林判别模型使用的特征数与内部表现:
IBD:18 个酶,auROC = 0.892
CRC:28 个酶,auROC = 0.763
NAFLD:22 个酶,auROC = 0.95
外部队列验证:三病均保持较高准确度(auROC 约 0.72–0.79)。
对照实验:若改用非饮食相关 EC(不与植物营养素关联)做 species-stratified 特征,验证集 auROC 降低:
IBD:0.779
CRC:0.698
NAFLD:0.584
这些可以推测与植物营养素生物转化直接相关的酶信号更贴近疾病相关的微生物功能改变,具有更强的可迁移判别力(尤其 NAFLD 的差距最大)。
总之,这些说明疾病状态不仅改变菌群有哪些菌,也改变其处理健康植物食物成分的功能酶库;这种功能改变具有食物/营养素通路特异性。
动物实验验证:草莓的抗炎作用为何依赖肠道微生物?
编辑
为了在体内测试参与植物营养素生物转化的肠道微生物酶是否对健康饮食的保护作用至关重要,研究人员在特定无病原体(SPF)和无菌(GF)条件下的结肠炎小鼠模型中研究了抗炎食物草莓的作用。
草莓的保护作用依赖肠道微生物
编辑
草莓抗炎作用的微生物机制
关键酶的作用:EC 4.1.1.11(天冬氨酸→β-丙氨酸和色氨酸→色胺)与减轻炎症相关
基因表达的重要性:不仅酶的存在(DNA水平)重要,其表达(RNA水平)也至关重要
治疗潜力:特定酶(如EC 6.3.1.1)可能成为炎症性肠病的治疗靶点
这些发现有力地证明,抗炎植物性食物(如草莓)的全部益处取决于特定肠道微生物酶的存在和转录活性。这解释了为什么同一食物可能对某些人有显著的健康益处,而对另一些人则效果有限——这完全取决于他们肠道中是否存在并表达这些关键酶。
这项研究通过整合全球微生物组数据和植物营养素信息,为我们理解饮食-微生物组-健康之间的联系提供了前所未有的深度和广度。研究结果不仅揭示了肠道微生物如何代谢植物性食物中的数千种化合物,还为个性化营养和精准医学开辟了新的可能性。
对益生菌和功能性食品开发的启示
这项研究对益生菌行业提出了重要挑战。虽然益生菌在各种疾病背景下已显示出疗效,但研究发现常见益生菌在代谢植物营养素方面的能力远不及我们肠道中自然存在的某些细菌。这表明,超越乳酸杆菌和双歧杆菌的益生菌开发对于更广泛地应用于人类健康至关重要。
研究人员发现,一小部分肠道细菌物种(如11种细菌的组合)可以潜在地修饰43种次级代谢相关代谢物(常见益生菌缺乏这种遗传潜力)。这为通过选择具有特定代谢能力的细菌进行发酵来开发功能性食品提供了新方向。
重新定义”健康饮食”
研究结果强调了重新审视”健康饮食”概念的重要性。饮食的有效性可能在很大程度上取决于健康微生物组的存在。对于肠道微生物组失衡的人来说,食用具有已证实健康益处的可食用植物可能不足以获得全部益处。因此,个性化营养可能需要特定食物和有益微生物的组合,或食物的离体发酵,以实现植物性饮食的全部营养潜力。
特别值得注意的是,植物营养素相关酶的比例与年龄呈负相关。这表明,老年人可能失去代谢某些植物营养素的能力,这为开发针对老年人的特定益生菌或发酵食品提供了依据,以弥补这种与年龄相关的代谢能力下降。

谷禾健康
维生素
B族维生素和维生素K是微生物和人类生理学中多种细胞过程所必需的微量营养素。传统上,这些维生素被认为主要来自饮食,但近年来的研究显示,人类肠道微生物群具备合成这些维生素的能力,提供了重要的内源性来源,这一贡献过去常被低估。
值得注意的是,微生物对宿主维生素储备的贡献正逐渐被视为维生素稳态的重要功能。在结肠中,微生物合成的维生素能够通过特定的运输机制被吸收,以发挥其生物学作用。
什么是维生素?
维生素是一类化学结构各异的有机化合物,人体只需要很少的量,但它们对维持细胞和全身的生理功能至关重要。
与提供结构或能量的宏量营养素(如碳水化合物、蛋白质和脂肪)不同,维生素主要通过作为辅因子或辅酶前体的形式发挥作用,帮助酶完成各种反应,这些反应涉及能量代谢、物质合成、氧化还原调节和细胞信号传导。
即使是轻微的维生素缺乏,也可能损害代谢的完整性,并增加患贫血、神经退行性疾病和免疫缺陷等多种疾病的风险。
分类
根据溶解性和生理特性,维生素通常分为水溶性和脂溶性两类。
• 水溶性维生素
水溶性维生素包括B族维生素(维生素B1、B2、B3、B5、B6、B7、B9和B12)和维生素C。它们通常作为辅基或辅酶参与酶促反应,尤其是在能量代谢、核苷酸合成和一碳单位转移反应中。由于其亲水性,水溶性维生素不易在体内储存。
• 脂溶性维生素
脂溶性维生素(维生素A、D、E和K)通过脂类介导的途径被吸收,并储存在脂肪组织和肝脏中,形成一个相对稳定的储备。
脂溶性维生素的功能更偏向于调节而非催化,但对于维持正常的视力、免疫调节、抗氧化防御以及钙磷平衡至关重要。比如,维生素K是凝血因子中谷氨酸残基γ-羧化反应的辅因子。
来源
• 大部分维生素依赖饮食获取或微生物产生
值得注意的是,由于人体无法合成大多数维生素,因此它们是必需的营养素。只有少数例外,如维生素D,它可以在紫外线照射下由皮肤中的7-脱氢胆固醇合成。对于其他所有维生素,人体主要依赖饮食和微生物的产生来获取。
• 肠道菌群合成维生素的能力对人体代谢非常重要
与高等真核生物不同,许多细菌物种具备从头合成必需辅因子的遗传机制,包括维生素B1、B2、B3、B5、B6、B7、叶酸(B9)、B12和甲萘醌(K2)。
其中,6种B族维生素(B1、B2、B3、B5、B6和B9)是关键的辅酶前体,这些辅酶在细菌和哺乳动物中发挥作用,如硫胺素焦磷酸(TPP)、黄素单核苷酸/黄素腺嘌呤二核苷酸(FMN/FAD)、烟酰胺腺嘌呤二核苷酸(NAD/NADP)、辅酶A(CoA)、磷酸吡哆醛(PLP)和四氢叶酸(THF)。
生物素则是一种重要的羧化/脱羧辅因子,在脂肪生成、碳水化合物和氨基酸代谢中发挥重要作用。钴胺素是B12辅酶家族(包括氰钴胺素、甲基钴胺素和腺苷钴胺素)的前体,对所有动物和许多(但并非所有)细菌物种都是必需的。
因此,人们越来越关注人体肠道菌群的合成能力,将其视为B族维生素和维生素K的潜在内源性来源,并逐渐认识到它对宿主维生素稳态的功能性贡献。
这种观念的转变得益于肠道菌群检测方法学的改进和多样化,使得谷禾能够以更高更大规模的分辨率检测和功能性地解释微生物维生素的生物合成。
本文主要关注和讨论目前对人体肠道相关细菌合成B族维生素和维生素K的理解。重点关注用于研究肠道菌群产生维生素潜力的各种方法,包括基因组学、宏基因组学等,以更好地了解微生物与宿主之间的维生素相互作用,从而改善人类健康。
人类肠道菌群是一个庞大且代谢活跃的微生物生态系统,包含丰富的基因功能库。随着新物种和功能的不断发现,我们对它的认识也在不断加深。
哪些证据说明肠道菌群合成维生素
(1)无菌大鼠 vs. 普通大鼠的对比实验
为了证明细菌生产的维生素,我们以一个关于维生素K的经典例子:
实验对象A(无菌大鼠): 肠道里没有任何细菌。
给它们吃不含维生素K的食物,它们就会生病(凝血酶原水平降低,容易出血),因为它们既没从食物中吃到维生素K,肠道里也没有细菌帮忙制造。
实验对象B(普通大鼠): 肠道里有正常的细菌。
结果: 即使吃完全一样的不含维生素K的食物,它们却很健康,凝血功能正常。
结论: 这说明普通大鼠肠道里的细菌“生产”了足够的维生素K,弥补了食物的缺失。进一步推测细菌的产量足甚至以满足生理需求。
因此以上实验表明细菌不仅仅是“产生”了维生素,而且产生的量很大,可能足够达到维持宿主健康(如正常凝血)的生理标准。
(2)人体研究
同样,在人类研究中,即使低维生素K饮食持续3-4周,也不会导致明显的维生素缺乏,除非同时使用广谱抗生素抑制肠道菌群。在这种情况下,会观察到血浆凝血酶原的明显下降,这清楚地表明了肠道菌群在维持维生素K稳态方面的重要作用。
(3)体外研究
一些计算机模拟研究也探讨了肠道菌群对宿主相关微量营养素摄入的潜在贡献,特别是关于B族维生素的膳食参考摄入量(DRI)。这些计算方法发现了能够产生多种B族维生素的微生物生物合成途径,表明肠道菌群可能对宿主的微量营养素摄入产生影响。
具体来说,尽管这仍然是推测性的,但有目前研究普遍认为肠道微生物合成的维生素B3、B6、B9和B12可能占人类相应DRI的约27%-86%。
(4)结肠转运系统
此外,在人类结肠中检测到B族维生素转运系统,进一步加强了微生物和膳食维生素可能协同作用,维持全身维生素稳态的观点。然而,由于微生物产生的维生素在结肠环境中的实际生物利用度仍未得到充分表征,需要进一步明确的研究。
!
注意局部和全身
与小肠中吸收效率明确的膳食维生素不同,微生物合成的维生素吸收取决于其化学形式和宿主转运蛋白的能力。例如,结肠中存在大量微生物合成的类咕啉素(包括维生素B12的变体),其结构常与具有生物活性的钴胺素不同,导致人类宿主的生物利用度降低。
此外,微生物合成的B族维生素主要满足自身的代谢需求,因此,与膳食来源相比,剩余部分(无论是主动分泌还是通过细菌细胞自发裂解释放)所占比例可能较小。
同时,微生物产生的维生素在塑造局部肠道环境(特别是在肠腔内和黏膜界面)中具有重要的生物学意义。已知这些维生素可以支持结肠上皮细胞的代谢需求,影响屏障完整性,调节局部免疫反应,并发挥抗氧化和保护作用。
谷禾数据库整理了大样本人群微生物基因组参考集合的基因组学研究,初步细致地刻画了肠道微生物群中各个成员合成维生素的潜力。
▸ 谁能自己造维生素?
把这些信息汇总到“门”(phylum)这一分类层级时,拟杆菌门(Bacteroidota)和假单胞菌门(Pseudomonadota)中“维生素原养型”(prototrophic,能从头合成某种化合物)的物种比例更高。
这两个门里包含能够合成几乎所有B族维生素的物种,但维生素B12(钴胺素)是个例外:大约只有30%的拟杆菌门基因组和约42%的变形菌门基因组显示出合成B12的潜力。
• 拟杆菌属和大肠杆菌是重要的B族维生素生产者
Bacteroides(拟杆菌属): 一些拟杆菌属的成员可以合成全部八种B族维生素。
E.coli(大肠杆菌), Phocaeicola dorei, Bifidobacterium(双歧杆菌属), Segatella copri(以前叫Prevotella copri): 这些也是肠道中B族维生素的重要生产者。
• 放线菌和芽孢杆菌中能产生维生素的较少
相对地,放线菌门(Actinomycetota)和芽孢杆菌门(Bacillota)中维生素“营养缺陷型/需外源型”(依赖外部来源满足维生素需求)的成员比例更高。
注:研究数量有限,仅供参考
▸ 肠型可能影响维生素的合成能力
• 不同肠型擅长合成的维生素不同(仅供参考)
肠道菌群的组成(肠型)会影响维生素的合成能力。
肠型1(以Bacteroides为主): 更擅长合成生物素、核黄素和泛酸。
肠型2(以Prevotella copri为主): 更擅长合成硫胺素和叶酸。
• 谁是维生素“消费者”?
Faecalibacterium prausnitzii(普拉梭菌): 它会消耗多种B族维生素,尤其是核黄素,核黄素能帮助它在肠道内抵抗氧化。
尽管基因组研究已阐明单个菌株或基因组的生物合成能力,现有方法仍较少将这一视角扩展至整个微生物群落,因此尚难在人群层面绘制维生素生物合成潜力图谱。
现有研究和谷禾的研发表明,特定微生物类群通过其生物合成能力模式和维生素利用策略,在塑造肠道维生素“生态”中起着关键作用。深入理解不同类群对维生素代谢的特异性贡献对于阐明微生物群与宿主营养状态的功能性相互作用至关重要,这些认识还可能指导基于微生物组的干预策略设计,以优化维生素可获得性并解决营养缺乏问题。
在此背景下,比如用能分泌叶酸的乳酸菌做发酵剂,生产叶酸强化乳制品;也在筛选肠道来源的产叶酸菌作为潜在益生菌,用于改善维生素供应。这些例子说明:关于微生物维生素代谢的知识可以转化为食品和治疗策略,最终为利用肠道微生物群有益代谢潜力的营养学方法铺平道路。
尽管人们越来越认识到人类肠道菌群具备合成维生素的能力,但最近的研究强调,肠道菌群中维生素的可用性并非仅仅由生物合成能力决定,还受到微生物群落内部以及宿主处复杂的代谢相互作用的影响。
这些动态过程包括微生物之间的交叉代谢、维生素摄取的竞争以及宿主介导的吸收,所有这些因素共同塑造了最终的营养产出。
▸ 微生物—微生物互作
在微生物层面,维生素的交叉喂养代谢源于不同分类群之间生物合成能力的不均匀分布,且这种差异不仅存在于亲缘关系较远的类群间,也体现在同一属内的不同物种之间。
最近的研究中,研究人员利用 SEED 平台分析了一个包含2000多个经过人工校对的人类肠道细菌基因组的参考数据集。该平台整合了所有已知和推断的生物合成及补救途径组件,包括酶和转运蛋白。这一方法通过预测不同分类等级中原养型(可自身合成)和营养缺陷型(需外部获取)菌株的比例,实现了对维生素生物合成潜力变异性的定量评估。这些跨越不同维生素和分类级别的保守性与变异性模式,为微生物群落中的潜在代谢相互作用奠定了基础。
• 原养型微生物的产物可能支持缺陷型的生长
当原养型和营养缺陷型微生物共存于同一生态位时,这种差异可能导致代谢相互依赖,即原养型微生物释放的代谢产物或中间体能够支持营养缺陷型微生物的生长和代谢功能。
一个明显的例子是维生素B12,这一结构复杂且代谢成本高昂的辅助因子。由于完整生物合成途径需高昂的代谢成本和遗传负担,只有少数厌氧肠道细菌(主要属于厚壁菌门和放线菌门)能够自主生产维生素B12。
然而,这种辅助因子对许多群落成员至关重要,众多成员已进化出高亲和力的转运系统以从环境中获取。例如,Anaerobutyricum hallii(以前称E.hallii)是一种特征明确的丁酸产生菌,能够合成维生素B12,而嗜黏蛋白阿克曼氏菌(Akkermansia)则通过共生利用该维生素进行丙酸的合成。
同样,在肠道微生物群落模型中,缺乏叶酸合成能力的Roseburia intestinalis M50/1生长受到经体外验证的叶酸原养型菌株双歧杆菌Bifidobacterium bifidum CNCM I-3650的益处。这些发现表明,原养型细菌可以通过主动交换代谢物和微量营养素,或通过被动机制(如细胞裂解并释放胞内维生素),来缓解共存菌株的营养缺陷。
这些以维生素为媒介的相互作用不仅是合作性的,竞争也在塑造人类肠道维生素可用性方面起着关键作用。
维生素的可用性受微生物间相互作用等的影响
• 肠道微生物群会竞争有限的维生素资源
微生物通过产生维生素结合蛋白和利用特定转运蛋白,与其他群落成员竞争有限的维生素资源。因此,肠道环境可视为一个严格调控的微量营养素市场,其可用性由生物合成能力和摄取效率共同决定。
例如,多形拟杆菌(Bacteroides thetaiotaomicron)表达三种功能性、同源的维生素B12转运系统以获取钴胺素。这些转运蛋白对不同类咕啉类似物表现出独特的特异性,这可能为其在竞争激烈的人类肠道环境中提供选择性优势,同时也体现了肠道微生物为确保获得必需辅助因子所采用的复杂策略和巨大努力。
▸ 微生物—人体互作
宿主与微生物之间的相互作用使这一网络愈加复杂。实际上,宿主通过免疫调节、结肠吸收、粘液分泌和饮食等方式,间接影响维生素的动态变化,从而影响微生物的组成和功能。
• 不同年龄的肠道菌群组成影响维生素合成能力
最近的研究显示,年龄和地理起源等宿主因素显著影响人类肠道菌群的维生素生物合成潜力。具体而言,肠道菌群的测序分析表明,婴儿期维生素B9(叶酸)和K2的生物合成途径非常普遍,这与能合成叶酸的双歧杆菌属(Bifidobacterium spp.)的主导地位以及能够产生维生素K2的大肠杆菌(E.coli)的存在相一致,这些都是早期肠道微生物组的特征。
另一项研究将人群按年龄分组(从几个月大到80岁以上),并将微生物特征与年龄进行关联分析。结果显示,新生儿组(0-4岁)的微生物与参与硫胺素(维生素B1)和烟酸(维生素B3)生物合成的酶呈正相关,表明这些维生素路径与生命早期相关,这与老年组形成对比。
成人的肠道菌群则富含Bacteroides和Prevotella copri,展现出更广泛的代谢能力,因为这些类群通常是维生素B的原养型生物,具备多种B族维生素的完整生物合成途径。
• 不同地区的饮食和生活方式导致维生素合成能力差异
此外,新兴证据表明,地理起源导致全球不同人群肠道菌群在B族维生素生物合成能力上的差异,可能与饮食和生活方式相关因素影响肠道菌群的组成和功能有关。
在这个复杂的生态和代谢网络中,结合本地大数据库明确哪些微生物类群在结肠中参与维生素生物合成、这些维生素是否被宿主吸收以及哪些因素影响其产生,对于理解微生物群对宿主的营养贡献至关重要。这也有助于设计针对微生物群的策略(如益生元、益生菌或工程菌群),以改善维生素状况。
1
维生素B1(硫胺素)
▸ 生物学功能与重要性
硫胺素(维生素B1)是一种水溶性维生素,作为辅酶参与多个核心代谢过程。在活性磷酸化形式TPP(焦磷酸硫胺素)下,维生素B1是碳水化合物代谢中多种酶的关键辅因子,包括丙酮酸脱氢酶、α-酮戊二酸脱氢酶和转酮醇酶,这些酶是三羧酸循环(TCA)和磷酸戊糖途径的关键参与者。
通过这些途径,硫胺素有助于能量生成、氧化还原平衡、核苷酸和脂质的生物合成,并间接参与神经递质的代谢。
▸ 微生物合成与已知微生物贡献者
细菌中的硫胺素生物合成遵循模块化途径,涉及两个部分的分别合成:4-氨基-5-羟甲基-2-甲基嘧啶磷酸(HMP-P)和4-甲基-5-(β-羟乙基)噻唑磷酸 (THZ-P)。随后,这些部分由 ThiE 酶偶联生成磷酸硫胺素,最后转化为活性的 TPP 形式。基因组调查显示,相当大比例的肠道相关细菌基因组,包括拟杆菌属(Bacteroides)、普雷沃氏菌属(Prevotella)、肠球菌属(Enterococcus)和双歧杆菌属(Bifidobacterium)等主要菌属的代表,都拥有从头合成磷酸硫胺素所需的完整基因库。
然而,许多肠道共生菌对硫胺素是营养缺陷型的,依赖于外源性硫胺素或诸如 HMP-P 之类的中间体,这些物质可以从环境中回收或在微生物之间交换。在这些维生素B1营养缺陷型细菌中,已鉴定出不同的补救策略:有些菌株可以回收两种硫胺素中间体,而另一些则缺乏其中一条生物合成支路,依赖外部来源提供相应的缺失化合物,还有一部分完全依赖于摄取预先形成的硫胺素。
值得注意的是,所有营养缺陷型变体都保留了一定的补救能力,反映了对不完整生物合成潜力的多样化进化适应。
有趣的是,普拉梭菌(F. prausnitzii)在将丙酮酸转化为乙酰辅酶A的代谢过程中需要这种维生素,这是丁酸生物合成途径的一部分,突显了维生素B1在肠道菌群成员生产短链脂肪酸(SCFA)中的关键作用。
// 小结
总结来说,维生素B1(硫胺素)不仅是你身体的“能量充电宝”,也是将食物(碳水化合物)转化为能量所必需的“点火器”。没有它,我们的细胞无法有效产生能量,也无法制造DNA和脂肪,甚至会影响神经系统的信号传递。
肠道里住着很多细菌,其中一些是“生产者”。比如拟杆菌和双歧杆菌等常见菌,可以自己从头制造维生素B1,不仅满足自己,可能还能分给别人。还有很多细菌是“消费者”(营养缺陷型),它们自己造不出维生素B1,或者只能造出一半。
有的细菌完全依赖别人,必须从环境里捡现成的维生素 B1 吃。有的细菌只能造一半零件,需要从邻居那里借另一半零件来组装。说明为了生存,细菌逐步进化出复杂的“借用”和“互助”关系。
2
维生素B2(核黄素)
▸ 生物学功能与重要性
核黄素(维生素B2)是两种主要黄素辅酶——黄素单核苷酸(FMN)和黄素腺嘌呤二核苷酸(FAD)的前体。这两种辅酶在核心代谢中广泛参与氧化还原反应,包括电子传递链、脂肪酸氧化、氨基酸分解代谢,以及维生素B6和B9的激活反应。
此外,核黄素通过支持谷胱甘肽还原酶的活性和调节氧化应激,有助于维持抗氧化防御系统。
▸ 微生物合成与已知贡献者
细菌中的核黄素生物合成涉及一组保守基因(ribA、ribB、ribC、ribD和ribE),这些基因将三磷酸鸟苷(GTP)和5-磷酸核酮糖转化为核黄素,随后核黄素被磷酸化为FMN,再腺苷酸化为FAD。
最近的基因组重构研究显示,完整的操纵子在肠道共生细菌中广泛存在,包括拟杆菌属(Bacteroides)、肠球菌属(Enterococcus)和多种乳杆菌。尤为重要的是,乳酸乳球菌(Lactococcus lactis)、植物乳杆菌(Lactiplantibacillus plantarum)和罗伊氏乳杆菌(Limosilactobacillus reuteri)已被提议作为天然生物强化剂,用于提高酸奶、奶酪和发酵乳制品中的核黄素浓度。
然而,基因组分析显示,芽孢杆菌门(Bacillota)和放线菌门(Actinomycetota)中的一些成员缺乏完整的核黄素生物合成操纵子。有趣的是,许多这些核黄素缺乏型细菌编码高亲和力的核黄素转运系统,如RibU(ECF家族)和RibXY(ABC超家族),显示出它们对核黄素衍生辅助因子(如FMN和FAD)的强烈需求。
除了营养作用外,微生物生成的核黄素还参与调节宿主粘膜表面的免疫反应,其关键机制涉及粘膜相关恒定T(MAIT)细胞,这些细胞在识别由非多态性MHC I类相关分子MR1呈递的核黄素生物合成中间体后被激活。该途径建立了微生物维生素B2代谢与粘膜免疫监视之间的直接联系,强调了微生物核黄素生成在宿主-微生物相互作用中的更广泛生理相关性。
// 小结
维生素B2(核黄素)不仅是身体必需的营养素,也是肠道细菌与免疫系统之间的沟通桥梁。进入体内后,维生素B2转化为两种“超级助手”(FMN和FAD),参与无数化学反应,尤其是在将脂肪和蛋白质转化为能量,以及协助产生其他维生素(如B6和叶酸)。它还能帮助抵抗“氧化压力”,保护身体免受损伤。
肠道细菌也能合成B2,许多细菌(如某些拟杆菌和乳酸菌)具备特定基因,能够自己生产维生素B2。然而,并非所有细菌都能合成B2;一些细菌缺乏这种能力,但进化出强大的“抢夺”能力(高亲和力转运系统),能够从环境中获取B2以维持生存,显示出B2对细菌生存的重要性。
意外发现是,细菌制造B2的过程中还会产生一些中间产物,这些产物能够激活免疫系统中的特殊巡逻兵“MAIT细胞”。当MAIT细胞探测到细菌生成B2时,这些细胞会被激活,像是细菌发送的信号,通知免疫系统“我在这里”。这不仅帮助身体识别细菌,还启动免疫监视,保护肠道粘膜的健康。
3
维生素B3(烟酸)
▸ 生物学功能与重要性
维生素B3,即烟酸,包含两种生物活性形式:烟酸(NA)和烟酰胺(NAM)。这两者都是烟酰胺腺嘌呤二核苷酸(NAD+)和烟酰胺腺嘌呤二核苷酸磷酸(NADP+)的前体。这两种二核苷酸是参与氧化还原反应、能量代谢、DNA 修复和细胞信号传导的重要辅助因子。
NAD+是糖酵解、三羧酸循环和氧化磷酸化中的关键辅酶,而NADP+对合成代谢途径和抗氧化防御至关重要。
▸ 微生物生物合成与已知贡献者
与大多数其他B族维生素不同,烟酸可以在体内(主要在肝脏)内源性合成,通过犬尿氨酸途径将色氨酸转化为喹啉酸,后者是NAD+生物合成的关键中间体。然而,这一途径通常不足以满足日常需求,因此饮食摄入或微生物的贡献显得尤为重要。
在肠道中,许多共生细菌拥有一条替代的烟酸从头合成途径,从天冬氨酸开始生成喹啉酸,再转化为烟酸单核苷酸,最终合成NAD+。这一生物合成途径广泛分布于假单胞菌门(Pseudomonadota)和拟杆菌门(Bacteroidota)中,尽管在属和种的水平上存在显著差异。大肠杆菌(E. coli)、脆弱拟杆菌(B.fragilis)和多种双歧杆菌(Bifidobacterium)被确认是肠道环境中的烟酸生成者。
然而,并非所有菌种都编码完整的生物合成基因,许多细菌依赖烟酸补救途径,利用宿主或微生物衍生的前体。最新证据强调了宿主与其肠道菌群之间维生素B3前体的双向交换,支持共享的NAD+代谢。
尽管饮食中的烟酸前体大多在上消化道/小肠中被吸收,但结肠中的微生物可以利用可发酵纤维(如菊粉)或宿主细胞代谢释放的NAM来合成NAD+。微生物来源的NAD+随后转化为NAM和NA,这些物质能够被宿主肠道组织吸收,并在宿主细胞内用于再生NAD+。这突显了一个动态循环,即微生物活动与宿主来源底物共同维持循环中NA水平,即使在缺乏直接饮食摄入的情况下。
// 小结
维生素B3是身体细胞正常运转不可或缺的成分。你可以把它想象成制造“电池”的原料。它主要有两种形式(烟酸和烟酰胺),在体内会变成两种超级重要的物质(NAD+和NADP+)。这两种物质负责帮我们把食物变成能量、修复受损的 DNA,以及保护细胞不“生锈”(抗氧化)。我们的肝脏可以利用蛋白质中的某种成分(色氨酸)自己合成一点维生素 B3,但这通常不够用,大部分需要通过食物摄入。如果你吃不够,你肚子里的细菌能帮大忙。
在你肠道里的许多细菌(比如大肠杆菌和双歧杆菌)拥有特殊的“生产线”,可以用完全不同的原料(天冬氨酸)制造出维生素 B3。
细菌还可以利用我们吃下去的膳食纤维(比如菊粉)或者我们身体代谢剩下的废料,制造出 NAD+。然后它们把这些成品分解成我们可以吸收的形式(烟酸或烟酰胺),这就相当于细菌在结肠里给我们“供货”。我们身体产生的某些前体物质也能被细菌利用。
4
维生素B5(泛酸)
▸ 生物学功能与重要性
维生素B5(泛酸)是辅酶A(CoA)的生化前体,辅酶A作为核心代谢辅助因子,参与广泛的生物过程,包括三羧酸循环(TCA)、脂肪酸合成与β-氧化以及氨基酸代谢,主要在能量生成和合成代谢中充当酰基载体。此外,胆固醇、类固醇激素、血红素和乙酰胆碱的生物合成也需要辅酶A。
维生素B5对于维持细胞能量平衡和代谢功能至关重要。
▸ 微生物生物合成与已知贡献者
细菌可以通过一条从头合成途径来制造泛酸,该途径将泛解酸与β-丙氨酸结合在一起。这条途径由一组高度保守的基因簇(panB、panC、panD、panE)编码,合成出的维生素随后会被转化为辅酶 A。
在人类肠道菌群中,基因组分析显示,几乎所有的假单胞菌门(Pseudomonadota)和拟杆菌门(Bacteroidota)都具备合成泛酸的遗传能力,而受评估的厚壁菌门(Bacillota)和放线菌门(Actinomycetota)成员中,只有不到一半编码了完整的泛酸合成途径。不过,许多无法自身合成泛酸的菌株(营养缺陷型)仍然保留了将外源性泛酸转化为辅酶A的酶机制,这使它们即使无法从头合成,也能完成基本的代谢功能。
正因为假单胞菌门和拟杆菌门普遍具备这种合成能力,所以在成年人的肠道菌群中,尤其是典型的以拟杆菌为主的肠型中,维生素B5的生物合成途径是最常见的代谢途径之一。
// 小结
维生素 B5 之所以重要,是因为它是我们体内一种叫“辅酶 A”的关键物质的原材料。你可以把辅酶 A 想象成细胞工厂里的“搬运工”和“点火器”。 它是身体燃烧脂肪、分解蛋白质和碳水化合物产生能量的过程中不可或缺的一环。身体制造胆固醇、性激素、血红蛋白(血液中携带氧气的成分)以及神经传导物质时,都离不开它。
5
维生素B6(吡哆醇)
▸ 生物学功能与重要性
维生素B6是由六种可相互转化的形式组成,包括吡哆醇(PN)、吡哆醛(PL)、吡哆胺(PM)及其磷酸化形式,其中5′-磷酸吡哆醛(PLP)是最活跃的形式。PLP作为辅酶参与了约140种酶促反应,使维生素B6成为功能多样且复杂的微量营养素之一。除了糖原磷酸化酶外,几乎所有依赖PLP的酶都涉及氨基化合物的生化过程,尤其是氨基酸代谢。
▸ 微生物生物合成与已知贡献者
在人类肠道中,PLP通过两条截然不同的途径合成:较长的依赖脱氧木酮糖5-磷酸(DXP)的途径,这在拟杆菌门(Bacteroidota)和假单胞菌门(Pseudomonadota)中普遍存在;以及较短的不依赖DXP的途径,这在放线菌门(Actinomycetota)中更为常见。
基因组分析表明,PLP的产生在肠道细菌中分布不均。尽管预测双歧杆菌属(Bifidobacterium)、拟杆菌属(Bacteroides)和普雷沃氏菌属(Prevotella)能合成PLP,但许多著名的肠道共生菌,如韦荣氏球菌(Veillonella spp.)、粪肠球菌(Enterococcus faecalis)、普拉梭菌(F.prausnitzii)和Roseburia inulinivorans则被认为是营养缺陷型(无法自行合成)。这些生物体通常携带补救途径的遗传特征,使它们能够将环境中的B6维生素形式转化为可用的PLP。
6
维生素B7(生物素)
▸ 生物学功能与重要性
生物素,也被称为维生素B7,是一种含硫的水溶性维生素。它作为羧化酶的辅酶,参与关键的代谢途径,包括脂肪酸代谢、氨基酸代谢、碳水化合物代谢、聚酮化合物生物合成以及尿素利用。生物素通过赖氨酸残基与靶酶共价结合,促进二氧化碳转移反应,这对维持能量稳态和脂质代谢至关重要。
▸ 微生物生物合成与已知贡献者
细菌中的生物素合成通过两条截然不同的途径进行。在大肠杆菌(Escherichia coli)中,生物素是通过一种改良的脂肪酸合成途径合成的,该途径依赖于bioC和bioH基因,这是一条规范的且在生物素合成的肠道细菌中广泛存在的路径。相比之下,枯草芽孢杆菌(Bacillus subtilis)则利用以bioW基因为中心的独立途径,这条依赖bioW的路线在Bacillota成员中更为常见。
根据最近的基因组调查,所有分析的放线菌门(Actinomycetota)基因组均缺乏从头合成生物素所需的必需基因,这种特征在厚壁菌门也很少见。与其营养缺陷特征相符,大多数放线菌门细菌编码高亲和力的生物素摄取系统,强调了它们对环境生物素来源的依赖,并表明存在选择性机制以确保高效清除这种必需辅因子。
7
维生素B9(叶酸)
▸ 生物学功能与重要性
维生素B9包括一类与四氢叶酸(THF)结构相关的水溶性化合物,THF在“一碳代谢”中作为辅酶发挥作用。THF及其衍生物对于许多反应中转移一碳单位至关重要,这些反应涉及嘌呤、胸苷酸和多种氨基酸的合成,以及核苷酸的合成、DNA的复制、修复和甲基化。
因此,叶酸在核苷酸合成、表观遗传调控和细胞分裂中发挥关键作用,尤其在妊娠和婴儿期等快速生长阶段更为重要。
▸ 微生物生物合成与已知贡献者
细菌从头合成叶酸需要先生成并缩合两种关键中间体:二氢蝶呤三磷酸(DHPPP)和对氨基苯甲酸(pABA)。这些前体通过不同的代谢途径生成,最终汇合形成二氢叶酸(DHF),随后被还原为具有生物活性的四氢叶酸(THF)。
在肠道微生物中,适应婴儿肠道的物种(如B.bifidum、B.longum和B.breve)被认为是重要的叶酸生产者。这些菌株已能在培养基中积累叶酸,表明它们可能为婴儿肠道的叶酸库作出贡献。然而,双歧杆菌通常需要外源补充pABA才能合成叶酸,表明它们在该前体上存在一定程度的“营养缺陷”。
乳杆菌也是能产叶酸的细菌。例如,德氏乳杆菌(Lactobacillus delbrueckii)、瑞士乳杆菌(Lactobacillus helveticus)、罗伊氏乳杆菌(Limosilactobacillus reuteri)、清酒乳杆菌(Latilactobacillus sakei)和植物乳杆菌(Lactiplantibacillus plantarum)的菌株在pABA存在的生长环境中能合成叶酸。
值得注意的是,植物乳杆菌表现出无需补充pABA就能稳定产生叶酸的能力。许多研究评估了肠道微生物对动物宿主叶酸摄入的贡献,已证实肠道细菌合成的叶酸可以在结肠被吸收并被宿主利用。基于这一发现,研究人员探讨了来自各种发酵食品的乳杆菌作为发酵剂生产叶酸强化乳制品,同时也探索将人体肠道来源的乳杆菌作为产叶酸益生菌。
8
维生素B12(钴胺素)
▸ 生物学功能与重要性
维生素B12(钴胺素)是一种含钴的辅因子,也是自然界中体积最大、结构最复杂的非聚合生物分子之一。在人体内,它作为辅酶参与两项关键的线粒体反应:
1)甲基丙二酰辅酶A变位酶(methylmalonyl‑CoA mutase):参与丙酸代谢与能量产生;
2)蛋氨酸合成酶(methionine synthase):在叶酸循环中把同型半胱氨酸再生为蛋氨酸,起关键作用。
在B12的多种形式中,氰钴胺(cyanocobalamin)化学性质最稳定;它被吸收后会经过酶促加工,转化为具有活性的辅因子形式:甲基钴胺(Me‑Cbl)和腺苷钴胺(Ado‑Cbl)。
▸ 微生物生物合成与已知贡献者
钴胺素的生物合成过程非常复杂且耗能,涉及30多个基因,可走需氧或厌氧两条途径,最终都汇合到咕啉环(corrin ring)形成以及钴离子螯合的步骤。
该合成能力主要局限于某些细菌类群,许多肠道微生物并不具备。大规模基因组分析显示,约60%–80%的人类肠道共生菌缺乏合成B12所需的完整基因组合,例如双歧杆菌属(Bifidobacterium)、普雷沃菌属(Prevotella)以及Bacteroides thetaiotaomicron等。
相反,Propionibacterium freudenreichii、梭菌属(Clostridium)、脆弱拟杆菌(Bacteroides fragilis)、阿克曼菌(Akkermansia muciniphila)以及部分Bacillota和放线菌门(Actinomycetota)细菌,编码了完整或近乎完整的B12合成操纵子。
但由于合成B12非常耗能,很多细菌进化出了替代策略来获得这种辅因子及其类似物(咕啉类/钴胺素类似物,corrinoids):例如回收前体或直接从环境/其他菌那里摄取完整分子,从而在肠道微生物群中形成合作与资源共享。
不过在人类体内,结肠产生的B12吸收效率很低,因此微生物来源B12对全身B12水平的直接贡献仍不明确。
// 小结
B12对人体很关键:它参与能量代谢的一条关键反应,以及“叶酸循环/同型半胱氨酸→蛋氨酸”的关键步骤。补剂里常见的氰钴胺只是稳定形态:进入人体后会被转换成真正起作用的两种活性形式(甲基钴胺、腺苷钴胺)。
肠道菌并非都能制造B12:多数常见肠道共生菌缺乏完整“生产线”,只有部分细菌具备较完整的合成基因。很多细菌选择“省钱模式”:不自己合成,而是从环境或其他细菌那里“捡现成/回收零件”。未来还需要继续加强研究。
9
维生素K2(甲萘醌,Menaquinones)
▸ 生物学功能与重要性
维生素K主要有两种形式:一种是叶绿醌(K1),存在于绿色叶菜中;另一种是甲萘醌(K2),它是一类多样的异戊二烯衍生物,主要由细菌产生。甲萘醌(MK-n)的差异在于其异戊二烯侧链长度不同,这会影响其生物利用度和在组织中的分布。
在细菌细胞中,MK-n在细胞质膜的电子传递中起核心作用,支持形成芽孢的过程,并与致病菌的毒力有关。在人类中,维生素K2不足与骨质疏松、心血管疾病以及胰岛素敏感性下降有关。
▸ 微生物生物合成与已知贡献者
细菌中的甲萘醌合成途径涉及两个关键部分的汇合:萘醌环结构与异戊二烯侧链。这两部分首先通过不同代谢途径分别生成,随后再结合形成醌的核心结构。之后,异戊二烯侧链通过逐步添加异戊二烯单位而延长,产生侧链长度不同的甲萘醌变体。这种结构多样性带来功能上的分工,也可能有助于不同细菌适应环境。
目前,若干能产生甲萘醌的细菌菌株已作为工业食品发酵的发酵剂使用,并显示出可提高培养基中维生素K含量的能力。例如,将乳酸乳球菌(Lactococcus cremoris、L.lactis)和乳明串珠菌(Leuconostoc lactis)在复原奶粉或豆奶中培养时,可产生长链MK,并且水平几乎接近成人每日适宜摄入量。这些发现支持:发酵食品可以成为维生素K2的重要膳食来源。
// 小结
总结来说,维生素K有两大类:K1主要来自绿叶菜;K2主要由细菌合成。K2不是单一物质,而是一系列“MK-n”家族成员,侧链长短不同,会影响人体吸收与在体内去向。
对细菌来说,K2是能量代谢(电子传递链)的关键分子,还与芽孢形成和致病性相关。对人来说,K2不足可能与骨质疏松、心血管问题以及胰岛素敏感性变差有关。有些用于发酵的乳酸菌在牛奶或豆奶里能产出一定量的K2,说明发酵食品可能是补充K2的一个重要来源。
▸ 分子测序方法
近一二十年来,高通量测序、多组学与计算建模的重大进展,极大提升了我们对人类肠道微生物组的理解,包括其参与维生素生物合成的作用。
通过测序的基因组与生物信息学绘制“群落层面”的维生素合成潜力。
分享一些有趣关联:高纤维饮食人群的肠道微生物组更富集叶酸与核黄素“潜在生产者”,而西式饮食与更高的“维生素合成缺失”(更依赖外源获取)相关。不同年龄也有差异:成人整体合成潜力更强;婴儿相对更富集维生素K2相关的甲萘醌与叶酸合成潜力。
在临床队列中,2型糖尿病患者的维生素K2及多种B族维生素合成基因丰度下降,提示微生物维生素代谢可能与健康状态相关。
有研究发现炎症性肠病(IBD)患者的钴胺素(维生素B12)与硫胺素(维生素B1)合成相关基因表达显著降低,说明疾病本身会影响微生物维生素代谢的“实际运行”。
!
不足之处
但需要强调的是:无论是16S rRNA 测序还是宏基因组测序,目前主要评估的是菌群的潜在功能能力”——其推断结果高度依赖检测机构所采用的整合菌群基因与代谢通路数据库。现有数据库仍可能存在注释不全或偏倚:替代通路、新发现酶及非模式菌的功能注释缺口更为常见,从而带来功能推断的不确定性。
现阶段,基因组学层面的“检出”并不能直接证明相关基因已在体内表达、对应酶活性确实发生并产生了可观的生理效应;这类问题往往需要结合转录组、蛋白组(必要时包括代谢组)等证据链来支撑。
因而,更理想的路径是将菌群组学结果与人群膳食摄入、临床指标与其他组学数据联合分析,才能更清晰地反映人体维生素“合成—转化—吸收—利用”的真实状态;这也与谷禾健康正在持续研发与构建的综合模型思路一致。
基于上述限制,在谨慎选择菌群检测机构的同时,也应对菌群评估报告中的营养相关指标保持审慎解读:建议结合受试者的症状体征、既往史与用药/补充剂使用、饮食结构及实验室检查结果,进行综合判断与个体化干预,避免将“功能预测”直接等同于“临床结论”。
总体而言,基因组与生物信息学为绘制肠道微生物合成、改造或获取维生素的遗传潜力提供了强有力的入口。它们无法单独给出真实产量与功能输出,但能指导饮食和了解评估肠道菌群代谢维生素的合成和利用潜力,并在群落尺度上建模预测代谢。
▸ 培养方法
测序等基于序列的方法极大拓展了我们对微生物代谢的认识,但要在实验上“坐实”某个微生物是否真的能合成维生素,培养(culture-based)方法仍然不可或缺。通过培养,我们可以直接测试代谢能力、分离出能产维生素的菌或需要外源维生素才能生长的菌(营养缺陷/依赖型),并在营养成分严格可控的条件下设计实验来研究微生物之间的相互作用。
在实验室条件下评估微生物维生素生物合成潜力的一种经典且至今仍广泛使用的方法,是利用“营养缺陷指示菌株”(auxotrophic indicator strains)。这类菌的生长依赖某一种特定B族维生素的外源供给,因此可以通过其生长反应来定量推断样品中该维生素的可用量。这种测定已大量用于研究乳酸菌和双歧杆菌产生叶酸、硫胺素(维生素B1)和钴胺素(维生素B12)的能力。
研究者也会用体外体系直接观察微生物之间的“维生素互喂/交叉供养”(cross-feeding):把一种需要维生素的菌和一种能合成维生素的菌配对共培养。比如在缺乏叶酸的培养基中,R. intestinalis 与 B. bifidum 共培养时能够生长,因为后者会被动向环境释放叶酸合成的中间产物。类似地,在没有外源硫胺素的情况下,Roseburia faecis 能维持对硫胺素依赖的F.prausnitzii 的生长,说明物种间确实发生了有效的营养共享。
为了更接近肠道微生物生态系统的复杂性并延长观察时间,更复杂的体外培养平台(如基于生物反应器的发酵系统)也被用于相关研究。这类封闭、动态系统可以精确调控环境参数(如pH、滞留时间、底物输入),并能长期维持稳定、可重复的“合成”微生物群落。尽管它们已被广泛用于模拟结肠条件下的微生物生态与代谢产物研究,但直接用于研究肠道菌B族维生素的合成与交换目前仍相对有限。不过,这些系统为未来在更接近生理条件的环境中捕捉微生物营养互作提供了很有前景的框架。
!
局限性
但培养方法也有先天局限:相当一部分肠道微生物在标准实验室条件下仍难以培养,限制了我们对一些关键群落成员维生素代谢能力的评估。尽管如此,培养仍是功能微生物学的基石,因为它能对维生素的合成、转运以及物种间交换进行直接的实验验证。
▸ 分析化学方法与代谢组学
代谢组学在阐明微生物代谢方面居于补充重要地位:它通过检测和定量代谢产物,直接提供“微生物实际产生了什么”的证据,因此适合用来研究宿主—微生物以及微生物—微生物之间的代谢互作,包括微量营养素(如维生素)的动态变化。
代谢组学方法大体分为靶向和非靶向两类。靶向代谢组学使用已知标准品,对特定代谢物进行精确定量,例如B族维生素及其衍生物/不同活性形式(vitamers)。
非靶向代谢组学则试图给出更全面的代谢物“全景图”,用于发现意料之外或新的代谢变化。
目前,对代谢物的检测、鉴定与定量能力依赖于分析化学平台的快速发展,包括:
-气相色谱(GC)
-液相色谱(LC)
-高/超高效液相色谱(HPLC、UPLC)。
其中,LC尤其是与质谱联用(LC‑MS)已成为最通用、应用最广的代谢组学工具之一。LC‑MS通常配合电喷雾电离(ESI),可以在食品、牛奶、药物制剂、婴儿配方奶粉、血液和人类粪便等复杂样品中,同时分析多种B族维生素和甲萘醌(维生素K2的一类化合物)。
LC‑MS的一种更高级形式是液相色谱‑串联质谱(LC‑MS/MS)。它通过两级连续的质量分析显著提高灵敏度和特异性,不仅能检测痕量化合物,还能区分结构非常相近的分子,比如同一种维生素的不同同分异构体/不同活性形式。例子是:近期已有研究用LC‑MS/MS实现了维生素B12三种活性形式的同时精准定量,体现了该技术在解析微量营养素“精细多样性”方面的价值。
!
注意
尽管这些方法已成功用于从复杂食物和生物样品中区分并定量多种B族维生素的活性形式,但它们用于研究人体肠道中“微生物自身合成维生素”的工作仍相对有限。不过,由于其高特异性和高灵敏度,这类技术很有潜力成为未来研究的重要工具。
▸ 临床前模型
要真正理解肠道菌群的维生素代谢是否真的有功能意义、能否影响维生素稳态,并为调控菌群来改善健康提供依据。因此,需要把“微生物活动”和“宿主反应”放在同一个整体里研究的宿主相关模型。
▸ 为什么常用鼠类模型?
研究肠道菌群与宿主互作最常用的是动物模型,尤其是小鼠和大鼠。它们的优势是环境和变量可控,能系统地观察“菌群—维生素—宿主”的因果关系。
例如:在叶酸缺乏的大鼠中,给予能大量产叶酸的双歧杆菌(如 Bifidobacterium adolescentis、B.pseudocatenulatum)可以帮助恢复叶酸水平。这类实验说明:微生物合成的维生素有可能在体内发挥营养补充作用。
此外,由于B族维生素与多种人类疾病相关,鼠类也常用来模拟疾病状态,以研究菌群维生素代谢对病理过程的影响。
▸ 维生素合成益生菌:对炎症性疾病的潜力
能合成B族维生素的益生菌在治疗炎症性疾病(尤其是肠道炎症)方面显示出潜力:产核黄素(维生素B2)的 Lactiplantibacillus plantarum CRL2130:在小鼠溃疡性结肠炎和肠黏膜炎模型中,可降低炎症细胞因子、改善症状。
产叶酸的 Streptococcus thermophilus(CRL808、CRL415):在化学诱导的小鼠模型中,可减轻肠黏膜炎症。
若联合使用“产叶酸 + 产核黄素”菌株:在乳腺癌化疗动物模型中,既能提高化疗效果,又能降低如黏膜炎等副作用。
▸ 对神经系统也可能有帮助(动物证据)
一些产B族维生素的菌株在神经保护方面也有动物实验支持:
在帕金森病小鼠模型中,口服产核黄素的L.plantarum CRL2130 或产硫胺素(维生素B1)的 L. plantarum CRL1905,可改善运动协调能力,并减少多巴胺能神经元损失。
这些效果与直接补充商业维生素相当,提示“微生物在体内合成的维生素”确实影响人体维生素的浓度和利用,并能在功能上支持宿主的营养与生理过程。
!
局限性
鼠类模型很有价值,但也要谨慎外推到人:小鼠/大鼠在人类的解剖结构、免疫系统、代谢方式上都有差异,所以动物结果未必等同于人体效果。即便如此,它们仍然是连接体外机制研究与人体真实生理的重要桥梁,用于在“整体生物体”层面验证假设。
人类肠道中微生物维生素生物合成形成的复杂网络,正在成为理解“微生物组—宿主”相互作用的核心内容之一。肠道微生物合成、共享并调节维生素可获得性的能力,并不只是简单的代谢功能,而是一条涉及生态协作、支持宿主以及进化适应的关键轴线。
在过去十年里,高通量测序、功能基因组学以及整合组学的发展显示:许多肠道细菌并不具备合成必需维生素的完整生物合成途径。相反,它们通过代谢交叉喂养形成合作网络,在群落成员之间交换代谢中间体或完整维生素。这些网络有力地说明:肠道微生物群并非一群彼此独立的个体,而更像一个协同运作的“代谢共同体”,并且与宿主健康紧密相关。
要在生态系统层面全面理解这种代谢,需要多学科的方法组合,用于绘制维生素合成的潜力,培养与实验验证能够提供因果证据与机制洞见;动物模型则用于在生理环境中验证微生物贡献。
只有整合这些方法,我们才能把握微生物维生素代谢的全部深度与细微差别。
首先,需要提升组学数据的分辨率与功能解读能力,尤其是针对非模式微生物类群以及非典型维生素变体。这要求建立更完整、更高质量的数据库,并使用更高分辨率,更大人群的组学技术来完善预测,揭示维生素代谢的调控与生态动态。
同样重要的是,将基础发现转化为可行干预手段的需求正在增长。通过使用具有明确维生素产能谱的益生菌、调整饮食以促进微生物合成,或开发个性化的微生物组营养治疗,用于营养支持、疾病预防与个体化健康管理。