-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
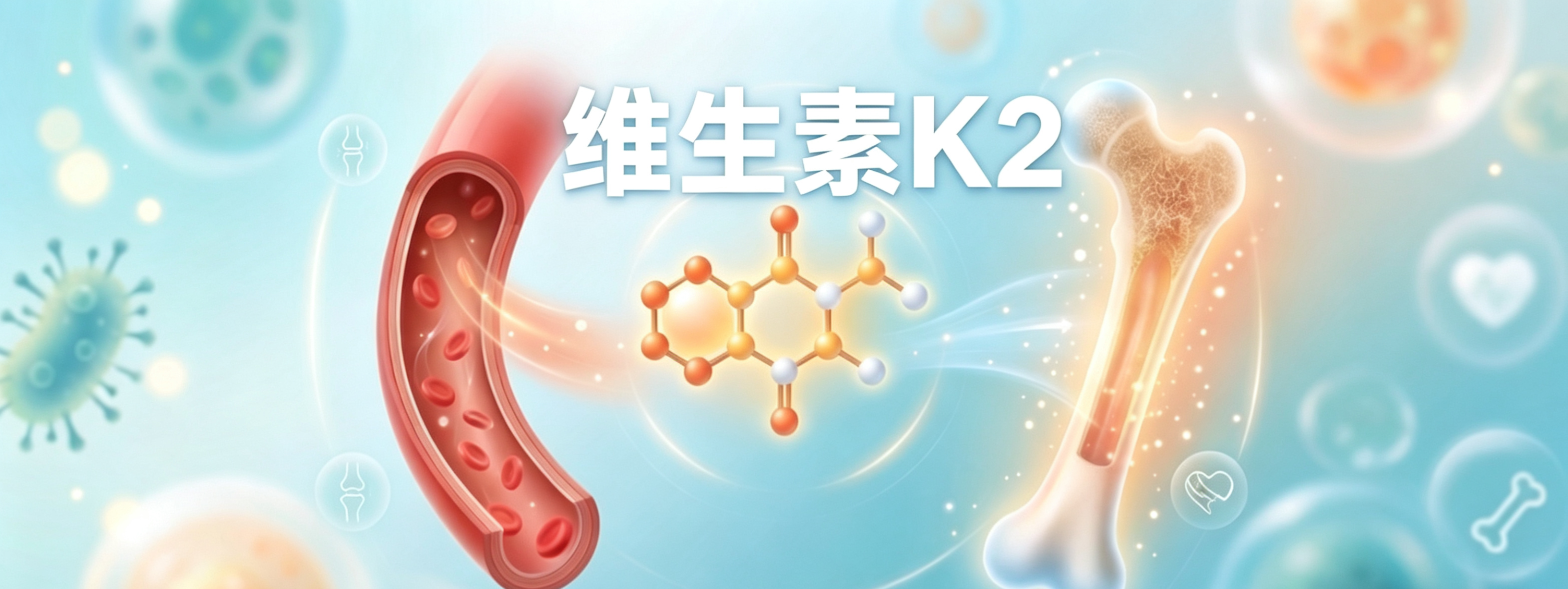
谷禾健康
维生素K是一种对身体功能至关重要的脂溶性维生素。它们被认为是人类产生多种参与凝血和钙稳态蛋白质的重要辅因子,所以又叫凝血维生素。
天然存在的维生素K主要分为两类:维生素K1(叶绿醌)和维生素K2(甲萘醌)。尽管同属维生素K,但它们之间有不少差异。
维生素K1主要存在于绿色蔬果中,K2存在于发酵食品或由肠道细菌产生。微生物K2通常半衰期更长,更容易在血清中积累,且其在肝外的血液分布范围更广,生物利用度更高。
并且维生素K2除了常规的凝血作用外,研究发现其可能还有益于心血管健康、改善骨质疏松、对肾病有帮助、或预防关节炎、改善血糖控制,并可调节免疫与神经认知,甚至具一定抗癌潜力。
维生素K2(主要为MK-7和MK-4)通过γ-羧基化(γ-carboxylation)机制激活两类关键蛋白:骨钙蛋白(Osteocalcin, OCN)和基质Gla蛋白(Matrix Gla-protein, MGP)。前者驱动钙沉积于骨基质,提高钙离子的结合力,使活化的OCN能够有效捕获血液中的钙离子并将其整合到骨矿物质中;后者则阻止钙在血管和软组织中的病理性沉积,减少血管异常钙沉积、降低心脑血管健康风险。
值得注意的是,维生素K2的内源性合成与肠道菌群状态密切相关,肠道益生菌(如乳酸菌、双歧杆菌,芽孢杆菌属等)是K2的主要内源合成者,其健康状态直接影响K2的体内浓度、肠道对外源K2的吸收利用效率,以及钙、维生素D等其他营养物质的生物利用度。抗生素使用、高脂高糖饮食、肠道炎症等因素导致的菌群失调会显著降低益生菌数量,进而导致K2合成下降,使得即使补充充足的钙和维生素D,其骨矿化效率仍然有限。
本文旨在从分子生物学、营养代谢和菌群微生态的多维度,系统阐述维生素K2的生理机制、吸收利用和代谢过程,阐明肠道菌群与K2的协同作用机制,并进一步介绍了从补给一饮食一菌群生态位层面的综合改善干预预防措施。
什么是维生素K?
维生素K(Vitamin K)又叫“凝血维生素”,因为它传统上最常与凝血相关:维生素K参与凝血相关蛋白的成熟激活(羧化),缺乏时会影响凝血功能。维生素K是一类脂溶性的小分子营养素,其最早于1929年由丹麦化学家达姆从动物肝和麻子油中发现并提取。
↘ K1、K2天然存在且为脂溶性
维生素K包括K1(叶绿醌)、K2(甲基萘醌)、K3(甲萘醌)、K4(甲萘氢醌)等几种形式,其中K1、K2是天然存在的,属于脂溶性维生素;而K3、K4是通过人工合成的。
这四种维生素K的化学性质都较稳定,能耐酸、耐热,正常烹调中只有很少损失,但对光敏感,也易被碱和紫外线分解。
虽然维生素K1和维生素K2同属于维生素K家族,并且都属于天然存在的营养成分,但它们在很多方面仍然直接存在着差异;这些不同并不只是来源层面的区别,而是会进一步体现在它们在体内的分布、可利用性以及由此带来的生理功能表现上。
注:本文这里主要介绍维生素K1和K2。
▸ 来源差异
↘ K1主要存在于绿色蔬果中
维生素K1主要存在于绿叶蔬菜、植物油和部分水果中(菠菜、卷心菜、羽衣甘蓝、牛油果),是饮食中维生素K的主要来源。
↘ K2存在于发酵食品或由肠道细菌产生
维生素K2存在于动物性和发酵食品(纳豆、奶酪、酸菜)中,或由人体肠道细菌产生。
维生素K1和K2的膳食来源
doi: 10.3390/ijms20040896.
▸ 结构差异
维生素K1含有一个由4个异戊二烯基单元构成的植酸侧链。
而维生素K2含有一个不饱和脂肪族侧链,并带有可变数量的异戊二烯基单元;丙烯基单元的数量对应不同的甲萘醌类型,包括短链的甲萘醌-4(MK-4)以及长链的MK-7、MK-8和MK-9。
其中,“n”越大代表侧链越长,从而显著影响其理化性质并进一步影响体内行为(如吸收、分布与停留时间)。
维生素K家族的化学结构
Zhang T,et al.Foods.2024
▸ 生物利用是关键差异
↘ K1主要分布在肝脏、K2分布更广泛
维生素K1和K2均为脂溶性,可与膳食脂肪和胆汁吸收。但它们在组织中的分布有所不同。肝脏、心脏和胰腺中维生素K1水平较高,但大脑、肾脏和肺部的浓度较低。
相较之下,维生素K2可存在于所有组织,但在肝脏、心脏和肺部的浓度较低。其长链衍生物可重新分布至循环系统,并用于肝外组织,如骨骼和血管。MK-4在大脑、肾脏和胰腺中的水平较高或接近K1,且其总体水平高于K1。
↘ 维生素K2半衰期更长,生物利用度更高
微生物K2(尤其是长链MK-7/MK-9)通常半衰期更长。一项比较维生素K1与MK-7的研究显示,MK-7半衰期可达68小时,而K1仅为1–2小时。
因此,维生素K2更容易在血清中积累,且其在肝外的血液分布范围更广,能更好地到达骨骼和血管等组织。相比之下,维生素K1从循环中清除更快,更易在肝脏中积累,并通过尿液和胆汁排出。
维生素K的生物利用度和生物分布
doi: 10.3390/ijms20040896.
研究发现,MK-7吸收效率和生物利用度最高。无论是摄入蔬菜来源的维生素K1,还是摄入富含K2的食物来源MK-7,二者在摄入后2小时内均可迅速吸收;但餐后血清中MK-7浓度约为K1的10倍。
同时,K1的空腹血浆浓度在个体间差异显著,且其吸收低于MK-4以及更长链的K2(如MK-8、MK-9)。长链维生素K2(尤其MK-7与MK-9)在循环中的半衰期更长,因此在循环中可被肝外组织更长时间利用。
不过,并非所有维生素K2都同样高效:MK-4的生物利用度并不一定对应给药后血清浓度升高;而MK-7给药可使血清MK-7水平在数天内持续升高,从而更好地反映并促进体内维生素K水平。
▸ 功能意义
两者均是维生素K依赖蛋白(凝血因子、骨钙素、基质Gla蛋白)γ羧化的辅因子。
K1:以肝脏凝血作用为主;
K2(尤其MK-7):更有效地激活肝外蛋白。现有证据提示其对骨骼健康和血管钙化可能有益,并被用于研究其在心血管、代谢及炎症相关疾病中的作用。
维生素K2如此重要,是因为它在人体内发挥多种关键作用,其中最为人熟知的是它作为维生素K依赖性凝血因子的激活辅因子。
近几十年的研究显示,维生素K2可能通过参与组织矿化、炎症、氧化以及年龄相关疾病过程发挥保护作用,从而为其在未来临床实践中的应用提供新的方向。
目前,维生素K2在心血管疾病、骨骼发育与骨折、慢性肾病以及某些癌症中的作用已较为明确;同时,越来越多的证据提示其在肝病、免疫功能、神经系统疾病和肥胖等方面也可能具有更广泛的作用。
对于凝血功能至关重要
维生素K是四种凝血蛋白(凝血酶原、转变加速因子、抗血友病因子和司徒因子)在肝脏内合成必不可少的物质,对γ-羧基谷氨酸的合成具有辅助作用。如果缺乏维生素K,则肝脏合成的上述四种凝血因子均为异常蛋白质分子,催化凝血作用的能力将大大下降。
维生素K促凝血和抗凝因子
Mladěnka P,et al.Nutr Rev.2022
维生素K2对心血管疾病有益
血管钙化是一个主动过程,推动心血管疾病发生,是全球主要死因之一。维生素K2依赖的蛋白质能够激活保护机制,抑制血管钙化发展。
↘ 维生素K2可以降低血管钙化风险
以MK-7形式存在的维生素K2,在健康与患病人群队列多项试验中表现出对钙化形成的长期保护作用;多项研究亦提示心血管疾病总体发生风险降低。甚至在健康人群补充后,也观察到动脉硬化的倒退与血管弹性的改善。
维生素K2的作用机制
编辑
doi: 10.1136/openhrt-2021-001715.
这些蛋白通过维生素依赖性羧化被激活。动脉粥样硬化斑块中识别的激活基质GLa蛋白可能抑制钙沉淀与软组织钙化。在一项前瞻性人群研究(LOE-A)中,纳入4807名基线无心肌梗死受试者并随访7年:维生素K2摄入量最高组相较最低组,冠心病风险显著降低。
编辑
影响因子:4.281
有趣的是,在对维生素K1和K2异构体进行全面评估的研究中,只有K2与心血管获益相关,K1则无。此外,维生素K2在心血管疾病中的作用范围较广。
目前全球仍有多项针对心血管患者补充维生素K2的大型临床研究在进行,结果将进一步完善其在该领域的证据体系。
维生素K2能改善骨质疏松症
维生素K2具有促进骨形成的作用,临床与实验研究均证实其对抗骨质疏松具有明确效果。在骨髓间充质干细胞中,维生素K2可促进骨生成的分化。
↘ 维生素K2支持骨分化,有助于抗骨质疏松
一项系统性综述(证据水平I[LOE = A])显示,维生素K2在日本患者中对预防椎骨骨折的比例为60%,对髋部骨折为77%,对非椎骨骨折为81%。其效果与传统双膦酸盐疗法相当。
另一项研究(LOE-B)纳入241名接受维生素K2(每天45 μ克)联合钙治疗的骨质疏松患者,结果表明其骨密度保持稳定;而接受钙与安慰剂治疗者的腰椎骨密度下降了2.5%,且治疗组骨折减少了65%。
与骨骼代谢相关的维生素K2作用通路
编辑
doi: 10.3390/foods10123136.
维生素K2可通过与OC无关的机制调节骨代谢。MK-7可促使成骨细胞前体细胞上调ALP、Runx2和Osterix,并促进其分化为矿化型成骨细胞;相反,在破骨细胞中维生素K2会降低其分化并抑制骨吸收。其机制与抑制细胞因子诱导的NF-κB激活有关:MK-7及较轻的MK-4可抑制NF-κB,而维生素K1未表现出类似活性。
↘ 维持骨密度,减少因骨质疏松引起的骨折
临床研究还表明,维生素K2可维持腰椎骨密度,减少年龄相关的骨质疏松性骨折;也能减少糖皮质激素诱发的骨质疏松性椎骨骨折,并在肝功能障碍相关骨质疏松中维持腰椎骨髓性低估表现;在瘫痪患者中,维生素K2则可增加脑血管疾病患者上肢的掌骨密度。
为期三年的随机对照试验(RCT,LOE = A)提示,每天补充维生素K2 180 μ克可减少腰椎和股骨颈骨密度的正常年龄相关下降,但不影响全髋关节;同时,维生素K2(MK-7)还能防止下胸椎椎高的下降。
维生素K2对肾病有益
去磷酸化-非羧化MGP(dp-ucMGP)是维生素K缺乏的公认研究标志物,最早在慢性肾病患者中被报道。dp-ucMGP水平与慢性肾病进展相关:晚期患者循环中的dp-ucMGP更高。
↘ 慢性肾病人群常缺乏维生素K
维生素K缺乏见于多种慢性肾病人群,包括尿路结石、肾结石与尿毒症。随着慢性肾病阶段加重并进入维持性血液透析,维生素K状态往往更明显受影响;动物模型与人体研究均提示,肾功能恶化时MGP可能发生过度表达,从而反映肾功能的进一步下降。
↘ 补充维生素K2有助于改善肾小球滤过等功能
已有研究表明,补充维生素K2可改善肾动脉功能,并抑制肾动脉钙化继续发展;同时也可能改善患者队列中的肾小球滤过。
因此,维生素K2在慢性肾病领域具有广阔前景,相关大规模研究正积极评估其作为治疗策略的效果。
维生素K2可能预防关节炎
↘ 缺乏K2可能导致骨关节炎
新出现的证据提示,维生素K可能在预防致残性骨关节炎方面发挥重要作用。维生素K摄入不足时,软骨与骨骼可能出现异常矿化。
动物与人体研究发现,维生素K状态与骨关节炎风险相关。以一项纳入719名日本参与者的基于人群横断面研究为例,膝关节骨关节炎风险升高与较低的VK摄入有关。在校正年龄、性别、BMI、总能量摄入、血浆维生素D以及股骨颈骨密度后,结果仍支持维生素K状态与手、膝骨关节炎呈负相关。
↘ 维生素K2有助于降低类风湿性关节炎
有证据表明,维生素K2补充剂通过降低CRP水平来减少类风湿关节炎中的炎症。维生素K2可能诱导类风湿关节炎滑膜细胞的凋亡。
在一项横断面研究中,给予100 μ克MK-7的组在3个月后病变活跃度评分显著下降,同时生化标志物(ESR、CRP和基质金属蛋白酶)有所改善。
维生素K2改善血糖控制
长期补充维生素K2可降低糖尿病发生风险。最大的一项研究纳入约38000名20至70岁男女,结果显示每日仅10 μg K2即可将糖尿病风险降低7%。
其潜在机制也逐渐被阐明:维生素K2可激活骨钙素;体外研究表明它能促进胰腺β细胞增殖,并提高胰岛素产生及CyclinD1表达。
↘ 补充维生素K2提升胰岛素敏感性并改善血糖控制
在人类研究中,饮食/补充维生素K2摄入与胰岛素浓度和敏感性显著变化相关。研究证明,高MK-4摄入量(30 mg/天)对健康年轻男性(n = 18岁,25.5–31.5岁)的胰岛素敏感性指数显著提升,相比安慰剂组(n = 15,24–31岁)在4周内显著增加。
同样,一项长期大型研究也显示,维生素K摄入量(平均摄入量:VK1 200±98微克/天,VK2 3±1/7微克)与2型糖尿病风险降低之间呈线性反相关。
维生素K补充剂可能作为一种新型辅助疗法,以改善血糖控制和生活质量。
维生素K2可能具有抗癌效果
目前已有大量研究探讨维生素K家族的潜在抗癌作用。
↘ 维生素K2有助于抑制肝细胞癌、肺癌细胞等
维生素K2可能通过多种途径发挥作用,包括蛋白激酶A、蛋白激酶C、核因子κB以及类固醇和外来受体。抑制人类肝细胞癌的生长与侵袭,并对肿瘤复发产生中度抑制。
体外研究还显示,其对肺癌细胞生长的抑制呈剂量依赖性,胰腺癌细胞亦观察到类似效应。一项纳入超过11000名患者的队列研究表明,较高的维生素K2与晚期前列腺癌显著减少相关;而维生素K1摄入增加与前列腺癌减少无关。
维生素K2调节神经认知
维生素K2可能通过多种机制支持神经功能。维生素K2的肝外活性已通过不同组织中K2循环酶的活性变化在体内得到阐明。
↘ 维生素K2对神经元有保护作用
值得注意的是,维生素K2酶在大脑中表达高度。已有文献记录K2对神经元的保护作用。MK-4提高了能量产生,并挽救了帕金森病中发现的PINK1突变。
此外,多种维生素K2类似物已被发现在神经元分化中起决定性作用。首个关于K2与神经元活动相关的流行病学研究涉及45名多发性硬化症(MS)患者和29名健康志愿者。与性别和年龄依赖的对照组相比,MS患者的K2水平显著降低。此外,K2水平与神经痉挛和视神经损伤相关。这些新兴研究表明,维生素K2在神经发育和疾病中可能扮演重要角色。
↘ 补充维生素K有助于改善认知及阿尔兹海默症
维生素K可能在阿尔茨海默病的发病机制中起作用,因为它在大脑中调节胰岛转移酶活性和生长因子/酪氨酸激酶受体活性。
有证据表明,阿尔茨海默病老年人的维生素K摄入量显著低于社区对照组。摄入维生素K可能改善健康老年人的认知功能。一项研究显示,维生素K与更好的言语情节记忆表现相关,尤其是在回忆任务中。维生素K拮抗剂的使用与认知障碍更频繁相关。
维生素K2影响免疫
近年来,体外研究提示维生素K2具有此前未知的免疫调节作用。
↘ 维生素K2可调节TNF-α、IL-1的表达
首先,MK-7可调节TNF-α、IL-1α和IL-1β的表达。研究还发现,维生素K2可抑制健康个体T细胞的增殖,而K1则无类似效果。
该结论在更多患有特应性皮炎的儿童及健康对照者的T细胞中得到验证,并得到透析患者独立研究的支持。两项研究均表明,维生素K2可减少活化T细胞数量并抑制其增殖。
综上,现有证据支持维生素K2可能作为新型免疫抑制剂发挥作用;其具体生理机制仍需进一步阐明。
维生素K2还可能影响肝脏健康与代谢
↘ 维生素K2对肝脏健康具有积极作用
维生素K在肝脏中的作用已明确:参与凝血因子生成及VKDP激活。既往研究多关注K1,但K2生物活性更高,可能与肝组织中的作用类似。近期研究还提示,维生素K2可能促进卵形细胞再生,并促进干细胞培养中肝细胞的成熟。
此外,多项试验发现MK-4对由酒精性与非酒精性肝硬化引发的肝癌具有有效的抑制作用;维生素K2补充在肝硬化治疗中也呈积极趋势。然而仍需进一步研究,以确定其效果是否具有统计学意义,并阐明维生素K2在肝病中的作用机制。
↘ 较高维生素K2与代谢综合征发生率较低相关
也有研究探讨了维生素K2与代谢综合征(MetS)之间的关联。
在一项10年随访研究中,纳入女性(n=402;49–70岁)和男性(n=400;40–80岁),结果显示高VK2摄入量(210.3 ± 127.0 μg/d)与MetS发生率降低相关(反映为三酰甘油水平下降及腰围减小);而VK1治疗组(31.1 ± 12.5 μg/d)未见类似相关性。
但一项包含5800名成年人(20–45岁)的横断面研究并随访6年发现,较高的VK1摄入量(84.8 ± 3.2 μg/天)可能有利于降低MetS患病率(以高血压下降和HDL胆固醇升高为指标)。
前面已梳理出维生素K2在多方面具有关键的生理作用,那么当机体处于维生素K2缺乏状态时,相关的代谢与调控过程可能就会失衡,从而带来一系列潜在危害。
维生素K缺乏的症状
维生素K是多种凝血相关蛋白的γ-羧化所必需的辅因子;当维生素K不足时,凝血级联中部分蛋白活性下降,可能导致凝血功能下降,增加出血倾向,表现出以下症状:
此外,维生素K缺乏还可能引发多个与维生素K相关的生理过程受到干扰:
1
骨代谢异常
维生素K2参与骨相关蛋白(如与矿化相关的蛋白)的激活,影响骨形成与骨矿化平衡。
缺乏时可能出现:
•骨密度下降;
•骨质量变差(矿化不足);
•骨折风险增加(尤其是中老年人群)。
2
血管钙化风险升高
维生素K2能促进某些“抗钙化”相关蛋白的活化;当K2不足,这些保护性作用减弱。
结果可能表现为:
•更容易发生动脉壁钙化;
•血管弹性下降;
•长期来看可能增加心血管事件风险。
3
慢性低度炎症与慢病风险
维生素K2除具有多种潜在作用外,还可能参与炎症相关的生物学反应;同时CRC相关材料提示,VK2同系物可影响炎症因子,例如在实验体系中抑制IL-8、IL-6、TNFα等促炎因子。
维生素K2缺乏的潜在危害:
•如果K2相关的抗炎/免疫调节效应不足,可能更容易出现或维持低度炎症状态;
•从而间接提高与炎症相关的慢病风险(具体到哪一种疾病,取决于人群、剂量、同系物构成、遗传与合并用药等)。
4
肠道菌群失衡
维生素K2很大一部分由肠道细菌合成,并与菌群之间存在互作关系;K2还能调节菌群动态,且与炎症及代谢相关状态密切相关。
因此维生素K2缺乏的危害可能还有:
•维生素K2缺乏可能反映或加重菌群失衡;
•菌群与K2的耦合变化可能影响代谢与炎症微环境;
•但“丰度变化→血清K2→临床终点”的因果链目前并不总是直观成立。
注:本文将在其中一个章节详细阐述肠道菌群与维生素K2之间的相互作用。
5
脂肪/代谢相关功能可能受损
维生素K2与代谢过程存在关联研究线索(例如与代谢综合征发生率、代谢指标的相关性)。
缺乏可能在部分人群中表现为代谢调控不佳,从而间接提高糖代谢异常、血脂异常与血压异常的风险(具体因果强度仍需进一步研究证实)。
6
神经系统与肌肉功能可能受影响
已有研究表明,维生素K2在神经系统相关通路中存在表达并发挥保护作用。
维生素K2缺乏可能在易感人群中与以下风险相关(多为“潜在/可能”):
•神经保护能力下降;
•神经功能维持不足;
•进一步影响认知与运动相关能力。
编者小结
维生素K2缺乏最可能的危害清单:
1.凝血风险升高:维生素K依赖凝血蛋白羧化不足。
2.骨健康下降:与骨代谢相关的维生素K依赖蛋白成熟不足,可能增加骨质疏松/骨重建失衡风险。
3.血管异常钙化倾向:影响连接组织钙化调控相关维生素K依赖蛋白。
4.炎症/慢病相关风险可能上升:维生素K2的抗炎/细胞保护方向减弱。
5.菌群生态可能进一步失衡:维生素K2—菌群互作可能形成双向反馈。
维生素K2不只参与凝血相关过程,还与骨代谢与矿化平衡、血管/组织钙化调控,以及炎症与慢性低度炎症、代谢稳态等生理活动存在联系。因此,当机体进入维生素K2缺乏状态时,相关代谢与调控环节可能失衡,进而带来潜在危害。
其风险表现包括:凝血功能受影响导致易淤青、瘀点、鼻/牙龈出血、伤口止血变慢、月经过多或异常出血等;以及骨代谢异常(骨密度下降、矿化不足、骨折风险增加)和血管钙化倾向(动脉壁钙化、血管弹性下降)等长期后果。
同时,维生素K2与炎症调节及肠道菌群互作的关联提示其缺乏可能影响炎症微环境与炎症相关慢病风险;但从“血清维生素K2变化”到“临床终点”的因果链并非在所有情境下都能直接成立。
总体来看,维生素K2缺乏的潜在影响具有多维度特征,后续可进一步结合肠道菌群与维生素K2的相互作用展开说明。
拓展:维生素K过高是否会有危害?
目前几乎没有动物或人类发生自然维生素K系统性毒性的报道。
尽管有人担心过多维生素K可能导致过度凝血,但尚未观察到这一现象,可能与γ羧化位点有限有关。相反,极高剂量维生素K可能引发低凝血酶原血症,人类病例中已有罕见记录。
★ 通常情况下不会造成危害
已有的人类数据表明,每日服用10 mg维生素K、连续1个月未见不良反应,与动物实验结果一致:在同等时间范围内,即使剂量达到2 g/kg也被认为是安全的。
一般来说,维生素K的副作用仅限于局部,例如过量服用维生素K2后出现轻微的胃肠道不适和皮疹,在停止服用后消失。
导致维生素K2缺乏,往往不是单一原因“突然缺了”,而更像是多条环节逐渐失去配合——从你吃进身体的食物,到它能不能被吸收、被运输、被利用,再到肠道菌群是否还在“按时供货”。当这些环节出现偏差,维生素K状态(尤其是与K2相关的功能供给)就可能跟不上身体需求。
此外,维生素K2并不是“一个数值搞定”。不同K2同系物在结构、生物利用度、体内分布与持续作用上存在差异;同样的摄入量,在不同人群中也可能转化为不同的功能性供给结果。再叠加个体差异(例如慢性炎症/代谢状态、可能影响维生素K相关通路的因素),就更容易出现“表面摄入并不极低,但功能层面仍接近不足”的情况。
1
摄入不足
↘ 饮食中维生素K2少、长期摄入不规律
人体自身不能在体内合成维生素K:脊椎动物依赖饮食供给;如果缺少规律摄入,体内储备会迅速被耗尽,这会构成维生素K缺乏的根本前提。
VK2主要来自特定食物谱:VK2主要来自发酵食品、部分奶酪及动物性来源;若饮食结构长期偏向“低发酵食品/低动物性、摄入以不含或少含VK2的食物为主”,更容易出现VK2摄入不足。
↘ 仅补“VK1”但不补“VK2”
部分研究/综述强调维生素K1与维生素K2在来源、吸收、靶向活性上存在差异;因此当饮食或补充只提供维生素K1而缺乏维生素K2等同系物时,不能自动等量“覆盖维生素K2的生物学需求”。
2
吸收障碍
维生素K属于脂溶性营养素,其被吸收与利用通常与消化吸收条件高度相关;若脂肪消化吸收受限,会削弱脂溶性维生素K(包含K2异构体)的吸收效率。
↘ 胆汁/脂肪吸收相关问题会降低K吸收与利用
维生素K2属于脂溶性维生素,其获得与利用往往高度依赖机体对脂溶性营养素的吸收条件;与此同时,不同形式的VK在吸收表现与体内运输机制上存在差异,而这些机制又要求机体能够完成有效吸收。因而,当这一有效吸收的过程出现不足或受损时,就会在总体层面上导致吸收环节出现障碍。
↘ 肠道转运与吸收相关蛋白/过程受影响
有文献中讨论维生素K在肠道吸收与代谢中的机制研究(如肠道受体/转运相关通路作为维生素K吸收调控因素),提示任何影响肠道吸收效率的因素,都可能把机体推向“维生素K状态下降”,从而造成VK2功能性缺乏。
3
肠道菌群失衡
↘ 肠道菌群失衡,内源性维生素K2供给下降
维生素K2可由肠道细菌生成,因此,当肠道菌群组成或功能受损时,内源性VK2供给可能下降。并且当炎症或代谢失衡导致菌群进一步失稳,也可能形成维生素K2生成/利用的不利循环。
↘ 化学用品和抗生素的过量使用
随着农业生产现代化推进,农药、化肥残留在农产品中的比例上升,且食品加工中防腐剂与人工添加剂的使用增加,可能导致肠道菌群失调,从而不利于VK2的微生物合成。
此外,抗生素可通过抑制或破坏产VK2细菌而降低VK2生成,头孢菌素类(如头孢糖素类)的影响尤为明显。质子泵抑制剂也可能影响VK2水平,其作用机制可能与对肠道菌群的影响相关。
4
药物使用
维生素K可与部分药物发生相互作用,从而影响体内维生素K2的浓度及其生物学效应。
↘ 华法林是一种维生素K拮抗剂
华法林属于香豆素类抗凝剂与维生素K拮抗剂,通过阻断肝脏及外周循环中的维生素K利用,导致继发性维生素K缺乏,进而可能对血管、骨骼、肾脏、大脑及其他系统(如炎症、免疫功能与肿瘤相关过程)产生不利影响。
VK2也可能与其他香豆素类抗凝剂(如酚丙香豆素、乙酰藻黄醇和硫氯马罗尔)发生严重相互作用。
↘ 降胆固醇和降脂药也会影响维生素K吸收
在胆固醇降药物方面,如考来替泊可通过阻断胆酸再吸收来降低胆固醇,但也会同时减少胆固醇及其他脂溶性维生素的吸收。奥利司他可降低肠道对膳食脂肪的吸收,因此可能削弱VK的吸收。
他汀类是常用的降脂药物,主要通过降低胆固醇以减少动脉粥样硬化风险,此外,他汀类药物还可能抑制VK2的合成。VK2是基质Gla蛋白(MGP)激活所需的辅因子,而MGP可帮助保护动脉免受钙化。有观点推测,由于他汀类在临床使用广泛,可能在心力衰竭与动脉粥样硬化的高发率上产生看似相互矛盾的加剧效应。
↘ 降磷药物治疗会影响维生素K的利用
在慢性肾病患者中,高磷血症较常见,因此需要使用降磷药物。研究发现,磷酸结合剂可能降低VK2的生物利用度。
体外研究显示,醋酸钙/碳酸镁、碳酸镧、碳酸钙及塞维拉麦碳酸盐均可能与VK2结合,影响其吸收,并可能削弱VK2抑制血管钙化的效果。
在一项纳入多位使用磷酸结合剂的慢性肾病患者研究中,使用塞夫拉默者的去磷化-非羧化MGP水平更高,提示维生素K水平可能降低。不过,这些相互作用的临床意义仍存在争议。
5
生理与遗传因素
↘ 体内转运与生物利用度差异
维生素K异构体的“运输路径、半衰期、组织分布、生物利用度”不同,这意味着即使总维生素K摄入看似存在,但维生素K2在体内到达靶组织并完成相关羧化修饰的效率也可能较低。
并且维生素K2包含不同链长同系物(如MK-4、MK-7等),其作用持续时间与体内行为不同。若某个群体摄入的维生素K2同系物谱不足(例如以某类同系物为主而另一类缺乏),可能出现对特定通路的激活不足,表现为功能性维生素K不足状态(即“缺乏的不是总量,而是对靶点的供能不足”)。
↘ 年龄、疾病会影响人体对维生素K的吸收
某些疾病状态会让机体更容易出现维生素K状态下降,如肝胆疾病患者以及代谢疾病老年患者的吸收能力下降可能导致维生素K2缺乏。
人体胃肠道的吸收能力随着年龄增长逐渐下降,这可能导致中老年人出现维生素K2缺乏症,因此有必要补充膳食。此外,新生儿也存在维生素K缺乏的风险,因为自身维生素K储备不足,母乳中的维生素K水平较低,未成熟的胃肠道微生物群很可能不是维生素的来源。
注:有研究发现纯母乳喂养的婴儿比配方奶喂养的婴儿更容易患维生素K维生素低水平。
总结:维生素K2缺乏的“病因框架”
1.摄入不足:人体不合成 + 储存被耗尽;维生素K2主要来自发酵/特定动物性食物。
2.吸收障碍:维生素K2为脂溶性,消化吸收条件改变会降低吸收与利用。
3.肠道菌群失衡:菌群生成维生素K2与互作关系,菌群功能下降→供给不足。
4.药物使用:部分药物是维生素K2的拮抗剂,影响其吸收利用。
5.体内利用/转运差异:维生素K1与维生素K2生物利用度、运输与靶向激活不同,可能出现“功能性缺乏”。
6.疾病/代谢需求改变:衰老与慢性疾病背景使维持足够状态更困难。
肠道微生物并不单是人体肠道的“居住者”,它们更像一个持续运转的代谢工厂:一方面,菌群能够合成并补给宿主所需的维生素家族(主要为大多数水溶性B族维生素及维生素K2);另一方面,这些微生物衍生分子又会进一步塑造肠道局部环境,影响免疫稳态与炎症生态位。
维生素K2正是这作用网络中的关键一环——它由特定肠道菌群产生,当宿主免疫状态、生命阶段与饮食结构改变时,这个供给又会被重塑。
本章将系统阐释维生素K2与肠道微生物群之间有哪些具体相互作用,以及这些作用如何为理解肠道稳态与疾病风险提供新的机制视角。
▸ 肠道菌群对维生素K2的“供给功能”
编辑
为了更好地理解肠道里维生素K2是怎么被细菌“造出来的”?我们先把生物合成这条“生产线”讲清楚。维生素K2的合成并不只依赖一种原料,许多碳源都可以进入细菌的代谢系统并最终被用来制造维生素K2。
↘ 维生素K2的生物合成过程
从结构上看,维生素K2的分子主要由两部分组成:
细菌生成VK2的过程可以概括为:先把甘油/糖类等底物加工出异戊二烯前体(用于做侧链)和DHNA骨架(用于做环),再通过特定的通路把侧链拼到骨架上,最后甲基化完成甲萘醌形成。
维生素K2的生物合成代谢途径
编辑
doi: 10.1016/j.biotechadv.2019.
↘ 能够合成维生素K2的肠道细菌
宏基因组和大规模人群研究表明,肠道菌群在结肠等部位对维生素稳态具有功能性供给作用。多数产维生素的微生物群通常只能合成单一类B或K2维生素,能够产生五种或更多维生素的细菌仅占2.7%。研究通过菌株筛查来评估内源性VK2的合成能力,并提出明确的候选菌有以下几种:
•Lactobacillus(乳杆菌属)
•Bifidobacterium(双歧杆菌属)
•Bacillus(芽孢杆菌属)
并且不同菌群可能负责不同MK形式(MK-4、MK-7、MK-9等),下面这些细菌也被发现能够产生维生素K2:
编辑
维生素合成途径的关键微生物贡献者
编辑
doi: 10.1128/msystems.00929-24.
▸ 菌群组成与菌源VK2水平的相关性
↘ 粪便中菌源维生素K2与肠道菌群组成相关
有研究测量粪便与血清中的维生素K2:发现粪便中菌源维生素K2呈现个体差异,且其水平与肠道微生态组成(如与特定菌群相关)有关;但在该研究中维生素K2未在血清中检出。
↘ 一些菌属可能与维生素K2水平共同波动
在健康成人研究中,识别了与维生素K2水平相关的菌群,提示Bacteroides、Prevotella等与维生素K2水平存在共同波动的现象,即当这些菌群相关指标发生变化时,维生素K2水平也会表现出相应的变化,从而反映出维生素K2与特定肠道菌群组成之间可能存在紧密耦合关系。
▸ 维生素K2在肠道的功能效应
↘ 可进入并作用于肠道相关细胞
编辑
结直肠癌研究显示,维生素K2(特别是长链同系物)能够进入肠上皮细胞(如Caco-2)并表现非毒性效果,提示其并非只能停留在肠腔中作为“旁观代谢物”,而可能在局部细胞界面发生作用。
↘ 长链同系物更可能与抑制促炎因子相关
研究强调:维生素K2-MK7与维生素K2-MK9可降低促炎细胞因子(如IL-8、IL-6、TNFα),从机制层面提示“菌群产出的哪类同系物”会影响“局部炎症生态位”。这把“菌群—代谢产物—炎症表型”连接起来。
维生素K2同源物对肠上皮细胞促炎IL-8浓度的影响
编辑
doi: 10.1186/s12935-023-03114-2.
炎症因子下降意味着肿瘤微环境的“驱动因子”被削弱:IL-8、IL-6与TNFα等促炎因子通常与肿瘤炎症微环境、免疫抑制/募集、上皮细胞应激等相关。维生素K2同系物能降低这些因子分泌,提示其可能改变CRC的炎症生态位,使得肿瘤发生发展的条件不那么“有利”。
▸ 在免疫稳态中的双向调控
↘ 微生物来源维生素可作为免疫稳态的协同因子
人类主要从饮食获得维生素,但肠道微生物是重要补给来源;并且肠道菌群与免疫系统互作会改变维生素生物合成,进而影响免疫反应与肿瘤免疫治疗效果。
↘ 供给不足在免疫层面可能放大风险
维生素在多种细胞过程中的重要性使得即便是轻微缺乏也可能造成健康问题。如果菌群失衡导致维生素K2供给减少或同系物谱改变,则可能通过免疫炎症通路影响肠道稳态与肿瘤微环境。
▸ 影响菌群产生维生素K2的因素
↘ 超过80岁老人与婴儿产生维生素K2能力较弱
对维生素相关基因丰度的分析显示,与宿主年龄密切相关的差异清晰可辨。具体而言,成人与80岁及以上健康老年人之间,在微生物维生素生物合成途径上存在最显著的差别。
80岁以下老年人的肠道微生物群维生素合成能力仍与年轻成人相对相近。相较之下,当年龄超过80岁时,九种维生素中有七种的生物合成能力显著下降,提示持续且渐进的功能衰退。
不同生命周期微生物群来源的维生素合成基因
编辑
doi: 10.1128/msystems.00929-24.
与健康成人相比,婴儿在九条微生物维生素生物合成途径中有六条的相关指标减少21%至49%。这一结果提示,出生后前三年微生物群的功能尚未完全成熟。
↘ 不同地域人群合成维生素K2能力不同,可能与饮食差异有关
此外,地理来源显著影响维生素生物合成通路的多样性/丰度,其中包括K相关合成潜力,从而意味着维生素K2“可由微生物提供”的程度会在不同人群中变化。
主要维生素生产细菌丰度因地理来源而异
编辑
doi: 10.1128/msystems.00929-24.
这可能与饮食差异有关,如全谷物与精制谷物饮食使粪便维生素K2水平出现差异,这提示饮食通过重塑底物/发酵环境,从而改写菌群组成,并最终影响维生素K2的输出。
不同地理区域的维生素生物合成潜力
编辑
doi: 10.1128/msystems.00929-24.
编者小结
肠道菌群和维生素K2之间的关系,像是一条在肠道里持续运转的协作链:菌群会提供维生素K2所需的合成能力与底物环境,而维生素K2不同的同系物(如长链形式)又可能反过来影响肠道局部细胞的状态与炎症水平。
在“供给”层面,能合成维生素K2的细菌并不只依赖单一原料;它们通过各自的代谢通路把底物加工成维生素K2的关键结构,再一步步完成拼接与修饰。研究提示,粪便中的菌源维生素K2水平会因人而异,并且可能与某些菌群的变化同步,说明菌群组成与维生素K2输出之间存在紧密关联。
在“效应”层面,维生素K2可以进入肠上皮相关细胞,并显示出调节作用。尤其是一些长链同系物更倾向于与促炎因子下降相关,这意味着它可能帮助“调低”肠道炎症生态位,从而为肠道稳态和结直肠癌等相关疾病微环境带来更好发展的方向。
从整体人群看,这套供给能力还会随年龄与生活方式而变化:例如80岁以上人群与婴儿的合成能力往往更弱;而不同饮食结构也会影响粪便维生素K2水平。总的来说,维生素K2像是菌群与肠道健康之间的一座“桥梁”,它的产量与类型会随菌群和环境而改变,进而影响炎症、免疫稳态以及相关疾病风险。
如何测量自身维生素K(K2)?
准确评估维生素K缺乏并不容易;即使借助最先进的检测手段,在人体生物样本中也很难准确确定其含量。维生素K可在血浆中测量,但由于其存在多种形式,且异常的脂质成分可能干扰结果,因此检测精度仍受限制。目前主要采用以下几种方法:
注:除非摄入量高,否则维生素K2在血液中通常非常低或难以检测,因此血液中的缺失并不总是意味着严重缺乏。
⑴ 直接测量—主要是色谱、质谱
色谱方法已成为直接测量维生素K的主流路线:先用液相色谱把不同K同系物分离,再用特定检测器(如荧光或质谱)定量。
在多种维生素K检测中,MS(质谱)成为首选的原因是它能更专一地区分多种同系物/同分异构体。不过局限仍存在:复杂样本前处理会影响回收率与结果一致性。
⑵ 不直接测K本身—功能性生物标志物
如果维生素K缺乏,会导致维生素K依赖的γ-羧化反应不足,于是相关蛋白以未羧化形式释放或检测到。
①未羧化的维生素K依赖蛋白
机制:维生素K缺乏 → Gla蛋白无法正常羧化 → 以未羧化形式释放。因此未羧化蛋白可作为替代定量靶点。
局限性:
-并非所有未羧化蛋白都会被分泌;
-表达升高可能压倒羧化系统;
-并非只有凝血相关Gla蛋白在健康人群都能100%羧化;
-时间因素/半衰期差异:不同凝血因子的半衰期不同,导致某些指标对轻度缺乏的敏感性与“出现时间窗口”不同。
②去羧前凝血酶—早期标志物,但敏感性较低
去羧前凝血酶(PIVKA II)是较早被用于维生素K状态评估的标志物,但敏感性较低。
③未羧化骨钙素—骨维生素K状态标志物
另一种评估骨相关维生素K状态的标志物是未羧化骨钙素。
局限性:
-总骨钙素含量不同个体差异大,所以“未羧化比例”可能更合适;
而且雌激素水平可能影响相关解读。
④未羧化-未磷酸化MGP—与血管/软组织钙化风险相关
MGP(矩阵Gla蛋白)与组织钙化调控相关,因此它更偏向“组织层面K效应”,但仍需结合检验方法与生理背景。
⑤尿液Gla残基(Gla残留浓度)— 维生素K膳食耗竭的敏感标志之一
对维生素K1膳食耗竭,尿液中Gla残留浓度是较敏感的标志物。对维生素K2缺乏的数据不足,尚未确定可靠的“界限值”。
通常需要24小时尿液采集(对依从性影响大)。
根据谷禾检测报告判断你的维生素K
谷禾肠道菌群检测通过分析你的肠道菌群组成与相关特征,可以在一定程度上提供关于你维生素K2状况的线索。
编辑
⑴ 维生素K2相关功能的线索
谷禾报告通常会给出菌群组成/优势菌属,有时也会做功能预测(或与健康维度的关联解释)。你要做的是:
找到报告里与“维生素相关合成/代谢”、或与维生素K(K2)、或与脂溶性维生素代谢/发酵相关的描述。
⑵ 可能影响维生素K2吸收的环境
维生素K2是脂溶性,吸收强依赖脂质运输与胆汁环境。所以你的菌群报告里如果能看到:
•胆汁酸相关、脂质代谢相关的风险信号(如某些与胆汁代谢不良有关的表现);
•导致吸收不佳的肠道状态(如消化吸收弱、炎症信号提示等)。
这部分能帮助解释:即便菌群“潜在供给”不错,你血/组织维生素K2也可能并不高。
⑶ 炎症/生态失衡解释“利用率”与长期效果
前面读到的维生素K2描述强调其可能涉及炎症相关过程。若谷禾报告提示菌群失衡或炎症倾向:
需要把策略从“只补维生素K2”转向“稳菌群生态 + 配合K2供给”,更符合系统性调节思路。
如何有效提高维生素K2水平?
由于维生素K2水平通常不会过高,所以我们这里只介绍在水平偏低时如何提升。具体可以从三个方向来入手,也就是围绕“补给 + 饮食 + 菌群生态位”这三方面展开:
首先在补给层面考虑提供与维生素K2相关的直接补充剂;其次在饮食层面,通过饮食结构与可发酵成分等因素提供合适的食物和发酵基础;最后在菌群生态位层面,结合菌群状态去理解并调整肠道内更利于相关功能发挥的生态环境。
维生素K的每日推荐摄入量因年龄、性别和生理状态而异。一般而言,成年男性每日需约120微克(0.12毫克),女性为90微克(0.09毫克),孕妇或哺乳期女性需75-90微克。婴幼儿及青少年需求量随年龄递增,从2微克到75微克不等。而饮食中的维生素K2摄入量平均约占维生素K总摄入量的25%。
⑴ 直接补充维生素K补充剂
常见策略是选:
•MK-7(长链,常用,半衰期通常更长);
•或在特定场景选MK-4。
具体选哪种同系物,最好与你的目标(骨/血管/炎症倾向)和复测结果对齐。
用法策略(安全与可评估):
先小幅、持续:连续用8–12周是比较常见的“观察窗口”;
复测同样指标:看浓度/同系物谱是否上升,以及你关心的功能读出是否改善。
药物干扰(非常关键):
若你在用华法林或其他维生素K拮抗药:
不要自行开始/加量补K2;
必须由医生调整剂量,并加强INR监测;
这点直接关系安全性。
⑵ 饮食补充外源性维生素K2
饮食层面要抓两个点:
1.维生素K2的直接来源;
2.脂肪同食与吸收环境(因为K2脂溶性)。
不同食物中维生素K1和维生素K2的含量
编辑
Zhang T,et al.Foods.2024
富含维生素K2的食物主要有以下几类:
①发酵食品(主要来源):
纳豆(由芽孢杆菌发酵大豆制成):含最高K2含量,尤其富含 MK-7(K1:321 ng/g;K2:10985 ng/g)。
酸菜:维生素K2的重要来源之一。
②发酵乳制品
奶酪:尤其陈年/发酵更久的类型。
酸奶:不同品牌/工艺差异很大,但可作为常规摄入。
③部分肉类和动物内脏
肉类中维生素K2在以下样本有较高含量(以微克/100克计):
鸡肉:10.1
牛肝:11.2
肉末:7.6
牛肉:1.9
猪肝:1.8
④部分鱼类
鳗鱼:63.1 微克/100克(最高);
比目鱼:5.3 微克/100克;
鲭鱼:0.6 微克/100克;
鲑鱼:0.6 微克/100克。
⑶ 调整饮食—改变菌群代谢环境和底物
饮食结构会通过改变肠道菌群在代谢过程中可获得的底物类型与供给方式,进而影响菌群代谢这些底物的效率与方向;在这样的变化之下,菌群产生维生素K2的过程也会随之受到影响。
建议的可执行食物结构:
主食优先全谷物:燕麦、糙米/胚芽米、全麦面包、藜麦等;
每天加入可发酵纤维:豆类(鹰嘴豆、扁豆、红豆等)、多种蔬菜(尤其叶菜之外),并适量藻类/菇类(按耐受选择)。
时间与频率:
建议至少持续8–12周再评估指标:菌群结构与代谢输出的改变通常不会在“几天内”立刻显现。
⑷ 菌群策略:补充益生菌
益生元(纤维)更像是“喂环境”,通常更稳妥。
也可以考虑直接补充枯草芽孢杆菌(Bacillus subtilis)、乳酸菌(Lactococcus lactis,Leuconostoc lactis,Leuconostoc mesenteroides)等能够代谢产生维生素K2的细菌。不过益生菌是否能提升K2同系物输出,取决于你的菌群背景与生态位是否匹配。
因此最好的做法仍然是:干预后复测K2或菌群结构,而不是只凭感觉。
维生素K2不仅参与传统的凝血过程,还与骨健康、心血管代谢、免疫炎症以及血糖控制等多方面息息相关。更关键的是,维生素K2在体内的“作用落点”,并非孤立存在——它离不开肠道菌群的持续参与与共同调控:一方面,肠道菌群能够合成并在一定程度上为宿主提供维生素K2相关物质;另一方面,菌群及其代谢产物会进一步塑造肠道局部环境,影响炎症水平与免疫稳态,从而左右炎症生态位与整体健康水平。
因此,在关注维生素K2时,更应该把视线延伸到肠道微生态与菌群结构。与其只纠结“要不要补维生素K2”,不如思考“为什么同样的补充效果在不同人身上可能不一样”。当菌群生态失衡、肠道环境不利时,维生素K2的摄入与利用可能难以充分发挥;而当饮食与生活方式为菌群提供合适的生存条件时,相关功能更可能在体内被更有效地启动。
维生素K2很重要,但菌群生态的健康状态同样决定了它在体内能否被更好地“用起来”。因此,除了饮食习惯以外,还要从更系统的生活方式入手——例如规律作息、适度运动、压力管理、减少过量饮酒和不必要的抗生素使用、保证充足睡眠等,帮助维持更有利的肠道环境与免疫稳态。或许正是这些协同作用,更能帮助降低疾病风险,提升整体健康水平。
注:本账号内容仅供学习和交流,不构成任何形式的医疗建议。
主要参考文献
Schwalfenberg GK. Vitamins K1 and K2: The Emerging Group of Vitamins Required for Human Health. J Nutr Metab. 2017;2017:6254836.
Halder M, Petsophonsakul P, Akbulut AC, Pavlic A, Bohan F, Anderson E, Maresz K, Kramann R, Schurgers L. Vitamin K: Double Bonds beyond Coagulation Insights into Differences between Vitamin K1 and K2 in Health and Disease. Int J Mol Sci. 2019 Feb 19;20(4):896.
Zhang T, O’Connor C, Sheridan H, Barlow JW. Vitamin K2 in Health and Disease: A Clinical Perspective. Foods. 2024 May 24;13(11):1646.
Yan Q, Zhang T, O’Connor C, Barlow JW, Walsh J, Scalabrino G, Xu F, Sheridan H. The biological responses of vitamin K2: A comprehensive review. Food Sci Nutr. 2023 Jan 6;11(4):1634-1656. doi: 10.1002/fsn3.3213. Erratum in: Food Sci Nutr. 2025 Jan 27;13(1):e70005.
Mladěnka P, Macáková K, Kujovská Krčmová L, Javorská L, Mrštná K, Carazo A, Protti M, Remião F, Nováková L; OEMONOM researchers and collaborators. Vitamin K – sources, physiological role, kinetics, deficiency, detection, therapeutic use, and toxicity. Nutr Rev. 2022 Mar 10;80(4):677-698.
Bus K, Szterk A. Relationship between Structure and Biological Activity of Various Vitamin K Forms. Foods. 2021 Dec 17;10(12):3136.
Simes DC, Viegas CSB, Araújo N, Marreiros C. Vitamin K as a Diet Supplement with Impact in Human Health: Current Evidence in Age-Related Diseases. Nutrients. 2020 Jan 3;12(1):138.
Tarracchini C, Lugli GA, Mancabelli L, van Sinderen D, Turroni F, Ventura M, Milani C. Exploring the vitamin biosynthesis landscape of the human gut microbiota. mSystems. 2024 Oct 22;9(10):e0092924.
Tarracchini C, Bottacini F, Mancabelli L, Lugli GA, Turroni F, van Sinderen D, Ventura M, Milani C. Approaches to dissect the vitamin biosynthetic network of the gut microbiota. Microbiome Res Rep. 2025 Oct 20;4(4):37.
Ren L, Peng C, Hu X, Han Y, Huang H. Microbial production of vitamin K2: current status and future prospects. Biotechnol Adv. 2020 Mar-Apr;39:107453.
Smajdor J, Jedlińska K, Porada R, Górska-Ratusznik A, Policht A, Śróttek M, Więcek G, Baś B, Strus M. The impact of gut bacteria producing long chain homologs of vitamin K2 on colorectal carcinogenesis. Cancer Cell Int. 2023 Nov 10;23(1):268.
Zhang Z, Liu L, Liu C, Sun Y, Zhang D. New aspects of microbial vitamin K2 production by expanding the product spectrum. Microb Cell Fact. 2021 Apr 13;20(1):84.
Hariri E, Kassis N, Iskandar JP, Schurgers LJ, Saad A, Abdelfattah O, Bansal A, Isogai T, Harb SC, Kapadia S. Vitamin K2-a neglected player in cardiovascular health: a narrative review. Open Heart. 2021 Nov;8(2):e001715.
Kozioł-Kozakowska A, Maresz K. The Impact of Vitamin K2 (Menaquionones) in Children’s Health and Diseases: A Review of the Literature. Children (Basel). 2022 Jan 5;9(1):78.
Gholami H, Chmiel JA, Burton JP, Maleki Vareki S. The Role of Microbiota-Derived Vitamins in Immune Homeostasis and Enhancing Cancer Immunotherapy. Cancers (Basel). 2023 Feb 18;15(4):1300.
Xue L, Singla RK, Qin Q, Ding Y, Liu L, Ding X, Qu W, Huang C, Shen Z, Shen B, Miao L. Exploring the complex relationship between vitamin K, gut microbiota, and warfarin variability in cardiac surgery patients. Int J Surg. 2023 Dec 1;109(12):3861-3871.
Karl JP, Meydani M, Barnett JB, Vanegas SM, Barger K, Fu X, Goldin B, Kane A, Rasmussen H, Vangay P, Knights D, Jonnalagadda SS, Saltzman E, Roberts SB, Meydani SN, Booth SL. Fecal concentrations of bacterially derived vitamin K forms are associated with gut microbiota composition but not plasma or fecal cytokine concentrations in healthy adults. Am J Clin Nutr. 2017 Oct;106(4):1052-1061.

谷禾健康
土壤作为地球上生物多样性最丰富的栖息地之一,不仅是植物生长的基础,也是众多微生物的”家园”。
然而,这片看似平静的地下世界,却隐藏着威胁人类健康的”隐形杀手”——人类细菌病原体。如单核细胞增生李斯特菌、铜绿假单胞菌、炭疽杆菌、沙门氏菌、大肠杆菌、结核分枝杆菌复合体和鼠疫杆菌等危险病原体,都可能在土壤中找到生存的港湾。
据统计,2019年全球约有770万例死亡与人类细菌病原体有关,这些数字背后,土壤作为病原体的自然来源和传播媒介,扮演着不容忽视的角色。
人类病原体通过多种途径进入土壤:动物和人类排泄物的渗透、农业实践中的灌溉和粪便应用、空气传播的病原体和孢子沉积等。一旦进入土壤,这些病原体可能成为常驻微生物组的一部分,并通过生食摄入、直接接触、吸入或昆虫及野生动物传播等多种途径感染人类。
本文基于最近一篇发表在《Cell Host & Microbe》上题为“Soil microbial diversity associates with lower prevalence of human bacterial pathogens across global soils”的研究,发现土壤微生物多样性与病原体流行率在全球呈负相关,并揭示病原体丰度与致病力及全球死亡率模式显著相关。在全球变化情景下,多数优势病原体比例可能上升。研究还绘制了全球土壤主要人类病原体分布图,阐明其生物地理与生态特征,为制定监测与风险管理策略、降低疫情暴发风险提供依据。
这意味着,在肠道菌群检测中发现的某些异常或致病菌,并不一定源于短期感染,而很可能是长期环境暴露累积的结果。因此,病原菌的控制不能仅依赖益生菌调节或抗生素干预,更重要的是减少其从环境进入人体的路径。
如果肠道检测报告中出现如沙门氏菌、致病性大肠杆菌、类鼻疽杆菌、克雷伯氏菌等,应重点考虑其环境来源。土壤是这些病原体的重要储存库,日常园艺、农作活动或儿童玩土等行为,都可能在无意识中增加暴露风险。特别是在潮湿或热带地区,这类病原体更易长期存活并富集。
农产品则是连接土壤与人体的重要媒介。叶菜类直接接触土壤,根茎类附着土壤较多,而水果也可能因灌溉水或土壤飞溅受到污染。如果种植过程中使用未经充分处理的粪肥,或灌溉水源受到污染,再加上洪水或暴雨等极端气候事件导致土壤扰动,都会显著增加农产品携带病原体的风险。此外,动物粪便通过土壤进入食物链,或宠物将病原体带入家庭环境,也构成重要的传播路径。
值得特别注意的是,病原体具有明显的地区性与季节性特征。不同地区的土壤病原谱差异显著,例如热带地区更易出现类鼻疽杆菌等环境致病菌,而干旱地区则更利于孢子类病原体的长期存在。同样,季节变化也会影响病原体丰度,雨季和洪水之后往往出现显著上升,高温条件促进细菌繁殖,这些都直接影响人群暴露风险。
▸ 土壤中的人类细菌病原体
一项覆盖59个国家、1602个土壤宏基因组的全球研究,为我们揭示了土壤中人类细菌病原体的分布模式。
研究中样本地点的全球分布
Xiong C,et al.Cell Host Microbe.2026
研究人员共鉴定出80个被归类为潜在土壤栖息人类病原体的细菌分类群,包括类鼻疽假单胞菌(Burkholderia pseudomallei)、结核分歧杆菌复合群(mycobacterium tuberculosis complex)、铜绿假单胞菌(Pseudomonas aeruginosa)、肠沙门氏菌(Salmonella enterica)、克雷伯氏菌(Klebsiella spp.)和志贺氏菌(Shigella spp.)等。
• 优势病原体主导模式
在这80种病原体中,25种被确定为优势病原体—它们不仅广泛存在(出现在≥80%的样本中),而且在病原体群落中表现出较高的相对丰度。
令人惊讶的是,这25种优势病原体仅占所有病原分类群的31.3%,却贡献了总病原体序列的90.8%。这种”少数主导”的模式与土壤中优势细菌和真菌的分布规律一致,反映了微生物群落结构的普遍特征。这些优势致病类群属于23种病原体,分布在19个属中,如伯克霍尔德氏菌属(Burkholderia)、分支杆菌属(Mycobacterium)、诺卡氏菌属(Nocardia)、假单胞菌属(Pseudomonas)、沙门氏菌属(Salmonella)和克雷伯氏菌属(Klebsiella)等。
▸ 高致病性优势病原体的健康威胁
在这23种显性病原体中,有11种被列为每年导致770万人死亡的33种人类细菌病原体之首,包括铜绿假单胞菌、肺炎克雷伯菌、肠道沙门氏菌、大肠杆菌、鲍曼不动杆菌、军团菌属、沙雷氏菌属和气单胞菌属等。
• 结核分枝杆菌每年导致大量感染且可在土壤中存活
其中,结核分枝杆菌尤为引人注目,其每年可导致约1060万人感染和160万人死亡。研究发现其成员能够感染各种家养和野生哺乳动物,并能在土壤中以毒力形式存活和增殖,使用变形虫和野生动物等替代宿主维持生存。
• 麻风分枝杆菌、淋病奈瑟菌也能在土壤中存在
值得注意的是,一些传统上不被认为是土壤传播的病原体(如结核分枝杆菌、麻风分枝杆菌和淋病奈瑟菌)也在土壤中被检测到。虽然它们从土壤到人类的传播途径尚不清楚,但最新研究发现蚊子可能是溃疡分枝杆菌(M. ulcerans)的新传播途径,这提示我们许多病原体的传播机制仍有待探索。
这些发现强调了重新关注病原体在土壤中的存活机制及其向人类传播途径的重要性,这对于有效监测和风险管理至关重要。
▸ 病原体的全球分布
通过对21个陆地生物群落的分析,研究发现了人类病原体在不同气候和植被类型下的分布模式。数据显示,主要人类病原体的相对丰度在热带和寒冷农田、温带湿地、温带农田以及热带和温带森林中最高。
进一步分析发现,农田、湿地和森林生态系统,特别是在热带和温带地区,表现出最高的优势病原体相对丰度。而在热带和干旱农田中,人类病原体的多样性最为丰富。
• 热带地区病原体多样性和患病率最高
分布模式与基于流行病学数据的临床研究结果一致——热带地区人类病原体的多样性和患病率最高。例如,东南亚(热带地区)报告了大多数结核病(由结核分枝杆菌引起)和麻风病(由麻风分枝杆菌引起)病例。
为探究环境因素对优势人类病原体全球分布的影响,研究人员通过随机森林和相关性分析发现:年均降水量(MAP)、年均温度(MAT)、降水季节性(PSEA)及土壤pH是最关键因素;此外,海拔、土壤质地、有机碳、土壤生物多样性(含微生物物种与功能基因多样性)及人口密度也具有重要作用。
全球土壤中主要人类细菌病原体的环境驱动因素
编辑
Xiong C,et al.Cell Host Microbe.2026
1
降雨与土壤病原体丰度正相关
• 降雨能够增加病原体在土壤中的分布
研究结果显示,降雨与23种主要病原体中的19种(约82.6%)相对丰度显著正相关。类鼻疽杆菌(B.pseudomallei)在潮湿土壤中生长最好,类鼻疽病例的增加与异常强降雨事件有关。
降雨增加可以提高土壤孔隙水含量,促进病原体的运动,增强其活性和流行性,从而促进其在土壤中的分布。
• 植物覆盖率和生产力也影响病原体丰度
植物覆盖率和生产力与病原体丰度同样呈正相关,这可能反映了植物驱动的土壤有机输入和水分稳定性增加,为某些病原体分类群的持久性提供了有利条件。
相比之下,降水季节性(23种病原体中的19种)、海拔(16种)和土壤pH值(14种)与这些分类群呈负相关。
全球不同生态系统中主要人类病原体的分布模式
编辑
Xiong C,et al.Cell Host Microbe.2026
2
土壤pH值与病原体呈负相关
土壤pH值与最主要的病原体呈负相关,表明酸性土壤往往含有更高的病原体丰度。
• 酸性土壤会破坏原有微生物结构,增加病原体增殖
这与最近的研究证据一致——土壤酸化会改变微生物群落结构,破坏关键的微生物功能,从而削弱微生物组限制病原体增殖的能力。
分析还揭示了降雨和土壤pH值之间存在显著的关系,这意味着气候变化下一些地区的降水增加可能会通过提供最佳水分条件和进一步酸化土壤来促进病原体的丰度。同样,不可持续的农业实践(如过度施肥)会加速土壤酸化,创造有利于人类病原体扩散的条件。
3
温度升高:病原体扩散的”催化剂”
温度与最主要的人类病原体(包括洋葱伯克霍尔德菌群(Burkholderia cepaciaComplex)、结核分枝杆菌复合体和炭疽杆菌)的相对丰度呈正相关,这表明全球变暖可能会进一步增加全球土壤中人类病原体的流行率。
这一发现说明气候变化可能会加剧人类致病性疾病,并表明在未来的气候情景下,降雨量和温度增加的地区可能成为人类细菌病原体的”热点”,进一步增加暴露、感染和死亡的风险。
除主要病原体外,研究还确定了其他8种病原体的环境驱动因素,包括柠檬酸杆菌属、志贺氏菌属、嗜肺军团菌、屎肠球菌、蜡样芽孢杆菌和问号钩端螺旋体等。这些病原体在降雨、温度和土壤特性方面与主要病原体具有相似的生态偏好。这些关于病原体当前分布和环境偏好的研究结果可以作为监测未来流行和爆发的基线数据,例如在筛查伯克霍尔德菌潜在爆发时,应优先考虑高降雨地区。
土壤微生物组多样性与病原体流行的关系是本研究的核心发现。研究基于宏基因组数据评估微生物物种多样性及功能多样性,并通过相关性与结构方程模型(SEM)分析其与人类病原体的关联;同时结合扩增子测序确定细菌和真菌多样性,并在全球200个土壤样本中加以验证。
▸ 微生物多样性与病原体丰度负相关
相关性分析显示,微生物物种多样性和功能多样性与大多数显性病原体的相对丰度呈显著负相关,包括类鼻疽杆菌、结核分枝杆菌复合物、炭疽杆菌、大肠杆菌和肠道沙门氏菌等。
• 土壤中细菌和真菌的多样性有助于减少病原体
在23种主要病原体中,15种和14种病原体分别与微生物种类和功能多样性呈显著负相关。研究还发现,与农业土壤相比,天然土壤中土壤微生物组多样性与主要人类病原体的相对丰度之间存在更强的负相关关系。
土壤微生物多样性决定主要人类病原体的全球分布
编辑
Xiong C,et al.Cell Host Microbe.2026
SEM分析进一步表明,土壤微生物多样性与优势人类病原体的相对丰度呈显著负相关。全球200个土壤样本亦证实:细菌和真菌多样性及细菌丰度均与主要病原体显著负相关。该结果拓展了以往土壤微观实验结论,表明提高微生物多样性可能抑制植物和人类病原体(如大肠杆菌O157:H7和肠道沙门菌)的增殖。
小编解释
较高的微生物多样性通过增加竞争者、耐压型和遗传策略类群,加剧竞争、扩大生态位占用并提升功能冗余,从而抑制病原体建立;这突显了保护土壤生物多样性对土壤、环境和人类健康的重要性。
为验证已鉴定的病原体与疾病毒力之间的关联,评估这些生物体的致病潜力,研究分析了病原体相对丰度与致病特征之间的关系。
• 土壤主要病原体丰度至少关联一种人类疾病特征
结果显示,23种主要病原体中有18种(包括类鼻疽杆菌、结核分枝杆菌、铜绿假单胞菌、鲍曼不动杆菌和大肠杆菌)以及一些其他重要病原体(如柠檬酸杆菌、嗜肺军团菌、伯氏考克斯氏体和蜡样芽孢杆菌)等,其相对丰度均与至少一种与人类疾病密切相关的关键致病特征显著正相关。
这些致病特征包括所有细菌病原体的毒力基因,根据其致病性分为不同的功能组,如毒素合酶、细胞毒性和毒力活性等。
人类细菌病原体与致病特征的关系
编辑
Xiong C,et al.Cell Host Microbe.2026
▸ 毒力因子与抗生素耐药基因
研究使用三个单独的数据库(PHI、VFDB和KEGG)进一步鉴定了与这些病原体相关的毒力因子,并使用DeepARG鉴定了抗生素耐药基因(ARG)。
• 土壤病原体相对丰度影响毒力因子与耐药性
分析表明,在全球范围内,显性病原体(类鼻疽杆菌、结核分枝杆菌、铜绿假单胞菌和鲍曼不动杆菌)的相对丰度与其相关毒力因子和抗生素耐药性之间存在很强的正相关关系。
这些发现进一步支持了识别人类病原体方法的稳健性,并证实了土壤中检测到的病原体可能会导致严重的疾病和死亡。
宏基因组揭示潜在病原体及其致病特征
为从全基因组层面验证宏基因组分箱识别显性病原体的可靠性,研究筛选出7个高质量(完整性≥90%、污染≤5%)MAG作为潜在病原体,其全球分布与定制数据库结果一致,且相对丰度与毒素合成和毒力活性等关键致病特征呈正相关。
• 宏基因组方法有助于识别土壤病原体
其中两个高质量MAG(T073.8和SRR8879131.4)与分枝杆菌的平均核苷酸同一性(ANI)达83%–84%,被认定为潜在重要病原体,并与温度和降雨量呈正相关。
这两者均携带与结核分枝杆菌复合群相关的五类致病特征(毒素合成、细胞毒性、毒力活性、器官损伤和宿主细胞死亡)、抗生素耐药基因(efpA,多重耐药)及外毒素基因(如磷脂酶C、VFC0235)。
结果表明,宏基因组学流程可用于从环境样本中识别潜在新兴病原体。
研究进一步评估了土壤人类病原体分布与全球相关死亡的关系。基于既往研究和世卫组织数据,汇总204个国家和地区的死亡率,并在国家尺度匹配到53个国家的数据。
• 土壤病原体分布与相关疾病死亡率相关
结果显示,麻风分枝杆菌(r=0.51,p=8.3×10^-5)和结核分枝杆菌复合物(r=0.47,p=0.0004)的相对丰度与病原体相关的全球死亡率(或感染病例)显著相关。
此外,细菌病原体相关死亡率与致病特征亦呈显著或接近显著相关,包括毒素合酶(r=0.45,p=0.0006)和毒力活性(r=0.25,p=0.07)。这些结果表明,土壤中人类病原体的流行可能与人类健康密切相关,提示土壤或为其天然宿主。
▸ 病原体的双向传播:土壤⇌人类
土壤与人类之间的病原体传播可能是双向的。一方面,土壤作为天然宿主,病原体可能通过各种途径感染人类;另一方面,人类病原体也可能通过空气、水道或农业实践(如施肥)等生物和非生物媒介从受感染的人群沉积或传播到自然和农业系统中。
• 土壤病原体多处于休眠状态,条件适宜时可经农产品等传播
传播到土壤中的病原体可能以低丰度或休眠孢子的形式存在,在土壤环境中找到替代宿主以生存和增殖;一旦满足传播条件,它们可能通过农产品、野生动物、病媒和城市扩张等各种传播途径重新进入人群,构成健康风险。
研究结果强调,在评估和管理与人类细菌病原体相关的风险时,需要明确考虑土壤因素。如果在地方和区域范围内得到证实,监测栖息在土壤中的人类病原体的流行情况可以为追踪全球疾病负担提供信息,简化流行病学监测,并有助于识别传染病热点。
研究局限性
疾病结果受到许多因素的影响,如人口密度、国家发展水平、社会经济状况和区域医疗质量等,在制定公共卫生政策时也应考虑这些因素。本研究所代表的国家数量有限可能会限制观察;随着数据库的改进,临床结果的预测能力也可能提高。
此外,一些病原体(如肠沙门氏菌和单核细胞增生李斯特菌)通过生食感染人类的途径已经确立,但其他病原体(如结核分枝杆菌)从土壤传播给人类的途径仍然难以捉摸。
基于环境变量(如气候和土壤因素)与人类病原体相对丰度的显著相关性,研究绘制了其在全球土壤中的当前及未来分布,模型可解释58%–88%的空间变异。
结果显示,总体显性病原体、特定病原体(如类鼻疽杆菌和结核分枝杆菌复合群)及关键致病功能(毒素合成、毒力活性和细胞毒性)在撒哈拉以南非洲、南美和东南亚等地区呈高丰度分布。
▸ 预测地图的验证及其应用价值
预测的结核病综合征全球地图显示了与世界卫生组织估计的全球结核病地图可比的模式——大多数结核病病例发生在东南亚、西太平洋和非洲。
• 病原菌分布与其致病人群分布相似
同样,预测的类鼻疽杆菌全球分布图与基于人类和动物感染病例的假鼻锤杆菌全球分布模式一致,类鼻疽在东南亚和南亚、澳大利亚北部、太平洋和印度洋岛国、撒哈拉以南非洲以及中南美洲的许多热带地区高度流行。这些结果为研究方法的稳健性提供了额外支持。
编辑
Xiong C,et al.Cell Host Microbe.2026
值得注意的是,在干旱地区(包括北非、西亚、澳大利亚北部和中部以及美国西部)发现了病原菌多药耐药诺卡菌的相对丰度最高。这与美国诺卡氏菌感染人数在西南部干燥温暖气候中最高的报告相吻合,表明不同病原体对特定气候条件的适应性。
▸ 气候变化下的病原体分布预测
在持续的气候变化下,降水模式和温度正在发生变化,这可能会对人类病原体的分布产生深远影响。
• 未来一些病原体和致病特征可能呈上升趋势
研究分析了病原体相对丰度从当前气候情景到预测的未来气候情景的潜在变化,结果表明在未来的气候情景下,一些关键病原体和致病特征的比例在全球大多数地区呈上升趋势。
例如,结核分枝杆菌复合物和致病群毒素合酶在撒哈拉以南非洲、南美洲、西欧、南亚和东南亚的某些地区呈现出明显的增长趋势。这与最近关于环境中人类病原体丰度总体增加的报告相一致。
这些地图提供了全球主要人类病原体的空间定量信息,填补了关于生活在土壤中的人类病原体的全球分布和环境偏好的关键知识空白。
然而,随着未来气候变化情景下土壤微生物组多样性的预测数据可用,以及更多宏基因组序列(特别是来自采样中代表性较低的地区/国家,如非洲国家)的积累,预测模型可以进一步改进。
总体而言,全球研究揭示了土壤中人类病原体分布的几个关键发现:主要栖息在土壤中的人类病原体的全球分布由气候条件和土壤生物多样性决定,在更潮湿的环境中观察到相对丰度更高。
土壤微生物多样性与这些病原体的流行率(即相对丰度)呈负相关,表明土壤生物多样性在最大限度地降低人类病原体爆发风险方面发挥着关键作用。
这些结果共同强调了土壤生物多样性是一个潜在的重要生态因素,它塑造了栖息在土壤中的人类细菌病原体的全球生物地理学。
▸ 研究的关键启示
这项研究确定了广泛分布在各种生态系统类型中的主要病原体,并揭示了这些病原体的相对丰度与传染病的致病特征和全球死亡率之间的正相关关系。此外,研究展示了主要病原体的环境偏好,并预测了未来全球变化可能如何影响其空间分布。这些发现对未来的研究重点和感染风险的管理具有重要意义:
1.病原体传播途径再评估:多种主要病原体(如类鼻疽杆菌、沙门氏菌和炭疽杆菌)已知可通过环境(如农产品、动物媒介和尘埃)传播,而结核分枝杆菌复合体等则传统上被认为仅经人际传播。研究表明这些病原体可在土壤中长期存活,未来应重点开展与风险管理相关的研究,如其在土壤中的存活与分布机制,以及潜在的土壤—人类传播新途径。
2.健康风险预警与管理:如果土壤中病原体的相对丰度与当地感染率之间具有稳定关系,就可以用于预测特定环境条件下的疾病风险,例如洪水或强降雨后类鼻疽的发生概率。通过持续监测土壤病原体分布与微生物多样性,并结合临床数据,有望建立更加精准和动态的疾病预警体系。
3.综合方法应对多重社会挑战:将气候、土壤与社会经济因素纳入同一分析框架,有助于提升人类病原体风险评估的系统性与前瞻性。相关研究已弥补了关键病原体在全球分布及其与生物多样性关系方面的认知空白,为预测不同气候情景下的人群健康风险提供了重要依据。
从健康管理角度看,对检测结果的理解也需要升级。肠道菌群检测不应只关注“缺乏哪些有益菌”,更应关注是否存在环境来源的病原体,以及这些菌是否反复出现。如果某类病原体持续存在,更可能提示持续暴露,而非单纯治疗不足。因此,定期监测趋势并结合生活环境变化进行分析,比单次检测更具意义。
在更宏观层面,未来公共卫生管理需要加强对土壤微生物的系统性监测,例如建立区域性的土壤微生物数据库与风险地图,对高风险区域进行预警,并在农业生产中规范粪肥处理和灌溉水管理。同时,在城市规划和灾害管理中,应将洪水后土壤病原风险纳入评估体系。
总体来看,对病原菌的控制应从“治疗导向”转向“源头管理”。环境中的土壤是一个关键但长期被忽视的源头,真正有效的健康策略应覆盖三个层面:上游减少环境暴露,中游控制食物污染,下游调节肠道生态。只有在这一整体框架下,才能更有效地降低感染风险并实现长期健康维护。
参考文献:
Xiong C, Delgado-Baquerizo M, Liang J, Wang J, Yan Z, Jensen SO, Gao M, Sáez-Sandino T, Guirado E, Muñoz-Rojas M, Román R, Maestre FT, Singh BK. Soil microbial diversity associates with lower prevalence of human bacterial pathogens across global soils. Cell Host Microbe. 2026 Mar 31:S1931-3128(26)00119-8.

谷禾健康
最近许多人开始关注一种细菌——Catabacter hongkongensis(香港过氧化氢酶杆菌),它通常以低丰度出现在人体肠道,但有一些病例报告显示其可能在黏膜屏障破损、免疫力下降等情况下带来机会性感染的风险。
该菌是少数由中国科学家2007年首次发现并命名的肠道细菌,它为何直到2007年才被发现?这反映了微生物分类研究的普遍规律:许多常见肠道细菌早已被认识,而如今更多发现的是低丰度、难以培养的罕见菌,需要借助现代分子生物学技术才能检测并分离培养。随着培养与测序技术不断进步,未来还会有更多类似的新物种被发现;香港过氧化氢酶杆菌正是其中的代表。
Catabacter hongkongensis是一种革兰阳性厌氧球杆菌。其最特别的一点是,它属于严格厌氧但又具有过氧化氢酶活性。这种特性有助于它在肠道黏膜表层的环境中存活并可能适应低氧状态,同时抵御潜在的氧化损伤,从而在特定生态位中维持生存与繁殖。
理解Catabacter hongkongensis的关键特征,有助于把握它在肠道中的“角色定位”。接下来,本文将围绕其基本属性、可能的临床相关性以及目前的研究发现,带你了解这一微生物。
Catabacter hongkongensis,中文译为香港过氧化氢酶杆菌,也可译为香港卡塔杆菌,是近年才被科学描述的一种新物种,种加词“hongkongensis”意为纪念其首次分离发现的地点——中国香港。该物种不仅代表一个新物种,也对应一个新属——Catabacter属;截至目前,该属下仅有这一种有效物种。它也是少数由中国科学家首次发现并命名的肠道细菌。
注:也有研究者认为,Catabacter是由Christensenella分化形成的多系群;基于二者的进化关系,他们提出将Catabacter hongkongensis归并为Christensenella属,并命名为 Christensenella hongkongensis。
Catabacter hongkongensis的特殊之处恰恰在于: 它是一种严格厌氧菌,却罕见地表现为过氧化氢酶阳性,这在厌氧菌中是非常少见的例外,也是它名字中“过氧化氢酶(Catabacter)”的由来。
很多人会好奇,肠道细菌有成千上万种,为什么我们要特别介绍这种罕见菌?原因是它是一种机会性致病菌,在特定条件下可导致严重败血症,病死率约50%。因此,了解它对临床诊断与治疗都十分关键。随着谷禾检测样本量的增大,更多人会在检测报告中看到它,因而需要进行科学解读。
◮ 属于厚壁菌门下梭菌目
在分类学上,香港过氧化氢酶杆菌隶属厚壁菌门(Firmicutes)、梭菌目(Clostridiales)、Catabacteraceae科,Catabacter属。该属为新建立属,分类地位相对独特,近缘属较少,提示其进化上也较为独立。
(注:分类地位目前仍有争议,详见后文说明)
尽管它最初是从血液标本中分离发现的,后续分子生态学研究表明,它实际上天然存在于人类肠道中,是肠道菌群中较罕见的成员。健康人粪便中可检测到其DNA,但丰度通常很低。因此,它本质上是肠道细菌:多数情况下长期以低丰度存在,只有在特定条件下才可能引发感染。
从进化关系看,它位于梭菌目较独立的分支,与其他已知梭菌属物种亲缘关系较远。基于这种差异,分类学家为其建立了独立的新属新种。其独特分类地位也对应其特殊生物学特征,例如过氧化氢酶阳性在厌氧菌中并不常见,而它恰是其中的例外。
Catabacter hongkongensis是一种革兰阳性厌氧过氧化氢酶阳性的球杆菌。
香港过氧化氢酶杆菌的生物学特性体现了其对肠道环境的适应,也决定了其分类鉴定上的一些特点。理解这些特性,有助于解释它为何能在肠道中定植,以及为何在特定条件下可能致病。
其一大特点是:它是严格厌氧菌,却呈过氧化氢酶阳性。严格厌氧通常对氧气十分敏感,接触氧气易死亡;而过氧化氢酶可分解氧气代谢产生的过氧化氢,使其具备一定的抗氧能力。看似矛盾,实则对应其生存环境:结肠整体厌氧,但偶有少量氧气渗入;过氧化氢酶帮助其抵御氧化损伤,是长期进化适应的结果。
1
形态
从形态上看,香港过氧化氢酶杆菌是一种球杆菌,也就是形状介于球菌和杆菌之间,球形到短杆形,大小约0.5-1.2×1.0-3.0μm。
运动性:它具有周生鞭毛,因此能够运动,这一点和许多肠道杆菌类似。
Catabacter hongkongensis
编辑
doi: 10.1128/JCM.01831-06.
孢子:它不产生芽孢,因此对高温和恶劣环境抵抗力不强,这一点和其他不产芽孢肠道细菌一致。
2
染色特性
从染色特性看,它为革兰染色阳性:细胞壁具有典型的厚肽聚糖层,染色后可保留紫色。该特征在分类鉴定中很关键,有助于与革兰阴性的肠道致病菌区分。
血液培养抹片显示革兰氏阳性杆状(红色箭头)
编辑
doi: 10.1016/j.idcr.2023.e01797.
3
生长特性
从生长特性看,它是严格厌氧菌,必须在无氧环境下生长;在有氧条件下不能存活。与此同时,它能产生过氧化氢酶,可将过氧化氢(H₂O₂)分解为水和氧气,从而消除由氧代谢产生的这种毒性物质,因此对氧气的耐受性相对其他严格厌氧菌更强。也正因如此,它能够在氧分压稍高的环境中生存,例如肠道黏膜表面附近——这里会有少量氧气从组织扩散进入。
4
代谢能力
从生化反应看,香港过氧化氢酶杆菌具有几个关键特征:其过氧化氢酶阳性是最重要的生化标志,也正是中文“过氧化氢酶杆菌”的命名来源。
其吲哚试验阴性、硝酸盐还原试验阴性;可发酵阿拉伯糖、葡萄糖、甘露糖和木糖产生酸,但不产气。
5
环境适应力
其最佳生长温度为37°C,与人体环境相近。由于不产芽孢,它对热、干燥和消毒剂的耐受力较弱;因此与其他不产芽孢的肠道细菌一样,充分加热即可有效杀灭它。
总结来说,香港过氧化氢酶杆菌的基本生物学特性可概括为:革兰阳性、严格厌氧、过氧化氢酶阳性、能运动、不产芽孢的球杆菌。
这些特征反映了它对人类肠道环境的长期适应,其中过氧化氢酶阳性最为独特,帮助其抵抗氧化损伤并适应肠道条件。了解这些基础后,下一章将重点讨论它的代谢特征,并进一步说明厌氧菌为何会进化出过氧化氢酶活性。
过氧化氢酶活性在兼性厌氧菌和好氧菌中较常见,因为它们需分解代谢过程中产生的过氧化氢毒性。但是香港过氧化氢酶杆菌是严格厌氧菌,为什么还需要过氧化氢酶呢?这个问题其实反映了它对肠道特殊环境的进化适应:结肠虽以厌氧为主,仍可能有少量氧与过氧化物形成,而过氧化氢酶对其生存具有关键意义。
结肠虽然以厌氧环境为主,但仍存在一定量的活性氧
要理解其作用,首先需弄清肠道过氧化氢的来源。过氧化氢主要来自:宿主细胞呼吸产生的活性氧、宿主免疫细胞“呼吸暴发”产生的过氧化氢,以及肠道细菌代谢产生的部分活性氧。
尽管结肠以厌氧环境为主,氧气仍可从黏膜组织微量渗入,同时免疫细胞活化也会带来活性氧。因此,即便是严格厌氧菌,也必须具备一定的抗氧化损伤能力,而过氧化氢酶正是用来分解过氧化氢、降低其毒性。
为减轻氧化损伤,适应进化出过氧化氢酶
香港过氧化氢酶杆菌的过氧化氢酶阳性,是进化适应的结果。它主要定植在肠道黏膜表面附近,这个位置离组织近,氧气渗入比肠腔更多,过氧化氢浓度也更高,因此需要过氧化氢酶来分解过氧化氢,保护自己免受氧化损伤。
如果没有过氧化氢酶,它就无法在黏膜表面生存,只能在肠腔深处生存。因此,过氧化氢酶活性实际上扩展了它的生态位,让它能够定植在黏膜表面,这就是为什么严格厌氧菌还需要过氧化氢酶的原因。
能够代谢多种碳水化合物并产生有机酸
它是严格厌氧菌,能量代谢主要通过发酵途径产生ATP,不能进行有氧呼吸。
从碳水化合物代谢看,香港过氧化氢酶杆菌能发酵多种碳水化合物并产酸,如阿拉伯糖、葡萄糖、甘露糖和木糖等,表明其可利用多样化的膳食碳水化合物获取能量、适应肠道环境。其发酵主要生成有机酸,且不产生大量气体。
过氧化氢酶还有助于分解毒性物质,增强抵抗力
过氧化氢酶活性还可能带来另一重益处:当宿主免疫细胞产生活性氧以杀菌时,过氧化氢酶可帮助细菌分解这些毒性物质,从而增强其对免疫攻击的抵抗力。由此可能提升其在炎症环境中的生存与侵入能力,尤其当肠道黏膜受炎症损伤时更为有利。
从生态代谢理论看,这一代谢特征对应其生态位:低丰度、黏膜表面定植,并需抵抗微量氧与活性氧。过氧化氢酶正好满足这些需求,因此被自然选择保留下来。
本节小结
香港过氧化氢酶杆菌(C.hongkongensis)虽然是严格厌氧菌,但它需要过氧化氢酶来抵抗黏膜表面微量氧气和活性氧造成的氧化损伤,这是对它特殊生态位的进化适应。这种适应让它能够定植在其他厌氧菌不能很好生存的位置,扩大了自己的生态位。
在理解了这些代谢特征之后,我们接下来讨论它在肠道中的分布情况,并进一步分析其在不同肠道环境中出现和维持的定植规律。
可定植于人体肠道,丰度一般较低
香港过氧化氢酶杆菌天然存在于人体肠道,是肠道菌群的成员之一,但因其毕竟罕见、丰度通常很低,长期未被科学家发现。它主要定植于结肠,尤其是黏膜表层,这与其过氧化氢酶阳性的特性相符:黏膜表层氧分压略高,需要过氧化氢酶来抵御氧化损伤。
分子生态学研究表明,健康人肠道中也可检测到该菌DNA,证实其为天然共生菌,而非外源性致病菌。由于丰度较低,通常占总菌群不到0.1%。它属于条件致病菌:在正常情况下与宿主共处,只有在特定条件下才可能引发感染。
哪些因素会影响它的丰度呢?
研究发现,剖腹产分娩的儿童肠道中香港过氧化氢酶杆菌丰度显著高于顺产儿童,具体原因还不清楚,可能和分娩过程菌群定植顺序改变有关。
编辑
影响因子:4.123
另外,帕金森病患者肠道中它的丰度也显著升高,但是否和帕金森病发病有关目前还不清楚,只是相关性,不能确定因果关系。
定植过程及影响因素
它的定植过程与大多数肠道细菌相似:经口摄入后,随食物从胃进入结肠,并在结肠黏膜表面找到合适的生态位而定植。由于其过氧化氢酶阳性,能够耐受黏膜表层的低氧环境,因此更倾向于在该位置定植。
影响其定植的因素主要包括:
1.正常菌群的定植抵抗——若菌群完整,它难以过度生长;
2.饮食因素——其可利用多种碳水化合物,膳食纤维丰富的饮食可能有助于其以低丰度持续存在;
3.以及黏膜屏障完整性——屏障完整时,它难以侵入深层组织并进入血流。
它的携带状态可以持续很长时间,健康人可以长期低丰度携带不发病,这就是它和宿主共生的状态。只有当黏膜屏障破坏,比如患有结肠癌,肿瘤表面溃疡破裂,细菌就可以侵入血流引起感染。
目前对香港过氧化氢酶杆菌(C.hongkongensis)的生理功能研究还很少,绝大多数健康人都低丰度携带,没有明显症状,因此很难研究它对宿主的生理贡献。基于目前的证据,我们可以给出以下结论:
C.hongkongensis一般处于低丰度,对宿主没有影响
首先,香港过氧化氢酶杆菌是人类肠道正常菌群中的罕见成员。以低丰度方式长期携带时,对宿主通常没有明显危害,也缺乏证据表明其具有特殊的有益作用。总体而言,它属于共生菌,在肠道生态系统中占据一个较小的生态位,对整体功能影响有限。
那么,它为何能在肠道中长期存在?关键在于它占据了其他细菌难以充分利用的生态位,同时对宿主缺乏明显危害,因此自然选择难以将其淘汰。由此,它得以持续存在于人类肠道,这也是许多低丰度肠道细菌普遍的生存方式。
可能代谢过氧化氢并影响肠道微生态
它是否可能对宿主有潜在益处?例如,由于它具有过氧化氢酶,可能分解过氧化氢、降低肠道局部活性氧,从而减轻氧化应激——从机制上看是可能的,但目前缺乏研究证据。虽然它的确可能降低局部氧化压力,但由于丰度很低,其整体作用可忽略不计。
从另一个角度,作为肠道菌群成员,它可能通过增加菌群多样性间接促进生态系统稳定。菌群多样性更高通常有利于维持生态稳定,因此低丰度的多种稀有菌可能共同带来这种间接效应。不过这种贡献并非由单一物种主导,香港过氧化氢酶杆菌的单独作用仍较小。
因其丰度低,对其他菌群的影响有限
香港过氧化氢酶杆菌在健康人肠道中丰度很低,因此与其他细菌的相互作用有限,难以对整体菌群结构造成明显影响。它主要占据结肠黏膜表层这一特定生态位,其他细菌难以充分利用该位置,从而缺乏剧烈的营养竞争,能够与周围菌群和平共处。
注:这一生态位的氧分压相对较高,需要过氧化氢酶才能更好生存。多数严格厌氧菌缺乏过氧化氢酶,难以在此环境下良好生长,因此香港过氧化氢酶杆菌能够在该处建立生态位分化。
只有在菌群失调、其丰度异常升高时,才可能打破平衡并产生影响,但这种情形较少见。
此外,它为过氧化氢酶阳性,可分解过氧化氢生成氧气和水,可能轻微改变局部微环境,从而对周围细菌产生一定作用。但由于其数量较少,整体影响通常很小,一般不会造成实质性改变。
正常菌群可能对其有一定抑制作用
正常菌群会抑制它吗?
答案是会。香港过氧化氢酶杆菌同样受到正常菌群的“定植抵抗”影响:当菌群完整时,它难以过度生长;只有在抗生素等因素破坏正常菌群后,才可能增加其丰度。因此,维持正常菌群完整性有助于维持其低丰度携带状态,避免过度繁殖。
总的来说,香港过氧化氢酶杆菌通过生态位分化实现与其他肠道细菌的和平共处:它占据黏膜表层这一相对竞争较少的区域,能相对稳定地以低丰度存在,从而对整体菌群功能影响有限。这种共存模式构成了它与宿主共生的基础。
香港过氧化氢酶杆菌可归类为机会性致病菌:多数情况下不致病,只有在特定条件下才可能引发感染。主要诱因包括肠道黏膜屏障破坏和宿主免疫力低下。截至目前,全球报道的确诊感染病例绝大多数患者均有明确基础疾病,提示其确属机会性感染。
我们来看看已经报道的感染病例都有哪些基础疾病和临床表现:
1
胃肠道恶性肿瘤
这是最常见的基础疾病,约占报道病例的一半。胃肠道恶性肿瘤(尤其结肠癌)常伴随肿瘤坏死溃疡,导致黏膜屏障破坏。
编辑
从溃疡处进入血液引起菌血症和败血症
香港过氧化氢酶杆菌便可能从溃疡处进入血流,引发菌血症与败血症。已报道的病例中,3位结肠癌患者感染后预后较差,其中2位死亡,提示恶性肿瘤患者感染后的病死率较高。
2
急性阑尾炎
香港过氧化氢酶杆菌可以引起急性坏疽性阑尾炎,这种情况下,细菌从破损的阑尾黏膜侵入血流,引起菌血症。
两位健康年轻人得过急性阑尾炎合并香港过氧化氢酶杆菌感染,手术切除阑尾抗感染治疗后痊愈,说明如果是单纯阑尾炎没有其他基础疾病,预后较好。
3
急性胆囊炎
也有报道显示,急性结石性胆囊炎可导致香港过氧化氢酶杆菌感染:胆囊黏膜受损后细菌进入血流。经抗感染治疗并在手术切除胆囊后痊愈。
感染的临床症状
因此,感染途径很明确:先发生肠道黏膜屏障破损,细菌发生移位进入血流,继而引起菌血症或败血症。绝大多数为内源性感染,即由患者自身肠道携带的细菌侵入血流,而非外源性传播。
临床上常见表现包括发热、寒战以及全身毒血症状;严重时可出现感染性休克。若不及时治疗,病死率较高,总体约50%,主要与患者多合并严重基础疾病(如晚期恶性肿瘤)有关。
总结来说,香港过氧化氢酶杆菌致病需要两个条件:一是肠道黏膜屏障破坏,让细菌能够进入血流;二是宿主免疫力低下,不能清除侵入的细菌。它几乎只在有基础疾病的情况下致病,健康人很少致病。
当你在谷禾肠道微生物检测报告中看到“香港过氧化氢酶杆菌(C.hongkongensis)”,很多人会担心:它是不是致病菌?我是否需要治疗?
单次检出不等于就已经生病。是否需要处理要综合判断其丰度以及是否有相关临床症状。在大多数情况下,如果丰度较低,通常不需要特殊处理。
编辑
我们一般分几种常见情况解读:
情况一
检出极低丰度,没有任何症状
这种情况是最常见的,说明你就是正常低丰度携带者,这是正常情况,不需要任何特殊处理。它本来就是人类肠道罕见共生菌,低丰度携带不发病,放心好了。
情况二
检出低到中等丰度,没有症状
这种情况通常不需要特殊治疗,建议以观察为主并定期复查。保持健康饮食、维持正常菌群完整性,有助于防止其过度生长。
情况三
检出中等至高丰度,没有症状
这种情况需要结合你的基础疾病判断:若你有胃肠道肿瘤、炎症性肠病等,提示黏膜屏障可能受损,应由医生评估是否存在细菌移位风险;若你是健康人、完全无症状,通常可定期复查并观察丰度变化,无需立即治疗。
情况四
检出丰度升高,同时有发热、腹痛等症状
这种情况需要高度警惕,应尽快就医并进一步检查,排除细菌移位及菌血症。若你本身有胃肠道肿瘤、阑尾炎、胆囊炎等基础疾病,更应及时评估。
5
通用预防建议(所有人适用)
预防其实主要就是维持肠道健康:
1.均衡饮食:足够膳食纤维,维持正常菌群平衡;
2.不滥用抗生素:保护你的正常菌群,就是保护你自己;
3.及时治疗胃肠道疾病:保持黏膜屏障完整,减少细菌移位机会;
4.定期体检:有基础疾病定期检查,早期发现问题早期处理。
几个重要原则需要强调:
第一,大多数健康人可低丰度携带、无需治疗。作为机会致病菌,它只有在特定条件下才可能致病,检出并不等同于必须杀菌。
第二,若出现致病,通常属于内源性感染,关键在于黏膜屏障破损后细菌移位进入血流。因此,基础疾病是最重要的风险因素,健康人总体上很少发病。
第三,现有研究仍有限:丰度升高与帕金森病等慢性病目前多为相关,并未证实因果关系。不应仅凭检出就判断其与你的疾病有关。
解读原则总结
低丰度且无症状,正常观察;高丰度且有症状,立即就医;有基础疾病则加强监测。多数情况下,检出本身不会影响健康,无需过度担心。
总的来说,香港过氧化氢酶杆菌(Catabacter hongkongensis)是一种较罕见的机会性致病菌,大多数人都是正常低丰度携带,不会致病,不需要过度担心。
作为一个近年才发现的新物种,它的故事告诉我们,人类肠道微生物组还有很多未知的物种等待发现,即使是低丰度物种,在特定条件下也可能给健康带来影响。随着测序技术进步,越来越多这样的罕见物种会被发现,我们对肠道微生物组的认识会越来越完整。
从目前研究来看,它是人类肠道正常的低丰度共生菌,只有在黏膜屏障破损和免疫力低下情况下才会引起机会性感染,多数感染发生在有严重基础疾病的患者,健康人很少发病。它最特别的生物学特性就是严格厌氧却具有过氧化氢酶活性,这是对黏膜表层低氧环境的适应进化,帮助它抵抗氧化损伤,这个特性让它占据了其他细菌不能很好利用的生态位。
未来研究需要进一步回答几个问题:
香港过氧化氢酶杆菌(Catabacter hongkongensis)的发现也说明,人类对肠道微生物的认知仍在深化。未来仍会有更多新发现,帮助我们更好理解肠道微生态,从而更好地维护健康。
参考文献
Liu X, Sutter JL, de la Cuesta-Zuluaga J, Waters JL, Youngblut ND, Ley RE. Reclassification of Catabacter hongkongensis as Christensenella hongkongensis comb. nov. based on whole genome analysis. Int J Syst Evol Microbiol. 2021 Apr;71(4):004774.
Lau SK, Fan RY, Lo HW, Ng RH, Wong SS, Li IW, Wu AK, Ng KH, Tseung S, Lee RA, Fung KS, Que TL, Yuen KY, Woo PC. High mortality associated with Catabacter hongkongensis bacteremia. J Clin Microbiol. 2012 Jul;50(7):2239-43.
Yang Y, Chen J, Gao H, Cui M, Zhu M, Xiang X, Wang Q. Characterization of the gut microbiota and fecal and blood metabolomes under various factors in urban children from Northwest China. Front Cell Infect Microbiol. 2024 Mar 22;14:1374544.
Itoh N, Akazawa N, Ishibana Y, Murakami H. Peritonitis with bacteremia due to Christensenella hongkongensis identified via ribosomal RNA sequencing in a Japanese patient with advanced colorectal adenocarcinoma: A case report. IDCases. 2023 May 9;32:e01797.
Lau SK, McNabb A, Woo GK, Hoang L, Fung AM, Chung LM, Woo PC, Yuen KY. Catabacter hongkongensis gen. nov., sp. nov., isolated from blood cultures of patients from Hong Kong and Canada. J Clin Microbiol. 2007 Feb;45(2):395-401.
Petrov VA, Saltykova IV, Zhukova IA, Alifirova VM, Zhukova NG, Dorofeeva YB, Tyakht AV, Kovarsky BA, Alekseev DG, Kostryukova ES, Mironova YS, Izhboldina OP, Nikitina MA, Perevozchikova TV, Fait EA, Babenko VV, Vakhitova MT, Govorun VM, Sazonov AE. Analysis of Gut Microbiota in Patients with Parkinson’s Disease. Bull Exp Biol Med. 2017 Apr;162(6):734-737.
Cabrol M, Rammaert B, Plouzeau C, Burucoa C, Pichon M. Phylogeny of Catabacter hongkongensis Strains Responsible for Bacteremia Is Not Associated with Clinical Outcomes or Therapeutic Efficacy. Diseases. 2021 Mar 25;9(2):24.
Mandin V, Corvec S, Chéné AL, Guillouzouic A, Dirou S, Bennouna J, Bémer P, Pons-Tostivint E. A fatal case associated with Catabacter hongkongensis bacteremia in lung cancer patient: A case report. IDCases. 2022 Jun 14;29:e01535.
Kaden R, Thelander M, Engstrand L, Herrmann B. First case of human bacteraemia by Catabacter hongkongensis in Scandinavia. New Microbes New Infect. 2016 Oct 4;15:6-8.
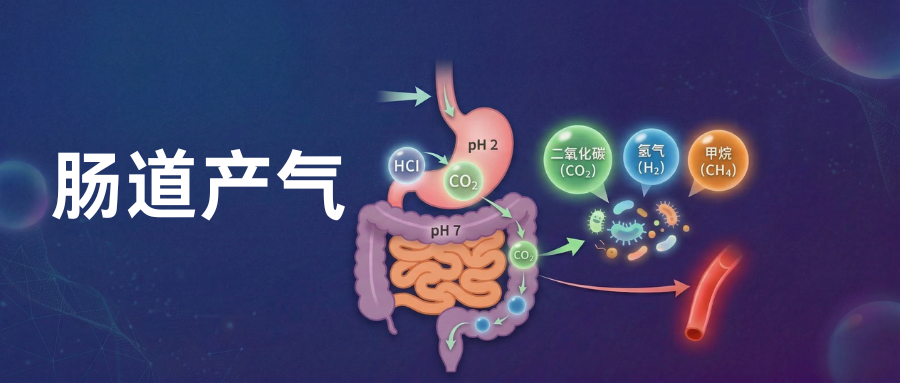
谷禾健康
在当代社会的饮食结构和生活节奏下,腹胀、排气增多以及进食后不适,已经成为越来越高频的人群亚健康反馈。根据以往的经验很多人会把这些情况简单理解为饮食不当、消化不好或肠胃比较敏感,但这样的解释往往只停留在表面。
如今,随着大量的临床研究推动,这类现象其实与肠道内一个重要过程有关——肠道产气。我们吃进去入的食物并不会被完全被身体消化吸收,其中一部分会进入大肠,被肠道菌群进一步分解利用。在这个过程中,微生物会通过发酵产生多种气体,比如氢气、甲烷、二氧化碳等。
在正常情况下,这些气体的产生与排出是一个相对平衡的过程,不会引起明显不适。但当饮食结构、进食速度、压力、疾病状态或肠道菌群组成发生变化时,气体产生和排出平衡就会改变,从而引发腹胀、频繁排气甚至餐后不适等症状。
例如,很多被视为健康的食物,本身恰恰富含可发酵底物,包括豆类中的棉子糖家族低聚糖(RFOs),蔬果和谷物中的果聚糖,以及部分人群无法在小肠充分消化的乳糖。这意味着,这些底物进入不同个体的肠道后,会被不同的菌群结构和代谢通路处理。
因此,从肠道微生态的视角来看,这些日常不适并不只是消化差或吃饭问题,而是身体内部微生物代谢活动的一种外在表现。理解这一点,有助于我们更理性地看待症状,也为通过饮食调整和生活方式干预来改善肠道状态提供了更清晰的方向。
我们先来了解这些常见的核心气体,这也是后续做症状分层和干预判断的基础。
氢气:结肠发酵最典型的气体信号
在肠道主要气体中,氢气(H₂)几乎可以看作结肠发酵最具代表性的代谢信号之一。H₂ 是由结肠细菌对不可消化底物进行代谢后产生的主要气体,而且它仅来源于细菌发酵,并不是宿主自身代谢直接生成的产物。也正因为这一点,H₂ 在理解肠道发酵活跃度时具有很高的指示意义。
在一定条件下,肠道在 24 小时内可产生高达 1 L 的 H₂。到了这个水平,H₂ 已经不只是一个代谢背景值,还可能和腹胀、腹痛、排气增多等症状联系起来。也就是说,H₂ 一方面是正常结肠发酵的组成部分,另一方面也可能成为症状形成的重要推动因素。
哪些菌群参与氢气生成?
从菌群来源看,和 H₂ 生成关系最密切的,主要包括拟杆菌属(Bacteroides)、瘤胃球菌属(Ruminococcus) 、 Roseburia。
此外,Anaerostipes caccae、Clostridium、Eubacterium rectale、Enterococcus、Victivallis vadensis等菌群,也都参与了 H₂ 的生成过程。
人类肠道菌群中存在的主要产气菌
doi.org/10.1016/j.jff.2022.105367
这些菌群有一个共同点:它们能够利用进入结肠、但没有在上消化道被充分处理的底物,尤其是各类可发酵碳水,在厌氧环境中进行分解和发酵。这个过程中,除了生成短链脂肪酸等代谢产物,也会释放出 H₂ 和 CO₂。换句话说,H₂ 的出现,本身就说明结肠内的代谢系统正在运转。
氢气是怎么产生的?
最常见的路径是经典的糖酵解过程(Embden–Meyerhof–Parnas pathway)。肠道细菌将碳水化合物分解为丙酮酸后,会产生大量还原型辅因子,而这些还原力需要被释放和再平衡,这时微生物中的氢化酶(hydrogenases)就会参与进来,推动 H₂ 的形成。
H2还可通过以下机制生成:
1)丙酮酸裂解生成甲酸,随后经甲酸裂解酶代谢;
2)丙酮酸在丙酮酸:铁氧还蛋白氧化还原酶和氢化酶作用下生成H2
所以,H₂ 的生成本质上反映的是肠道厌氧发酵系统在处理底物和转移电子,它是代谢网络维持运行的重要一环。
细菌代谢产生H2、H2S、CH4和CO2的生化途径
编辑
doi.org/10.1016/j.jff.2022.105367
为什么氢气不会一直在肠道里累积?
虽然 H₂ 是主要产气成分之一,但它不会在肠道内无限累积。原因在于,过高的 H₂ 分压会抑制进一步代谢,从热力学上影响细菌继续代谢底物。因此,肠道必须持续把过量 H₂ 移走,整个代谢系统才能维持运行。
肠道中约三分之一的H2被结肠中的其他微生物利用,其余则以肛门排气或呼气形式排出。而承担“耗氢”任务的主要有三类微生物:
这意味着,H₂ 在肠道里更像是一个代谢枢纽。它一头连接上游代谢,一头连接下游的甲烷通路、硫代谢通路和乙酸生成通路。
利用气体的肠道微生物
编辑
doi.org/10.1016/j.jff.2022.105367
氢气升高,通常说明什么?
从功能上看,H₂ 最值得关注的地方,在于它对“代谢负荷”的提示作用。
H₂ 并不只是“会不会胀气”的问题,它更像是一个代谢系统是否处于高负荷状态的功能读数。
也正因为 H₂ 与细菌代谢过程关系非常直接,呼气氢已被广泛用作评估糖类吸收不良和小肠细菌过度生长(SIBO)的重要指标。原因在于,H₂ 直接反映了肠道细菌对未被吸收底物的代谢活动。
二氧化碳:始终处在代谢主通路中
二氧化碳也是结肠代谢的主要气体之一
在讨论肠道产气时,二氧化碳(CO₂)往往没有氢气和甲烷那么受关注,但从代谢过程看,它一直是结肠细菌在结肠中对膳食底物进行代谢代谢时产生的主要气体之一。
和 H₂ 一样,CO₂ 也是菌群处理未被上消化道充分吸收底物后的结果,因此它本身就处在肠道代谢主通路中。
哪些菌群参与了二氧化碳生成?
参与膳食碳水发酵并产生 CO₂ 的菌群,主要仍然集中在厚壁菌门和拟杆菌门两大类群中。
也就是说,CO₂ 的产生与结肠中最主要的发酵菌群密切相关,和 H₂ 的菌群背景有很大重叠。
二氧化碳是怎么产生的?
CO₂ 的产生与结肠发酵中最基础的碳流动和能量代谢过程直接相关。只要结肠内的发酵在持续,CO₂ 就会持续生成。
二氧化碳会如何被处理?
和其他肠道气体相比,CO₂ 的一个重要特点是:相当一部分会被肠黏膜吸收。研究指出,大多数 CO₂ 会经结肠黏膜被动吸收,进入循环后再通过呼吸排出体外。
除此之外,未被吸收的 CO₂ 也可以随排气排出,或者被肠道内其他微生物继续利用。
在后续代谢中,CO₂ 还会继续参与两类重要通路:
所以,CO₂ 在肠道里既是代谢产物,也是后续代谢网络中的重要底物。
二氧化碳升高,通常说明什么?
从肠道代谢角度看,CO₂ 的持续生成说明结肠代谢通路处于活跃状态,尤其与碳水底物的厌氧分解密切相关。它本身虽然不如 H₂ 和 CH₄ 那样常作为单独指标被强调,但在代谢网络中,CO₂ 一直是一个基础节点:
硫化氢:量不大,但值得关注
和 H₂、CO₂、CH₄ 相比,硫化氢(H₂S)在肠道总气体体积中的占比并不高,但它的生物学意义很强。原因在于,H₂S 不只是一个代谢产物,它同时还是一种具有信号作用和细胞效应的含硫小分子。我们来看看,H₂S 来自哪条通路、处在什么部位、达到什么浓度,以及宿主能否把它及时解毒和清除。
人类大肠和粪便中的 H₂S 浓度大约在 0.17–0.38 mmol/L。
哪些菌群参与了硫化氢生成?
肠道内负责生成 H₂S 的核心菌群是硫酸盐还原菌(sulfate-reducing bacteria, SRB)。
其中最常见、最典型的代表是Desulfovibrio piger。此外,Desulfovibrio fairfieldensis、Desulfovibrio desulfuricans、Desulfobulbus、Desulfomicrobium、Desulfomonas 等,也都属于与 H₂S 生成相关的菌群。
SRB 在结肠中的数量范围大约可达 10³–10¹¹ CFU/g 。其中 Desulfovibrio 属是人类结肠中最主要的一类 SRB。
不过,现有研究也提示,肠道 H₂S 的来源并不只有 SRB。Fusobacterium 这类菌同样能够参与 H₂S 生成。
此外,卡拉胶(carrageenan) 等硫酸化多糖也可以提供相关底物。研究还指出,粪便 H₂S 升高与红肉摄入增加存在相关性。
硫化氢是怎么产生的?
最经典的一条路径,是 SRB 在厌氧环境下利用 H₂ 作为电子供体、利用硫酸盐作为末端电子受体,通过异化硫酸盐还原作用,最终生成 H₂S。
换句话说,这条路径本质上是一个典型的耗氢分流过程:上游发酵菌持续产出 H₂,下游 SRB 再把这部分 H₂ 转入硫代谢通路。
另一条重要路径,与含硫氨基酸代谢有关。像 Fusobacterium nucleatum 这类菌通过半胱氨酸脱硫酶将L-半胱氨酸等含硫氨基酸转化为 H₂S、丙酮酸、氨。
也就说,H₂S 的生成至少同时连接了两条线索:
H₂S 的问题,关键在浓度和部位
H₂S 的作用具有明显的浓度依赖性。
研究还表明,正常情况下结肠本身具有相当强的 H₂S 解毒能力,可将 H₂S 氧化为毒性更低的代谢物,例如亚硫酸盐。这意味着,结肠并不是被动暴露于 H₂S,而是长期处在“产生—解毒—利用”之间的动态平衡里。
问题通常出现在两个场景:
一是菌群失衡导致 H₂S 暴露增加;
二是暴露位置前移。
小肠对 H₂S 的解毒能力约只有结肠的 1/20。这意味着,一旦菌群失衡或 SIBO 让 H₂S 暴露更多发生在小肠,同样的 H₂S 负荷,其后果可能就会完全不同。
编辑
doi: 10.3390/ijms2607334
H₂S 为什么更值得放在黏膜环境里理解?
和 H₂、CO₂ 主要反映发酵是否活跃不同,H₂S 更值得放在黏液层—上皮屏障—局部炎症这个框架里理解。
现有研究提示,较高浓度的 H₂S 可抑制线粒体呼吸、影响 ATP 生成,并改变细胞内氧化还原状态;同时,它还会影响黏液层稳定性,增加上皮暴露风险。
——炎症性肠病
研究普遍发现,IBD 患者肠道内 H₂S 暴露增加,SRB 相关菌群也更活跃。IBD 患者结肠腔内 pH 往往高于健康对照,这会为 SRB 提供更有利的生长环境。
与此同时,研究表明,H₂S 还会进一步影响结肠能量代谢。H₂S 可干扰丁酸盐氧化,削弱结肠细胞对丁酸的利用。考虑到丁酸本来就是结肠上皮的重要能量来源,这种变化会让黏膜环境更脆弱,也更容易进入炎症状态。
——结直肠癌
现有研究已经提示,多种 H₂S产生菌 与肿瘤发生发展有关,尤其是 Fusobacterium nucleatum 这类既参与肿瘤微环境、又可参与 H₂S 生成的菌。
H₂S 在肿瘤发生中可能参与多个过程,包括细胞代谢调节、血管生成、细胞周期调控以及 DNA 损伤相关效应。
硫化氢升高,通常说明什么?
从肠道代谢角度看,H₂S 升高通常提示的不是单纯“产气增多”,而是以下几件事可能同时存在:
所以,H₂S 更适合放在“耗氢分流—硫代谢负荷—黏膜环境风险”这条线上看。
氨:常被忽略的蛋白发酵代谢产物
氨主要来自氨基酸脱氨作用
和前面几类气体相比,氨(NH₃)在肠道产气讨论里经常被忽略,但在肠道代谢层面,它同样是一个重要信号。
研究提到,NH₃ 主要由结肠细菌对氨基酸进行脱氨作用后产生。它和 H₂、CO₂、CH₄ 这类主要以气体形式存在的成分不同,NH₃ 在胃肠道内主要溶解于液相环境中。
研究表明,人类粪便中检测到的 NH₃ 浓度大约在 10 – 30 mM,而且随着蛋白质摄入增加,NH₃ 的排泄量也会增加。
氨和哪类底物关系更密切?
如果说 H₂ 和 CO₂ 更直接对应碳水代谢,那么 NH₃ 更直接对应的是蛋白质及氨基酸代谢。它的生成意味着,进入结肠的含氮底物正在被菌群进一步分解。
也就是说,NH₃ 更多反映的是肠道内蛋白发酵这一端的代谢活动。
它不是典型的“排气体积贡献者”,但在结肠代谢环境中,NH₃ 所代表的含氮代谢负荷值得注意。
氨升高后,会对肠道产生什么影响?
NH₃ 可通过改变肠道细胞形态和中间代谢,对细胞产生多方面毒性作用,包括抑制 DNA 生物合成以及缩短细胞寿命。
此外,一些研究还发现,较高水平的 NH₃ 可损伤结肠黏液层,并降低结肠细胞的寿命;在动物研究中,升高的 NH₃ 还与抑制结肠细胞对丁酸的氧化利用有关。
NH₃ 的问题在于它会直接影响结肠上皮细胞所处的代谢环境。
氨升高,通常说明什么?
从肠道代谢角度看,NH₃ 升高通常提示的是蛋白发酵负荷增加,以及结肠内含氮底物代谢活跃。它反映的重点在肠道局部代谢环境是否承受了更高的含氮代谢压力。NH₃ 更适合放在“蛋白发酵—黏膜环境—上皮代谢”这条线上看。
近年的研究还开始把 NH₃ 的意义从肠道局部环境延伸到更广的系统轴线。已有系统综述和 Meta-analysis 提出,肠道内源性 NH₃ 与 H₂S 可能还与男性生殖健康相关。
这项荟萃分析发现,体内氨(NH3)和/或硫化氢(H2S)与对男性生育有益的乳杆菌呈负相关,而氨和硫化氢与对男性生育有负面影响的拟杆菌属呈正相关。对乳杆菌属和拟杆菌属水平与氨进行比较的荟萃分析显示,结果具有统计学意义(p<0.001)。
不过,这部分内容目前可视为前沿方向,而不是已经定论的临床结论。
甲烷:更值得关注的是肠道动力变化
在肠道主要气体中,甲烷(CH₄)的来源和 H₂、CO₂ 不太一样。CH₄ 主要不是由普通细菌直接产生,而是由产甲烷古菌(methanogens)利用 H₂ 作为电子供体,将 CO₂ 还原后生成。
甲烷的出现同时说明肠道内前端已经存在 H₂ 和 CO₂ 的持续生成,并且下游存在活跃的甲烷生成通路。
哪些菌群参与了甲烷生成?
人类胃肠道内最主要的产甲烷微生物属于 Euryarchaeota。其中,最常见、最重要的代表是 Methanobrevibacter smithii。
此外,Methanosphaera stadtmanae 和 Methanobrevibacter oralis 也被认为参与了 CH₄ 的生成。
研究还提到,人体粪便中产甲烷古菌的丰度差异很大,可从几乎检测不到到 10⁹ CFU/g stool 不等;而只有当其数量达到大约 10⁷ 时,呼气中才更容易检测到 CH₄。
甲烷生成会带来什么代谢后果?
甲烷生成有一个很重要的特点:它能够明显减少总气体摩尔数。对应反应为:
CO₂ + 4H₂ → CH₄ + 2H₂O
也就是说,原本的 5 mol 气体,最终会变成 1 mol 气体。从这个角度看,甲烷生成实际上会降低总体气体体积。
甲烷相关问题的重点,往往在于它改变了气体代谢的去向,也改变了肠道内 H₂ 的分流格局。
从机制上看,这是一条非常典型的耗氢通路。
也就是说,上游发酵菌把未被小肠吸收的底物分解后,产生 H₂、CO₂ 和短链脂肪酸;下游产甲烷古菌再把 H₂ 和 CO₂ 进一步转入甲烷通路。
这一点很关键。因为 H₂ 在肠道里并不会无限累积。研究表明,较高的 H₂ 分压会抑制发酵菌继续代谢底物,并影响 NADH 再氧化以及后续发酵过程。换句话说,把 H₂ 持续移走,本身就是维持发酵系统继续运转的一部分。 甲烷生成正是这套移走 H₂的方式之一。
编辑
甲烷和便秘经常一起出现
较高水平的 CH₄ 与肠道动力减慢相关,并且和便秘,尤其是 IBS-C(便秘型肠易激综合征) 的关系更为密切。
一些研究还显示,CH₄ 可增强肠道收缩活动,同时减慢肠内容物推进速度。
也就是说,甲烷相关问题的重点,并不主要体现在总气体体积上,而更多体现在动力学后果上。
甲烷可能不仅影响动力,也和能量提取效率有关
一项研究发现,高甲烷产生者并没有表现出更高的 SRB 或 产乙酸菌活性下降,也就是说,甲烷通路未必是在压制其他耗氢通路;但高甲烷产生者却表现出更高的血清丙酸水平,并且在高纤维、全食物模式饮食下,具有更高的宿主可代谢能量(metabolizable energy, ME)。
进一步的网络分析还发现,和 CH₄ 生成正相关的,不只是 Methanobrevibacter smithii,还包括一组更适合降解膳食纤维、发酵纤维降解产物并生成丙酸的菌群,例如 Bacteroides caccae、Bifidobacterium adolescentis、Coprococcus、Eubacterium、Lachnospira、Prevotella copri、Roseburia faecis 等。
与甲烷产生率呈正相关的微生物
编辑
doi: 10.1093/ismejo/wraf103
这提示,甲烷有时更像是一个微生物协同网络的标志:它背后可能对应的,是一整套更擅长处理纤维底物、维持发酵链条、促进短链脂肪酸生成和吸收的菌群结构。
甲烷升高,通常说明什么?
从肠道代谢角度看,CH₄ 升高通常提示的是:
从实际理解上看,CH₄ 更适合放在“耗氢通路—肠道动力—微生物能量提取效率”这条线上看。提示气体代谢结构和肠道动力状态可能已经发生了变化。
上一章节我们了解到,H₂、CO₂、H₂S、NH₃、CH₄ 这些产物,来源不同、代谢路径不同,对宿主的影响也不同。也正因为如此,临床上看起来都叫“腹胀”“排气多”或“吃完不舒服”,背后的机制并不一定相同。
气体本身只是结果,真正决定症状和结局的,是气体类型、代谢去向以及宿主对这套代谢输出的承接能力。
肠道气体本身并不一定带来症状,关键在于能否被及时清除
肠道菌群发酵产生气体,本身并不等同于一定会出现不适。研究提到,发酵过程中产生的气体主要有三种去路:
一项研究显示,大约 77% 的微生物气体会通过前两种方式被清除,以排气形式离开的约占 23%。
这意味着,对多数健康人来说,产气和清除之间通常可以维持相对稳定的平衡。气体生成本身是结肠发酵的一部分,问题往往出在这套清除机制是否还能及时承接当前的发酵输出。
为什么有些人更容易腹胀、排气多?
主诉频繁腹胀和排气增多的人群,往往存在肠道气体清除受损。也就是说,他们的问题不一定只是产气更多,还可能是气体在肠道内停留得更久、清除得更慢。
另一要点是,功能性胃肠道症状患者对气体负荷的耐受阈值往往更低。
研究提到,这类人群相对正常人可以耐受的气体量,也更容易出现腹胀、胀满感和不适。也就是说,同样的气体负荷,在不同宿主背景下,主观症状的强弱并不一致。
这一点也和近期的干预研究相呼应。
一项随机、双盲、安慰剂对照试验纳入了 350 名自述存在腹胀/消化不良症状的健康成年人,连续 6 周给予多菌株合生元或安慰剂。结果显示,干预组不仅腹胀和胀气评分下降,腹部不适也显著改善,同时便秘相关不适和排便规律性也有所改善。
研究者指出,这类变化可能并不只是来自单纯产气减少,还与胃肠动力改善、内脏高敏感缓解以及肠道屏障功能支持有关。
这种差异在实际菌群报告中也很常见。看起来都属于产气相关问题,但进一步拆开后,背后的代谢结构并不一样。
比如在谷禾的一些样本中,除了主诉腹胀或排气增多之外,还可以直接看到肠道产气本身已处于超标水平;进一步拆开后,又伴随对甲酚过多、丁酸盐不足、Bacteroides fragilis 增殖,以及腹胀相关的 Bacteroides、Clostridium、Proteobacteria 等线索。
编辑
<来源:谷禾肠道菌群检测数据库>
这样的结果提示,问题往往不只是气体多了,而更像是发酵负荷升高、腐败代谢增强、保护性代谢物不足和菌群生态失衡同时存在。也正因为如此,同样表现为腹胀或排气增多,背后的处理逻辑并不一样。
气体的产生位置和产生速度,也会影响症状表现
除了总量和清除能力,气体在肠道内的产生位置和产生速度同样重要。研究指出,肠道内气体的数量或分布位置出现异常,都可能导致不良症状。
例如,当碳水化合物在小肠内被细菌过度发酵时,更容易引起明显的腹胀和腹痛,这也是小肠细菌过度生长相关症状的重要机制之一。
像RFOs(raffinose family oligosaccharides)这类发酵速度较快的底物,更容易因为短时间内迅速产气,而增加肠道清除负担,从而放大腹胀等症状。
注:RFOs是一类由多个单糖(通常是葡萄糖、果糖和半乳糖)通过糖苷键连接而成的低聚糖。在植物中广泛存在,尤其是在豆类、十字花科蔬菜(如卷心菜和花椰菜)、全谷物和某些根茎类蔬菜中。
有些“产气食物”在持续摄入后,症状反而会减轻
研究提出,肠道清除气体的能力并不是固定不变的,而是可以被饮食改变的。
例如,在刚开始摄入GOS这类短链、发酵较快的非消化性碳水时,总气体体积可增加 37%;但在连续摄入两周后,气体体积又会下降回基线水平。与此同时,在到达肛门前就被清除掉的气体比例,还呈现上升趋势。
这说明,持续摄入某些产气性食物后,肠道菌群的耗气能力和宿主对气体的处理能力可能会发生适应。一些所谓“容易产气”的食物,并不一定会长期维持同样强度的不适,症状程度可能随着时间而改变。
在炎症性肠病中,硫化氢相关信号更值得警惕
临床研究和实验研究都提示,菌群产生的 H₂S 与 IBD,尤其是溃疡性结肠炎(UC),存在关联。体外研究发现,UC 患者粪便样本中的硫酸盐还原菌数量高于健康对照,同时 H₂S 生成水平也更高。
还有研究发现,去除富含含硫氨基酸的食物,如 牛奶、鸡蛋和奶酪,在部分溃疡性结肠炎场景中曾显示出一定治疗意义。这提示,H₂S 所对应的也可能反映结肠局部代谢环境的改变。
甲烷菌和硫酸盐还原菌,与结直肠癌有关
多项研究还发现,甲烷菌和硫酸盐还原菌相关通路与结直肠癌(CRC)存在关联。
结直肠癌患者中,CH₄ 排出的发生率高于健康对照;与此同时,较高水平的 H₂S 也出现在结直肠癌高风险人群和溃疡性结肠炎患者中。
还有研究发现,在结直肠癌患者活检组织中,负责 H₂S 解毒的酶硫代硫酸盐硫转移酶丰度较低。这些结果提示,肠道气体及其相关代谢通路的意义,可能已经不只是功能症状层面,还涉及肠道局部病理环境的变化。不过,目前这类关联背后的分子机制仍有待进一步研究。
我们知道导致肠道气体的原因很多,但食物是影响产气症状的主要因素之一。食物之所以会影响肠道产气,关键是其中有哪些成分没有在上消化道被充分消化吸收,最终进入结肠,成为菌群发酵的底物。
与肠道气体关系最密切的食物类型,主要包括豆类、部分蔬菜和水果、全谷物、乳制品。这些食物虽然来源不同,但在机制上有一个共同点:都可能为结肠菌群提供可发酵底物,从而增加气体生成。
豆类——RFOs
豆类,包括干豆、扁豆、豌豆等,是很多人日常饮食中的重要组成部分。它们通常脂肪含量较低,同时富含蛋白质、维生素、矿物质和膳食纤维,因此在营养学上一直被视为具有较高价值的食物。
然而,尽管具有上述优点,许多消费者仍避免食用豆类,尤其是多种菜豆,原因是担心排气过多和胃部不适。
豆类引起的排气增加往往出现在摄入早期
有报道指出,部分人在饮食中加入豆类后,尤其是纤维摄入本就偏低的人,排气增加是预期结果。然而,有报道指出,随着更频繁地摄入豆类,排气会有所减少。
在一项随机对照试验中,50%的健康受试者报告在食用斑豆或烤豆的第一周内排气增加,但第二周后这一比例降至38%。
此后,报告胀气症状的受试者百分比在第6~12周持续下降至15%~23%。
在另一项随机对照试验中,评估了连续28天食用豆类(鹰嘴豆、扁豆和青豆)的影响。在干预早期,食用每种豆类都会引起胀气显著增加,但在后期胀气水平显著下降。
基于以上这些研究,排气的频率取决于个体食用豆类的频率,且个体之间的反应可能存在显著差异。
豆类中的关键底物是 RFOs
研究认为,豆类与肠道气体关系密切,主要与其中较高含量的膳食纤维和RFOs 有关。
研究指出,人类缺乏足够的 α-galactosidase 来降解这类寡糖,因此 RFOs 不能在小肠被充分消化吸收,而会进一步进入结肠,被菌群发酵,并在这一过程中产生气体。也正因为如此,豆类摄入后出现排气增加,在机制上是可以解释的。
不同豆类中的 RFOs 含量存在差异
扁豆、青豆、黄豆、豇豆、红芸豆等都属于 RFOs 含量相对较高的代表,不同种类之间在棉子糖、水苏糖、毛蕊花糖的含量上存在明显差异。
这说明,豆类引起的肠道产气并不是完全一致的,不同品种、不同底物组成,都会影响进入结肠后的发酵负荷。
编辑
加工方式可能影响豆类的产气潜力
除了原料本身的差异,综述还提到,一些加工处理方法可能降低植物中的 RFOs 含量,例如挤压、浸泡、高压灭菌、酶处理、煮沸。这些方法在不同程度上都有助于减少 RFOs,因此也被认为可能降低后续发酵产气。
从应用角度看,这部分研究的意义比较明确:豆类相关问题并不一定只能通过回避摄入来处理,底物含量本身和加工方式同样会影响最终的耐受性表现。
蔬菜和水果:果聚糖和多元醇
食用富含果聚糖的蔬菜后,人体可能会出现肠胃胀气。在蔬菜和水果中,与肠道气体关系更密切的是其中所含的某些可发酵碳水化合物。主要包括两类成分:一类是果聚糖,另一类是多元醇。
蔬菜和水果中的果聚糖属于菊粉型,由一个至多个与蔗糖的果糖部分相连的2→1连接的β-果聚糖基单元组成。此类果聚糖中最简单的物质是1-蔗果三糖,即β-果聚糖基-(2→1)-β-果聚糖基-(2→1)-α-吡喃葡萄糖苷。菊粉型果聚糖的聚合度通常在3~约60之间。
菊粉和低聚果糖(FOS)在蔬菜中含量丰富,如洋蓟、芦笋、菊苣根、大蒜、洋葱、蒲公英叶、韭菜等。
编辑
果聚糖在小肠中无法被消化,在结肠中发酵后产生过多肠道气体
果聚糖不能在小肠被消化吸收,因此会进入结肠,被菌群进一步发酵,从而增加气体生成。
最近的两项研究探讨了食用含菊粉食物对胃肠道症状的影响。排气增加是健康志愿者中最常见的胃肠道症状之一。
两项研究的结果均显示,健康志愿者在食用富含菊粉的饮食后,报告的最常见的胃肠道症状是肠胃气胀加重。
在另一项研究中,据报告,在连续5周食用富含低聚果糖的饮食期间,低聚果糖会导致肠道气体产生增加。
多元醇也是蔬菜和水果中重要的产气相关成分
多元醇天然存在于一些水果和蔬菜中,也被广泛作为无糖甜味剂用于口香糖和饮料中。
FDA批准的多元醇包括甘露醇、山梨醇、木糖醇、异麦芽酮糖醇、乳糖醇、麦芽糖醇、赤藓糖醇、氢化淀粉。
一项研究评估了山梨醇吸收不良与胃肠道症状之间的关系,该研究让7名健康参与者摄入不同剂量的山梨醇。在大多数受试者(7人中的4人)中,仅摄入5克山梨醇就与气体显著增加相关,而大多数受试者在摄入20克山梨醇后出现了严重症状(气体、腹胀、痉挛和腹泻)。
同样,在另一项包含10名健康志愿者的研究中,与摄入低剂量甘露醇相比,摄入含有高剂量甘露醇的水溶液导致的副作用发生率最高,包括肠胃气胀、腹泻和腹痛。
多元醇所带来的不适程度不仅与成分本身有关,也与摄入剂量密切相关。
蔬菜和水果相关的不适,本质上仍然是底物问题
从现有研究看,蔬菜和水果与肠道气体的关系,主要还是通过其中的果聚糖和多元醇来体现。
全谷物——果聚糖
全谷物常被视为健康食物,但并不意味着所有人都有同样的耐受性
小麦、玉米、大麦、黑麦、燕麦等谷物是世界大多数人口主要依赖的主食。与此同时,营养学和公共健康建议通常鼓励更多摄入全谷物而不是精制谷物。
《2020–2025美国膳食指南》建议每天至少摄入 3 盎司(约85克)全谷物,这主要基于全谷物与代谢健康相关的益处。
不过,这并不意味着全谷物在所有人群中都具有同样的胃肠耐受性,研究指出,全谷物虽然含有部分可发酵膳食纤维和抗性淀粉,但其中更值得关注的一个成分,是果聚糖。
全谷物中的果聚糖——与产气关系更密切的底物
谷物中的果聚糖与蔬菜水果中的果聚糖并不完全相同。前者主要属于左聚糖型,其结构特点是 β-呋喃果糖基单元通过 (2→6) 键连接到蔗糖分子上。这类果聚糖在谷物中的聚合度可达到约 90(说明它们属于结构相对复杂、可进入结肠发酵系统的一类底物)。
大量研究已将全谷物摄入与排气增加联系起来。
在一项非对照研究中,55%的IBS患者报告麸皮加重了其胃肠道症状,包括产气相关主诉。
关于麦麸的对照试验也报道,与安慰剂相比,参与者出现更多排气和腹部不适。
此外,Vuholm等考察了全麦和全黑麦对胃肠道症状的影响,发现摄入这些全谷物与健康志愿者排气增加显著相关。
部分人群需要关注FODMAPs
尽管小麦及其他谷物中的果聚糖等可发酵低聚糖、二糖、单糖、多元醇(FODMAPs)普遍被认为会在IBS患者中引起严重胃肠道症状,但也有研究指出,许多非乳糜泻性麸质敏感者实际上可能对小麦中的FODMAPs而非麸质敏感。
加工方式可能影响全谷物的产气潜力
已有研究开始评估不同工艺对谷物中果聚糖的影响。据推测可减少肠道气体及其他胃肠道症状。
一项研究评估了不同酸面团酵母菌株在96小时发酵过程中降解小麦粉中果聚糖的能力。发酵后,与传统面包酵母S. cerevisiae分离株相比,从澳大利亚酸面团中分离得到的Saccharomyces cerevisiae和Torulaspora delbrueckii能显著降低小麦粉中的果聚糖含量。
另一项研究还考察了,植物乳杆菌与酿酒酵母延长发酵时间在黑麦面团发酵过程中降解黑麦面包果聚糖的潜力。发酵3小时后,作者注意到,与仅用面包酵母发酵的面包相比,采用植物乳杆菌和酵母发酵的黑麦面包中果聚糖含量显著下降。
也就是说,原料本身之外,发酵工艺和加工方式同样会影响最终进入结肠的发酵底物负荷。
全谷物相关问题,更适合放在底物耐受性框架中理解
这里还可以区分两个层面来看:
前者有时可以结合菌群功能线索辅助理解,后者则更多与宿主本身的肠道动力、气体清除和感觉敏感性有关。
从干预角度看,全谷物相关不适更适合先放在果聚糖耐受的框架里理解,再进一步结合加工方式、摄入剂量和摄入频率做调整;如果同类底物反复诱发症状,后续也可以结合饮食记录、症状出现的时间窗,包括菌群线索,进一步判断问题更偏向发酵太强,还是本身更不耐受。
编辑
doi.org/10.1016/j.jff.2022.105367
乳制品——乳糖
乳制品相关不适,核心在于乳糖处理能力
牛奶和其他乳制品是加重胃肠道症状的食物之一,尤其是在全球相当一部分成年乳糖酶缺乏人群中。
乳糖是哺乳动物乳汁中特有的碳水化合物,人乳中含量约为7.2 g/100 ml,牛乳中约为4.7 g/100 ml。乳糖本身并不会直接引起气体问题,影响后续结局的是它能否在小肠被及时水解。
编辑
乳糖能否被消化,取决于乳糖酶活性是否足够
乳糖酶是负责水解乳制品中乳糖的酶。乳糖需要依赖乳糖酶水解为葡萄糖和半乳糖后,才能在小肠被吸收。
在大多数哺乳动物中,乳糖酶活性在儿童期较高,但会随年龄增长迅速下降。在人群中,仅约 30% 的人群在成年后仍保持乳糖酶持续表达,但这种乳糖酶持续性在不同人群中的分布差异很大。
这意味着,对相当一部分成年人来说,乳糖并不能在小肠被充分处理,而更容易进入结肠。
未被消化的乳糖会进入结肠并被菌群发酵
研究指出,当乳糖没有在小肠被充分水解为单糖时,它会进入大肠,成为菌群可利用的发酵底物。随后,肠道菌群会对其进行代谢,并产生相关气体。乳糖酶缺乏患者因结肠内乳糖发酵而出现排气增加。
乳糖进入结肠后,菌群组成会影响后续的代谢走向。某些菌群,尤其是双歧杆菌属和乳杆菌属,具有β-半乳糖苷酶活性,能够利用乳糖并完成进一步分解;同时,这两类菌更倾向于产生短链脂肪酸,而不像其他部分异型发酵菌那样更容易形成较高的氢气和甲烷输出。也就是说,乳糖进入结肠之后,会不会形成更明显的产气表型,和菌群如何处理这部分底物也有关系。
益生菌可能缓解乳糖不耐受症状的机制
编辑
doi: 10.1007/s10482-026-02278-x
部分人群对乳糖的耐受,可以随着菌群适应而变化
在乳糖不耐受人群中,长期、规律地接触乳糖后,肠道菌群可能发生适应性变化。
这种变化通常表现为具有β-半乳糖苷酶活性的乳糖发酵菌增加,例如双歧杆菌属和乳杆菌属丰度上升,同时结肠对乳糖的处理效率提高,呼气氢排出下降,症状减轻。
基于证据的、对乳糖不耐受有益的益生菌菌株
编辑
doi: 10.1007/s10482-026-02278-x
如果把肠道产气仅仅理解为吃了某类食物之后会不会胀,那么临床和健康管理层面的处理往往容易停留在经验性忌口、临时回避和泛化建议上。但从现有研究看,肠道产气所连接的并不只是食物和症状,还包括底物进入上消化道后的处理效率、结肠阶段的菌群代谢方式、不同耗氢通路的分流方向,宿主本身的动力和耐受状态等。这意味着,产气相关问题更适合被理解为一个可分层、可追踪、可干预的功能表型。
未来关于产气相关问题的管理,会包括对具体底物、具体通路和具体耐受阈值的识别。进一步判断:究竟是哪一类底物在反复诱发症状,进入结肠后被怎样的菌群结构处理,又是在什么剂量和什么摄入方式下开始越过个体的耐受边界。
肠道产气相关问题也越来越值得放到多维评估框架里看。饮食记录能帮助识别底物来源,症状时间窗能帮助判断反应时序,呼气试验能帮助捕捉功能性输出,而菌群相关信息则更适合帮助理解:个体的肠道,当前更偏向哪一种发酵和耗氢模式,是否已经出现了保护性代谢物不足、腐败代谢增强,或者硫代谢相关通路偏移。也就是说,菌群检测帮助我们把原本笼统的症状,推进到更具体的代谢理解和功能识别。
总的来说,肠道产气更像是肠道持续暴露自身代谢状态的一种外显方式。把这些信号读得更清楚,就更接近下一阶段更精细的营养干预和功能管理。
注:本账号内容仅供学习和交流,不构成任何形式的医疗建议。
主要参考文献
Hao Y, Du X, Cai C, Zhao Y, Ren Y. Ammonia and hydrogen sulfide – new insights into gut microbiota and male infertility through meta-analysis. Front Cell Infect Microbiol. 2025 Jan 6;14:1449453.
Dirks B, Davis TL, Carnero EA, Corbin KD, Smith SR, Rittmann BE, Krajmalnik-Brown R. Methanogenesis associated with altered microbial production of short-chain fatty acids and human-host metabolizable energy. ISME J. 2025 Jan 2;19(1):wraf103.
Birg A, Lin HC. The Role of Bacteria-Derived Hydrogen Sulfide in Multiple Axes of Disease. Int J Mol Sci. 2025 Apr 3;26(7):3340.
Chen B, Zhang Y, Du L, Jin J, Dai N. Advances in the mechanism of low FODMAP diet in the treatment of irritable bowel syndrome: a review. Front Nutr. 2026 Mar 12;13:1719048.
Allegretti JR, Kassam Z, Kelly CR, Grinspan A, El-Nachef N, Van Den Elzen C, Jäger R, Feuerstadt P. A Randomized, Placebo-Controlled Trial Evaluating Multi-Species Synbiotic Supplementation for Bloating, Gas, and Abdominal Discomfort. Nutrients. 2026 Jan 14;18(2):255.
Perets TT, Gingold-Belfer R, Dickman R. Lactose intolerance and probiotics: from pathophysiological mechanisms to clinical applications. Antonie Van Leeuwenhoek. 2026 Mar 17;119(4):67.
Bin Waqar SH, Rehan A. Methane and Constipation-predominant Irritable Bowel Syndrome: Entwining Pillars of Emerging Neurogastroenterology. Cureus. 2019 May 28;11(5):e4764.
Erasme Mutuyemungu, Mukti Singh, Sean Liu, Devin J. Rose, Intestinal gas production by the gut microbiota: A review, Journal of Functional Foods,Volume 100, 2023
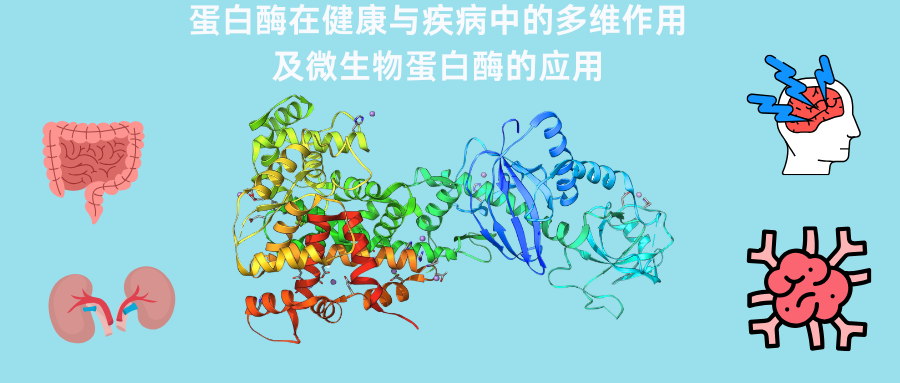
谷禾健康
你可能没听过“蛋白酶”,但它其实每天都在默默影响你的健康。
蛋白酶是一类帮助分解蛋白质的关键工具。但研究发现,它的作用远不止消化这么简单——它还参与调节免疫、影响肠道状态,甚至和很多常见不适有关。
本文结合近年来的研究进展,系统介绍了蛋白酶的基本概念、不同类型及其来源(包括人体自身和微生物)。重点讲解了蛋白酶在肠道消化中的作用,以及某些细菌产生的蛋白酶如何与肠道健康相关。例如,在炎症性肠病、肠易激综合征和乳糜泻等常见肠道问题中,蛋白酶可能参与影响肠道状态和症状的发生。
此外,还分析了多种会影响蛋白酶活性的因素,以及微生物蛋白酶如何通过特定受体(PARs)影响肠道屏障、细胞信号以及腹痛等不适感。
最后,结合实际健康管理需求,本文讨论一种更为主动的干预思路:通过持续的症状监测与生物标志物评估,配合个性化饮食调整及生活方式优化,系统性改善肠道微环境,并进一步探索恢复蛋白质分解平衡作为潜在的健康干预策略。
蛋白酶(Proteases,也叫proteinases或peptidases)是一类催化蛋白质中肽键水解的酶,是生命系统中不可或缺的分子工具。自1905年P. A. Levene在《生物化学杂志》上发表关于“蛋白胨裂解产物”的研究以来,该领域已发展百年。
在很长一段时间里,蛋白酶主要被视为执行蛋白质分解代谢和为生物体提供氨基酸的简单工具。然而,随着科学的进步,人们逐渐认识到,蛋白酶还能够催化高度特异性的蛋白水解反应,生成具有新功能的蛋白质产物,开启了蛋白酶研究的新阶段。
★ 蛋白酶在大部分生物中起核心作用
如今,蛋白酶被公认为在所有生物体的几乎所有生物学过程中都发挥着核心调控作用。它们通过精确的切割,调节着蛋白质的命运、定位和活性,调控蛋白质间的相互作用,创造新的生物活性分子(如激素和神经肽),并参与细胞信息的处理、转导和放大。
因此,蛋白酶深刻影响着DNA复制与转录、细胞增殖与分化、组织形态发生与重塑、血管生成、神经发生、排卵、受精、伤口修复、止血、炎症、免疫、自噬、衰老、坏死和凋亡等关键生命活动。
人类基因组已鉴定出600余个蛋白酶基因,约占总基因数的3%,凸显其在维持生命稳态中的核心地位。
相应地,蛋白水解系统失调与癌症、神经退行性、炎症性及心血管等多种疾病密切相关,使蛋白酶成为重要的药物靶点及诊断与预后生物标志物。
1
宿主来源的蛋白酶
宿主自身是蛋白酶的主要来源,这些内源性蛋白酶广泛分布于全身各组织和体液,承担多种生理功能。
• 蛋白酶在消化系统中大量分布且起重要作用
消化系统中,胃主细胞分泌的胃蛋白酶原在酸性环境下被激活为胃蛋白酶,启动蛋白质的初步消化。随后,胰腺分泌的多种蛋白酶原(如胰蛋白酶原、糜蛋白酶原)在十二指肠内经肠激酶及胰蛋白酶级联激活,进一步将蛋白质分解为小肽和氨基酸。
• 蛋白酶还参与组织稳态与病理过程
除了消化功能外,宿主蛋白酶在组织稳态与病理过程中同样至关重要。金属蛋白酶是最大的一类, 人类基因组中约有200个成员。其中,Meprins(Meprin metalloproteases)属于astacin家族,具有独特的寡聚金属蛋白酶结构:
由α、β两种亚基通过二硫键连接,形成同源或异源寡聚体。Meprin A以α亚基为主,主要为分泌型;Meprin B以β亚基为主,为膜结合型。它们在肾脏和肠道刷状缘膜高表达,参与生物活性肽及细胞外基质蛋白的加工。
比如,Meprin B可切割E-钙黏蛋白等细胞表面蛋白,影响细胞间黏附;Meprin A可切割紧密连接蛋白occludin,破坏上皮屏障。Meprins异常表达或活性与炎症性肠病(IBD)、肾脏疾病及癌症的发展密切相关。
Metzinicins、astacins和meprins的进化关系与结构
doi: 10.1074/jbc.TM118.004156.
2
微生物蛋白酶
肠道微生物群构成一个庞大且动态的蛋白酶库。细菌、真菌和古菌分泌的蛋白酶不仅可增强宿主消化能力,还能深度参与宿主-微生物相互作用。
相比动植物来源的蛋白酶,微生物蛋白酶通常具有生长周期短、产量高、生化性质多样(如耐酸、耐碱、耐高温)以及便于基因工程改造的优势,因此在工业与生物技术领域应用前景广阔。
• 细菌蛋白酶能补充人体内源性蛋白酶的不足
在生理功能上,细菌蛋白酶可补充人体内源性蛋白酶不足,尤其是在胰腺外分泌功能不全、老年人消化能力下降或胃酸缺乏等病理状态下。此时胃蛋白酶和胰蛋白酶分泌减少,导致蛋白质消化不良,未消化蛋白进入结肠,引起腹胀、腹泻并造成营养不良。
此时,肠道微生物,特别是具有强大蛋白水解能力的细菌,如:
芽孢杆菌属(Bacillus)、
假单胞菌属(Pseudomonas)、
肠球菌属(Enterococcus)、
拟杆菌属(Bacteroides)、
梭菌属(Clostridium)
• 肠道菌群产生的蛋白酶有助于肠道营养代谢
它们产生的蛋白酶可以在肠道的中性或弱碱性环境中发挥作用,降解宿主未能消化的蛋白质,从而在一定程度上补偿了宿主消化酶的不足。例如,来源于解淀粉芽孢杆菌(Bacillus amyloliquefaciens)的工业级蛋白酶已被开发为口服消化酶补充剂,用于替代疗法,显示出良好的临床价值。
拟杆菌属作为肠道中的优势菌群,其基因组中编码了大量的糖苷水解酶和蛋白酶,使其能够高效利用各种膳食多糖和蛋白质,是肠道营养物质代谢的关键参与者。
1
基于催化机制的功能和分类
随着对结构和催化机制认识的加深,蛋白酶的分类不断完善。最初,蛋白酶根据其切割位点被简单地分为内肽酶(endopeptidases,切割蛋白质内部的肽键)和外肽酶(exopeptidases,从蛋白质的N-末端或C-末端切割肽键)。外肽酶又可细分为氨肽酶和羧肽酶。
注:根据国际生物化学与分子生物学联合会(IUBMB)的酶学委员会(EC)命名法,蛋白酶被归类为水解酶(EC 3)中的肽键水解酶(EC 3.4)。
目前最常用的分类依据其活性位点的化学性质和催化机制,将其分为六大类。
•丝氨酸蛋白酶(Serine Proteases):活性位点含有丝氨酸残基,其羟基作为亲核试剂。例如胰蛋白酶、胰凝乳蛋白酶。
•半胱氨酸蛋白酶(Cysteine Proteases):活性位点含有半胱氨酸残基,其巯基作为亲核试剂。例如木瓜蛋白酶、组织蛋白酶。
•天冬氨酸蛋白酶(Aspartic Proteases):活性位点含有两个天冬氨酸残基,它们协同激活一个水分子作为亲核试剂。例如胃蛋白酶、HIV蛋白酶。
•金属蛋白酶(Metalloproteases):催化需要一个或多个金属离子(通常是锌离子)作为辅因子,金属离子激活水分子。例如胶原酶、基质金属蛋白酶(MMPs)、Meprins。
•苏氨酸蛋白酶(Threonine Proteases):活性位点含有苏氨酸残基,其N-末端苏氨酸的羟基作为亲核试剂。例如蛋白酶体的催化亚基。
•谷氨酸蛋白酶(Glutamic Proteases):活性位点由谷氨酸和谷氨酰胺残基组成,激活水分子进行催化。此类蛋白酶主要在真菌和细菌中发现,哺乳动物中尚未发现。
为了更系统化整理庞大的蛋白酶家族,研究人员依据氨基酸序列同源性将蛋白酶划分为不同“families”,再结合三维结构相似性将其进一步归类为“氏族”(clans)。MEROPS数据库是目前最权威的蛋白酶及其抑制剂的在线资源,它详细记录了这种分层分类系统,为蛋白酶的进化和功能研究提供了宝贵信息。
代表性真核生物基因组中蛋白水解景观的全球视图
doi: 10.1074/jbc.R800035200.
该图展示了人类、小鼠、果蝇和拟南芥中蛋白酶在催化类别及其相关成员家族中的分布。
其中催化类别用天冬氨酸(A)、半胱氨酸(C)、苏氨酸(T)、丝氨酸(S)和金属蛋白酶(M)表示。例如,人类天冬氨酸蛋白酶A01家族包含5个成员,而拟南芥中该家族有55个。
2
基于底物特异性的分类
微生物蛋白酶因来源广泛、功能多样而广受工业与生物技术领域的青睐。根据来源、性质等多维标准可对其进行精细分类,从而更好地揭示其功能特异性,并为应用提供指导。
这类蛋白酶可水解特定蛋白底物,在组织重塑及特定工业应用中发挥关键作用。
•胶原酶:主要负责降解胶原蛋白——结缔组织中最丰富的蛋白质。微生物胶原酶(如梭菌属Clostridium 的相关酶)在伤口清创、组织分离和细胞培养中应用广泛。
•弹性蛋白酶:特异性降解弹性蛋白,对维持血管与皮肤弹性至关重要。来自假单胞菌属(Pseudomonas)的弹性蛋白酶常作为研究病原菌侵袭机制的重要模型。
•角蛋白酶(Keratinases):能够降解难溶的角蛋白,如羽毛、毛发和指甲。来源于芽孢杆菌属(Bacillus)和曲霉属(Aspergillus)的角蛋白酶在处理羽毛废料、生产动物饲料和皮革工业的生物脱毛中显示出巨大潜力。
3
基于与经典蛋白酶同源性的分类
许多微生物蛋白酶在结构和功能上与哺乳动物的经典消化蛋白酶相似,这为它们在食品工业中的应用提供了理论基础。
•胰蛋白酶样蛋白酶:切割赖氨酸(Lys)或精氨酸(Arg)残基的羧基端肽键。例如,某些链霉菌(Streptomyces)产生的胰蛋白酶具有与牛胰蛋白酶相似的底物特异性。
•胰凝乳蛋白酶样蛋白酶:优先切割芳香族氨基酸(如苯丙氨酸、酪氨酸、色氨酸)残基的羧基端。
•胃蛋白酶样蛋白酶:属于天冬氨酸蛋白酶,在酸性条件下切割疏水性氨基酸残基之间的肽键。来源于真菌(如曲霉属)的酸性蛋白酶在功能上模拟胃蛋白酶,用于蛋白质水解。
拓展:蛋白酶vs.淀粉酶vs.脂肪酶
蛋白酶是指一类分解蛋白质的酶。胃蛋白酶在胃中启动消化,胰蛋白酶和乳糜胰蛋白酶由胰腺产生并释放到小肠,帮助将蛋白质分解为氨基酸,随后被循环吸收。
淀粉酶是消化酶,可将淀粉分解为简单糖类以供能。它主要来自唾液腺;因此一开始咀嚼就会启动消化。随后胃淀粉酶继续将食物分解为糜粒,并促使分泌素释放,推动胰腺分泌胰酶,从而完成消化。
脂肪酶同样是消化酶,能分解膳食脂肪以供肠道吸收。它主要由胰腺分泌(胰腺脂肪酶),但也存在于血液、胃液、肠液和脂肪组织中。胆汁通过把脂肪乳化成小脂肪球来启动消化;随后脂肪酶将脂肪球分解为脂肪酸和甘油,甘油为多种脂质中的基础小分子,细胞可将其用于获取能量。
1
肠道内肽酶与外肽酶在消化中的作用
蛋白质消化是多步骤过程,依赖内肽酶与外肽酶协同完成。食物蛋白先由胃蛋白酶及胰腺分泌的内肽酶(如胰蛋白酶、糜蛋白酶)切割为较大多肽。
随后,肠刷状缘膜肽酶与细胞内肽酶,以及微生物分泌的外肽酶(氨肽酶、羧肽酶)进一步将多肽水解为寡肽、二肽和游离氨基酸,最终由肠上皮吸收。
• 未分解的蛋白质还可成为肠道菌群的发酵底物
在肠道远端,未被宿主完全消化的蛋白质与肽类成为微生物发酵底物。在厌氧消化中,微生物蛋白酶承担关键“水解”作用,过程可分为四阶段:水解、产酸、产乙酸与产甲烷。
蛋白质在水解阶段被分解为氨基酸,随后在产酸、产乙酸和产甲烷阶段被微生物群落逐步转化为甲烷、二氧化碳等最终产物。
水解阶段,复杂蛋白质在微生物(如 Clostridium、Proteiniborus)分泌的胞外蛋白酶作用下分解为氨基酸;随后氨基酸在产酸阶段经Stickland反应等途径继续分解,生成挥发性脂肪酸(VFAs)、氨及硫化氢等产物。以丙氨酸和甘氨酸为例,它们可在Stickland反应中转化为乙酸并伴随产能(ATP)。该过程既为微生物提供能量与氮源,也使代谢产物(如短链脂肪酸)影响宿主能量代谢与免疫调节。
2
蛋白酶的信号传导:蛋白酶激活受体
蛋白酶不仅是消化酶,更是重要的信号分子。它们可通过激活蛋白酶激活受体(Protease-Activated Receptors, PARs)调控细胞功能。
• 蛋白酶激活受体与通透性、炎症调节等相关
PARs目前已知有四种成员(PAR1、PAR2、PAR3、PAR4),广泛表达于肠道上皮细胞、免疫细胞、神经元和内皮细胞。
其独特激活机制称为“系链配体模型”:蛋白酶切割受体N端特定位点,暴露新的N-末端序列;该序列作为“系链配体”与受体跨膜结构域结合,从而激活受体并引发下游信号,包括钙离子动员、MAPK通路激活以及基因转录等。
在胃肠道中,PARs的激活参与多种生理与病理过程。例如,PAR1激活与上皮细胞凋亡及通透性增加相关;PAR2激活则与炎症、内脏痛觉过敏及屏障功能调节密切相关。
编辑
宿主胰蛋白酶是PAR2和PAR4的天然激动剂,肥大细胞释放的类胰蛋白酶同样可激活PAR2。更关键的是,肠道微生物蛋白酶(如牙龈卟啉单胞菌的gingipains、艰难梭菌毒素A)已被证实可直接激活PARs,进而介导炎症反应并破坏屏障。因此,PARs是宿主与微生物蛋白酶信号交流的重要枢纽。
3
Meprins在组织与疾病中的双刃剑作用
Meprins作为astacin金属蛋白酶家族成员,在组织重塑与炎症反应中发挥“ 双刃剑”作用:一方面参与ECM降解与重塑、生长因子激活及生物活性肽加工等正常生理过程;另一方面,其异常表达或活性失调与多种疾病的发生发展密切相关。
•炎症性肠病(IBD):研究发现,MEP1A基因(编码meprin α亚基)是IBD(尤其溃疡性结肠炎)的易感基因之一。Meprin α的表达下调或功能缺失会加重实验性结肠炎,其机制可能与调控肠道屏障及降解促炎细胞因子有关。Meprin A可切割紧密连接蛋白occludin,破坏上皮屏障完整性并促进单核细胞迁移。
•肾脏疾病:敲除小鼠的Meprin β基因可减轻缺血再灌注引起的肾损伤,提示Meprin β在肾损伤中具有促炎和促纤维化作用。此外,人类MEP1B基因多态性与皮马印第安人的糖尿病肾病有关。
•癌症:Meprins在多种癌细胞(如结肠癌和乳腺癌)中异常表达,被认为可通过降解细胞外基质、促进细胞迁移与侵袭,进而参与肿瘤进展和转移。
•神经退行性疾病:Meprins能够切割阿尔茨海默病(AD)的关键蛋白淀粉样前体蛋白(APP),提示其可能参与AD的发生与进展。
肠道菌群失调是多种胃肠道疾病的关键病理生理环节,而失衡菌群产生的蛋白酶是衔接菌群变化与宿主病理反应的重要介质。它们可通过多种机制破坏肠道稳态,从而诱发或加重疾病。
1
炎症性肠病
炎症性肠病(包括克罗恩病与溃疡性结肠炎)病因复杂,但普遍认为其发生与遗传易感宿主对肠道微生物的异常免疫应答有关。已有大量证据显示,IBD患者粪便中的蛋白酶活性显著高于健康人群,且该活性主要来源于微生物。
• 炎症性肠病患者蛋白酶活性相比正常人更高
研究发现,IBD患者粪便中具有明胶水解活性的微生物多样性更高。进一步的高通量测序分析表明,粪便中的胰蛋白酶样活性与特定菌群丰度密切相关:毛螺菌科(Lachnospiraceae)和链球菌科(Streptococcaceae)的丰度与蛋白酶活性呈正相关;而有益的瘤胃球菌科(Ruminococcaceae)则呈负相关。
• 过量蛋白酶破坏屏障完整性并加重炎症
这些过量微生物蛋白酶可激活上皮细胞PAR2受体,启动下游信号通路(如ERK1/2、MAPK),从而促进紧密连接蛋白ZO-1的磷酸化并引发其重新分布,破坏屏障完整性、增加通透性。屏障受损后,更多微生物抗原(如LPS)进入黏膜下层,持续刺激免疫反应,形成恶性循环并加重炎症。
其中,粪肠球菌(Enterococcus faecalis)分泌的明胶酶GelE可直接降解上皮细胞间关键粘附分子E-钙粘蛋白(E-cadherin),导致细胞连接松散、屏障功能受损。
在IL-10基因敲除小鼠中,定植野生型粪肠球菌会诱发严重结肠炎,而定植缺乏GelE的突变株则炎症显著减轻,直接证实微生物蛋白酶在IBD发病中的致病作用。
肠道微生物蛋白酶依赖性肠道通透性增加
编辑
doi: 10.3748/wjg.v19.i43.7531.
微生物蛋白酶通过激活上皮细胞的PARs,启动细胞内信号传导(如ERK1/2和MAPK),破坏紧密连接,导致肠道通透性增加。这使得微生物及其蛋白酶能够穿透屏障,进一步作用于免疫细胞和细胞因子(如IL-8),加剧炎症反应。
2
乳糜泻
乳糜泻(Celiac Disease)是一种对麦麸(gluten)中谷蛋白(gliadin)产生的自身免疫性疾病。
• 乳糜泻患者菌群组成改变,谷蛋白肽异常加工
谷蛋白中富含脯氨酸和谷氨酰胺的肽段对宿主消化酶具有抵抗性,未被完全消化的肽段可诱发免疫反应。其中,长达33个氨基酸的肽段(33-mer)被认为是主要的免疫毒性肽。
肠道微生物参与谷蛋白降解,部分细菌(如拟杆菌属)可产生能够切割脯氨酸富集区域的蛋白酶。乳糜泻患者中,菌群组成与功能改变,可能导致谷蛋白肽异常加工,生成更具免疫原性的肽段。
• 利用微生物蛋白酶可能是治疗乳糜泻的新方法
另一方面,利用微生物蛋白酶开展“酶疗”被认为是乳糜泻治疗的重要研究方向。研究显示,米曲霉(Aspergillus oryzae)来源的脯氨酸特异性内肽酶,或细菌(如Pseudomonas aeruginosa)产生的谷蛋白酶(glutenases),可在体外有效降解包括33-mer在内的免疫原性谷蛋白肽。
相关酶制剂有望作为口服补充剂,在谷蛋白进入小肠之前将其分解为无害片段,从而预防或缓解乳糜泻症状。
3
肠易激综合征
肠易激综合征(IBS),特别是腹泻型IBS(IBS-D),其核心症状是腹痛和排便习惯改变。
• IBS-D患者类胰蛋白酶活性升高致内脏高敏
研究显示,IBS-D患者粪便上清液中的类胰蛋白酶活性显著升高,约为对照组的4倍。将这些上清液灌注至小鼠结肠可诱导结肠通透性增加并出现内脏高敏感(疼痛阈值降低)。机制研究表明,该效应由蛋白酶介导:蛋白酶通过激活PAR2受体发挥作用。
PAR2不仅存在于上皮细胞,也表达于感觉神经末梢;其激活可直接提高神经元兴奋性,引发腹痛。进一步地,蛋白酶激活感觉神经元上的PAR2,增强TRPV1等伤害性感受器活性,从而降低痛觉阈值,构成IBS腹痛与内脏高敏感的重要分子基础。
编辑
因此,肠道微生物蛋白酶活性的升高被认为是连接菌群失调、肠道屏障功能障碍和内脏高敏感性这三个IBS关键病理环节的核心机制。
4
胃肠道感染
在胃肠道感染中,病原菌分泌的蛋白酶通常作为重要的毒力因子,直接参与致病过程。一个典型的例子是艰难梭菌(Clostridium difficile)感染,其产生的毒素A和毒素B是主要的致病因子。
• 蛋白酶可作为毒力因子削弱屏障促进病原菌定植
研究表明,毒素A可通过激活PAR2受体间接诱导肠道炎症与液体分泌,进而导致腹泻和伪膜性结肠。同时,多种肠道病原体(如致病性大肠杆菌EPEC/EHEC和志贺氏菌)会分泌丝氨酸蛋白酶自身转运蛋白(SPATEs),例如Pic蛋白酶,能降解黏液层黏蛋白,削弱第一道物理屏障,促进细菌粘附与定植。
编辑
• 益生菌蛋白酶的保护作用
相反,部分益生菌可产生蛋白酶以对抗病原体:布拉氏酵母菌(Saccharomyces boulardii)分泌54 kDa丝氨酸蛋白酶,可直接降解艰难梭菌毒素A和毒素B,并抑制毒素与肠上皮细胞受体的结合,从而中和其致病作用。这体现了微生物蛋白酶在肠道中的双重效应——既可能是“破坏者”,也可作为“保护者”。
蛋白酶的活性受到多种内外因素的精密调控,这些因素共同决定了蛋白水解事件在何时、何地以及以何种强度发生。理解这些影响因素对于研究蛋白酶功能和开发相关应用至关重要。
1
物理化学因素
• pH值
pH是影响蛋白酶活性的关键因素之一。不同蛋白酶的最适pH不同:胃蛋白酶在pH 1.5–2.5的强酸环境活性最高,胰蛋白酶在pH约8.0的弱碱环境中表现最佳。
微生物蛋白酶的pH适应性更广,常分为三类:酸性蛋白酶(最适pH 2.0–6.0),主要来自曲霉属和根霉属,常用于食品与饮料工业;中性蛋白酶(最适pH接近中性),多见于芽孢杆菌属和曲霉属,用于烘焙与肉类嫩化;碱性蛋白酶(最适pH 8.0–13.0),主要来源于芽孢杆菌属,是洗涤剂工业的重要成分。
• 温度
温度会同时影响酶促反应速率与酶稳定性。多数蛋白酶在特定温度范围内活性最高,超出范围可能因热变性而失活。
嗜温微生物来源的蛋白酶通常在25–40°C最活跃;而嗜热微生物(如Bacillus stearothermophilus)的蛋白酶可在50–65°C甚至更高温度下保持稳定与高活性,因此在高温工业应用(如洗涤剂)中尤具价值。
2
生化因素
• 金属离子
许多蛋白酶,尤其是金属蛋白酶,其催化活性高度依赖金属离子。锌(Zn²⁺)是最常见辅因子,直接参与astacin和MMP等家族酶的活性位点催化。钙(Ca²⁺)和镁(Mg²⁺)等离子虽不直接参与反应,但常作为稳定剂与酶蛋白结合,维持正确三维构象,从而提升酶的热稳定性与活性。
• 前肽(Propeptide)的调控
许多蛋白酶以无活性的酶原形式合成,N端或C端带有前肽序列。前肽一方面作为分子内伴侣,帮助新生肽链正确折叠形成具催化潜能的构象;另一方面作为抑制剂,通过占据或封闭活性位点,阻止蛋白酶在合成与运输过程中提前激活。蛋白酶只有在特定生理条件下,经有限蛋白水解切除前肽(自身或由其他蛋白酶)后才会被激活。例如,Meprin α亚基的成熟与分泌依赖前肽的正确加工。
• 特异性抑制剂
生物体内存在多种内源性蛋白酶抑制剂,可与蛋白酶形成复合物,精确调控蛋白水解活性。例如,丝氨酸蛋白酶抑制剂(Serpins)借助“自杀性底物”机制对靶蛋白酶进行不可逆抑制;组织金属蛋白酶抑制剂(TIMPs)则可逆抑制MMPs。
微生物也会产生抑制剂,如部分双歧杆菌的Serpins可能有助于维持肠道蛋白水解平衡。药物研发中同样常用小分子抑制剂,例如Pepstatin A可强效抑制天冬氨酸蛋白酶。
野生型meprin α及其突变体在内质网中的折叠、加工
编辑
doi: 10.1074/jbc.TM118.004156.
该图直观表明,前肽及MAM等结构域对蛋白酶的正确折叠、成熟与转运至关重要。缺失MAM结构域(ΔMAM)会导致蛋白酶错误折叠并被蛋白酶体降解,进一步说明分子伴侣与质量控制机制对蛋白酶功能的关键作用。
微生物蛋白酶的准确检测是筛选新型酶、优化生产工艺、解析生理功能及研究病理作用的基础。检测方法按目的分为定性与定量两类,且随着技术进步不断发展出高灵敏度的新方法。
1
定性检测:琼脂平板透明圈法
琼脂平板透明圈法(Halo Formation Assay)是经典且直观的蛋白酶活性定性筛选方法。将不溶性或大分子蛋白底物均匀分散于琼脂培养基,形成浑浊背景。接种能分泌胞外蛋白酶的微生物后,若其产生蛋白酶,会将周围蛋白底物水解为可溶性小分子肽和氨基酸,从而在菌落周围形成透明区域(“水解圈/透明圈”)。透明圈大小通常与蛋白酶产量和活性正相关,因此适用于初步筛选高产菌株。
常用的蛋白质底物包括:
• 脱脂牛奶琼脂:这是最常用的底物:成本低、制备方便且效果直观。牛奶中的酪蛋白(casein)是主要蛋白质,蛋白酶水解后,浑浊的乳白色培养基会变透明。
• 酪蛋白琼脂:使用纯化的酪蛋白作为底物,背景更均匀,结果更具可比性。
• 牛血清白蛋白琼脂:BSA是一种标准蛋白质,用于需要更精确控制底物浓度的研究。
• 明胶琼脂:明胶是胶原蛋白的水解产物。蛋白酶水解明胶后,培养基在低温下不再凝固,或在染色后(如用酸性品红)显示出清晰的降解区。
• 角蛋白琼脂(Keratin Agar)和弹性蛋白琼脂(Elastin Agar):用于筛选能降解特定底物的蛋白酶,例如角蛋白酶和弹性蛋白酶。将相应底物以粉末加入培养基,降解后即可形成透明圈。
此类方法主要适用于检测能够降解大分子蛋白质的内肽酶。对于外肽酶或活性较弱的蛋白酶,可能无法形成肉眼可见的透明圈。此外,该方法只能进行半定量评估,精确的活性测定需要依赖定量方法。
2
定量检测:分光光度法
分光光度法通过测定蛋白酶水解产物或剩余底物的量来定量酶活性,常用“单位时间内产物生成量”或“底物消耗量”作为酶活单位。
基于天然蛋白质底物的检测以天然蛋白质(如酪蛋白)为底物,检测水解后产生的小分子产物以定量酶活。
• Folin-Ciocalteu法(Lowry法)
原理:蛋白酶水解酪蛋白后,生成含酪氨酸(Tyr)和色氨酸(Trp)的游离氨基酸或小肽。在碱性条件下,酚类氨基酸残基可还原Folin-Ciocalteu试剂(磷钼酸-磷钨酸混合物),形成蓝色的钼蓝/钨蓝复合物;颜色深度与产物浓度成正比。
操作:通过在680 nm测定反应产物吸光度进行定量。
优缺点:方法操作简便,定量范围为5–100 μg氨基酸;但特异性较弱,易受样品中其他还原性物质(如酚类、柠檬酸、含硫化合物)干扰,且结果受底物中Tyr与Trp含量影响。
• 茚三酮法(Ninhydrin Method):
原理:蛋白酶水解产生的游离氨基酸及肽中α-氨基在加热条件下与茚三酮反应,生成蓝紫色化合物(Ruhemann’s purple),其在570 nm处吸收最大。脯氨酸与羟脯氨酸则生成黄色产物(440 nm)。
优缺点:方法灵敏度高(检测限可达0.5 μg氨基酸),且适用范围广。但茚三酮试剂稳定性较差,不同氨基酸的显色效率存在差异,可能造成测量偏差。
• 邻苯二甲醛法(OPA Method)
原理:在硫醇(如β-巯基乙醇)存在下,OPA与伯胺(α-氨基和赖氨酸的ε-氨基)快速反应,生成高度荧光或在340 nm有强吸收的黄色异吲哚衍生物。
优缺点:OPA法比茚三酮法灵敏5-10倍,反应快速且在室温下进行。但其主要缺点是无法检测脯氨酸等仲胺,且与半胱氨酸的反应较弱或不稳定。反应产物不稳定,需要严格控制反应时间。
• 三硝基苯磺酸法(TNBS Method)
原理:TNBS在碱性条件下与伯胺反应,生成黄色的三硝基苯基-氨基衍生物,并可在420 nm处定量。
优缺点:灵敏度适中(0.05–0.4 μmol氨基酸),但反应时间较长;赖氨酸的ε-氨基也会参与反应,从而影响对水解程度的准确判断。此外,该方法不能检测脯氨酸。
为了提高检测的灵敏度和特异性,研究人员开发了多种基于化学修饰底物的检测。
• 偶氮酪蛋白法(Azocasein Assay)
原理:酪蛋白与对氨基苯磺酸重氮盐反应后生成红色偶氮酪蛋白。经蛋白酶水解,可产生可溶于三氯乙酸(TCA)的红色小肽段;未水解的偶氮酪蛋白则被TCA沉淀。离心后,上清液在440 nm处的红色吸光度与蛋白酶活性成正比。
优点:相较于Folin法,该方法背景干扰更少,操作简便,适用于常规蛋白酶活性测定。
• 合成显色/荧光底物法
原理:将显色基团(如对硝基苯胺,pNA)或荧光基团(如7-氨基-4-甲基香豆素,AMC)通过肽键连接到一段短的特异性肽序列上。当蛋白酶识别并切割该肽序列时,显色或荧光基团被释放出来,其浓度可以通过测定特定波长的吸光度(pNA在405 nm)或荧光强度来精确测量。
应用:这种方法的关键优势在于其高度的特异性。通过设计不同的肽序列,可以用于检测特定类型的蛋白酶。例如:此方法同样适用于外肽酶的检测,例如,通过合成Leu-pNA或Arg-AMC等底物来检测氨肽酶的活性。
-Tosyl-Gly-Pro-Arg-pNA:用于检测胰蛋白酶样蛋白酶。
-Suc-Ala-Ala-Pro-Phe-pNA:用于检测胰凝乳蛋白酶样蛋白酶。
-Suc-Ala-Ala-Pro-Val-pNA:用于检测弹性蛋白酶样蛋白酶。
3
高灵敏度检测技术:荧光共振能量转移
当需要检测超低浓度或微弱活性的蛋白酶时,基于荧光共振能量转移(FRET)的检测技术具有极高灵敏度,检测限可达ng级别。
通过荧光同质转移检测蛋白酶的原理
编辑
doi: 10.3389/fmicb.2023.1236368.
原理:FRET是一种发生在两个荧光生色团之间的非辐射能量转移过程。一个荧光团(供体,Donor)在被激发后,如果其发射光谱与另一个荧光团(受体,Acceptor)的吸收光谱有足够重叠,并且两者空间距离足够近(通常在1-10 nm),供体的能量就会转移给受体,导致供体荧光猝灭,而受体发射荧光(或受体本身是暗猝灭剂,导致整个体系荧光降低)。
在蛋白酶检测中,通常将供体和受体(或荧光团和猝灭剂)分别标记在一段特异性肽底物的两端。在底物完整时,FRET发生,荧光信号很弱。当蛋白酶切割肽链中间的识别位点后,供体和受体分离,FRET效应消失,供体荧光恢复。通过监测荧光信号的增强,可以实时、连续地测定蛋白酶活性。
• 同源转移荧光标记
同源转移荧光标记(Homotransfer Fluorescence Labeling)这是一类特殊的FRET应用,同一类荧光分子同时作为供体和受体。例如将高密度BODIPY染料标记到酪蛋白上。
完整蛋白中染料彼此距离近、发生自猝灭,荧光较弱;当蛋白酶将蛋白水解为小片段后,染料分散,猝灭解除,荧光显著增强。该方法(如ThermoFisher Scientific公司的EnzChek™蛋白酶检测试剂盒)相较传统FITC-酪蛋白法灵敏度提高约100倍,且操作更简便,无需分离步骤。
总之,从经典的平板筛选到现代的FRET技术,微生物蛋白酶的检测方法已经发展成为一个多层次、高精度的技术体系,为蛋白酶的发现、研究和应用提供了强有力的支持。
鉴于肠道蛋白酶失衡在多种胃肠道疾病中起关键作用,采取主动健康策略——通过监测症状、调整饮食与生活方式以维持肠道蛋白水解平衡——对预防和管理这些疾病至关重要。
1
症状监测与生物标志物评估
个体可通过留意特定胃肠道症状,初步判断是否可能存在蛋白酶活性异常。所谓“预警信号”与潜在生物标志物的变化紧密相关,建立“症状—标志物”的关联体系是精准干预的第一步。
• 慢性或复发性腹泻:该症状与粪便中升高的类胰蛋白酶活性(Tryptase-like activity)直接相关。研究显示,IBS-D及活动期UC患者粪便蛋白酶活性较健康人可高出数倍。这些蛋白酶通过激活PAR2受体,促进肠道氯离子分泌并引起水分丢失,从而导致腹泻。因此,腹泻的频率与严重程度可作为蛋白酶活性过高的临床指标。
• 腹痛和腹胀:持续的腹部不适或疼痛(尤其餐后加重)可归因于蛋白酶对感觉神经的直接激活。过量蛋白酶激活结肠传入神经上的PAR2受体,进一步敏化TRPV1等伤害感受器,降低痛觉阈值,进而引起内脏高敏感。腹胀则可能与蛋白质发酵过度产气有关,或与菌群失调(如产蛋白酶的毛螺菌科增多)导致的纤维发酵失衡相关。
• 食物不耐受与“肠漏”:对多种食物(尤其是蛋白质类)出现不良反应,常提示肠道屏障功能受损。生物标志物方面,可在粪便或血液中检测紧密连接蛋白(如ZO-1、Occludin)以及黏附连接蛋白(E-cadherin)的降解片段。粪肠球菌分泌的明胶酶GelE可直接降解E-cadherin,其活性水平可作为屏障破坏的直接评估指标;同时,粪便钙卫蛋白(Calprotectin)反映中性粒细胞相关炎症,其升高也间接表征由屏障破坏引发的黏膜炎症。
• 菌群特征监测:通过宏基因组测序重点追踪与蛋白酶活性相关菌群的丰度变化:如拟杆菌属(Bacteroides)、链球菌科(Streptococcaceae)和毛螺菌科(Lachnospiraceae)相对丰度升高,而具有保护作用的瘤胃菌科(Ruminococcaceae)比例下降,提示蛋白水解失衡的重要菌群特征。
当出现上述症状组合时,应及时就医,并考虑进行粪便蛋白酶活性、钙卫蛋白及菌群检测,以获得更全面的评估。
2
精准干预策略
基于监测结果,可以制定个性化的干预方案,旨在重建“蛋白水解平衡”(Proteolytic Balance)。
• 低FODMAP饮食:对IBS患者而言,短期(4–6周)低FODMAP饮食可通过减少肠内易被微生物快速发酵的底物,降低渗透压与产气,从而缓解腹胀和腹泻。其结果也可能减轻对肠道屏障的机械刺激,并改变产蛋白酶菌群的生长环境。
• 蛋白质摄入管理:对蛋白酶活性升高者,应适度控制总蛋白摄入,避免过量未消化蛋白进入结肠。优先选择易消化的优质蛋白(如乳清蛋白、鱼肉),并可考虑使用酶解蛋白,以减轻内源及微生物蛋白酶的消化负担。
• 益生元靶向补充:增加特定可溶性膳食纤维(如低聚半乳糖和抗性淀粉)可选择性促进双歧杆菌及产丁酸菌群(如瘤胃菌科)生长。丁酸盐不仅为结肠上皮细胞提供主要能量,增强屏障功能,还可通过降低肠腔pH抑制多种蛋白水解腐败菌。
• 补充产抑制剂的菌株:直接补充经临床验证可产生蛋白酶抑制剂的益生菌菌株,是极具前景的靶向策略。长双歧杆菌(Bifidobacterium longum)和短双歧杆菌(B.breve)产生的丝氨酸蛋白酶抑制剂(serpins)可中和肠道内过量有害丝氨酸蛋白酶,从而保护肠道屏障并减轻炎症。
• 补充产有益蛋白酶的菌株:布拉氏酵母(Saccharomyces boulardii)通过分泌一种蛋白酶来降解艰难梭菌的毒素A和B,是“以酶治酶”的典范。筛选和应用能够降解特定病理相关蛋白或毒素的益生菌,是未来的重要研究方向。
• 规避非甾体抗炎药(NSAIDs):非甾体抗炎药可抑制前列腺素合成,从而削弱黏膜的保护作用,增加对蛋白酶等攻击因素的易感性,进而诱发或加重黏膜损伤。有胃肠道风险的个体应严格避免使用或仅在医生指导下服用。
3
生活方式优化:建立“蛋白水解平衡”
长期的生活方式调节对于维持肠道稳态至关重要。
• 压力管理:慢性心理压力通过“肠-脑轴”影响肠道功能,可能增加肠道通透性、改变菌群组成,并影响蛋白酶的分泌与活性。通过冥想、瑜伽或认知行为疗法(CBT)等进行压力管理,有助于稳定肠-脑轴,从而间接维持蛋白水解平衡。
• 适度运动:规律的适度运动(如快走、慢跑)可改善肠道菌群多样性,增加有益菌丰度,并通过改善肠道血流和抗炎作用,有助于提升肠道通透性稳定性,从而维持屏障对蛋白酶的防御能力。
综上,主动健康管理提供了一个多维度、系统性的框架,通过整合症状监测、生物标志物评估、精准饮食、靶向微生态干预和生活方式优化,旨在从根本上恢复和维持肠道的蛋白水解平衡,从而有效预防和管理相关的胃肠道疾病。
微生物蛋白酶凭借其来源广泛、生化特性多样、生产成本相对较低以及易于基因改造等优势,在众多工业领域展现出巨大的应用价值和商业潜力,是现代生物技术不可或缺的组成部分。
蛋白酶的应用
编辑
微生物蛋白酶在各个工业领域的广泛应用,包括洗涤剂、皮革、食品、饮料、饲料及废物处理等。
1
工业应用
• 洗涤剂工业:碱性蛋白酶是现代洗衣粉和洗涤液的主要活性成分,正占据全球工业酶市场最大份额。它们可在碱性pH及不同温度条件下高效分解衣物蛋白质类污渍(如血渍、奶渍、草渍),提升洗净效果,并减少对环境有害化学助剂的依赖,契合绿色化学发展趋势。
• 皮革工业:传统皮革制造中,脱毛和软化多依赖硫化钠等高污染化学品。采用具备角蛋白酶与弹性蛋白酶活性的微生物碱性蛋白酶进行生物脱毛和软化,可有效去除毛发与杂质,使成品皮革更柔软、纹理更清晰,同时显著降低废水COD及硫化物污染,推动清洁生产。
2
食品与饮料
• 肉类嫩化与加工:中性或弱碱性蛋白酶(如来自芽孢杆菌的蛋白酶)被用于注射或浸泡肉类,通过水解肌肉纤维蛋白和结缔组织中的胶原蛋白,显著改善肉的嫩度。此外,蛋白酶还用于生产肉类水解蛋白和风味增强剂。
• 烘焙工业:在面包和饼干制作中添加中性蛋白酶,可对面筋蛋白进行适度水解,降低面团筋力并提升延展性与可操作性,从而得到体积更大、质地更均匀、口感更酥脆的产品。
• 乳制品工业:来源于米黑根毛霉(Rhizomucor miehei)等真菌的酸性蛋白酶,因其具有与小牛凝乳酶相似的凝乳活性,已成为奶酪生产中替代动物凝乳酶的主要选择,解决了来源有限和伦理问题。
• 酿酒与果汁澄清:在啤酒酿造中,添加脯氨酸特异性蛋白酶可降解造成“冷浑浊”的富含脯氨酸蛋白;在果汁生产中,酸性蛋白酶用于分解导致浑浊的蛋白-多酚复合物,从而提高澄清度与稳定性。
• 动物饲料工业:在动物饲料中添加外源性蛋白酶,可提高植物蛋白源(如豆粕)的消化利用率,降解抗营养因子,促进幼龄动物生长性能,同时降低粪便氮排放,减轻环境压力。
3
生物技术与特殊应用
在生命科学研究与生物制药中,高纯度重组蛋白酶是关键工具酶。凭借强序列特异性,TEV蛋白酶、SUMO蛋白酶和Kex2蛋白酶常用于在蛋白纯化后精确切除亲和标签(如His-tag、GST-tag)。
• 核酸提取中降解核酸酶
蛋白酶K(Proteinase K)具有广谱蛋白水解活性,并能在变性剂(如SDS)存在下保持活性,常用于核酸提取中降解核酸酶及其他蛋白污染物。
部分微生物蛋白酶因其温和、高效特性,也被应用于隐形眼镜清洁液中,有效去除镜片蛋白沉积,同时降低对眼睛的刺激。
蛋白酶是生命科学中功能高度多样的一类酶,其研究已从传统生物化学表征拓展到复杂系统生物学与疾病病理学。在胃肠道这一特殊生态系统中,宿主与微生物来源的蛋白酶共同构成精密的蛋白水解网络,维持消化吸收、黏膜屏障和免疫稳态至关重要。
但当肠道菌群失调时,该网络失衡:过量的微生物蛋白酶可通过激活PARs受体、直接降解屏障蛋白等途径,成为推动IBD、IBS等多种胃肠道疾病发生发展的关键因素。
深入解析这一复杂蛋白水解网络,不仅有助于揭示肠道疾病的深层病理生理机制,也为创新治疗策略的开发奠定了坚实基础。靶向特异性微生物蛋白酶、调控PARs信号通路,或通过微生态干预恢复蛋白水解平衡,正成为有前景的研发方向。与此同时,微生物蛋白酶凭借高效催化与生产优势,在食品、洗涤剂及环保等产业中持续创造显著的经济与社会价值,推动绿色生物制造与循环经济发展。
展望未来,随着基因组学、蛋白质组学(尤其是全面解析蛋白酶及其底物的“降解组学”degradomics)和合成生物学等前沿技术持续发展,我们有望发现更多功能新颖、特性独特的微生物蛋白酶。借助基因工程与定向进化,可实现“定制化”改造,满足医疗健康与绿色工业的特定需求。将基础研究的深刻洞见与应用技术的创新突破相结合,将更有效地驾驭蛋白酶这一强大的“分子剪刀”,服务于人类健康与可持续发展的宏伟目标。
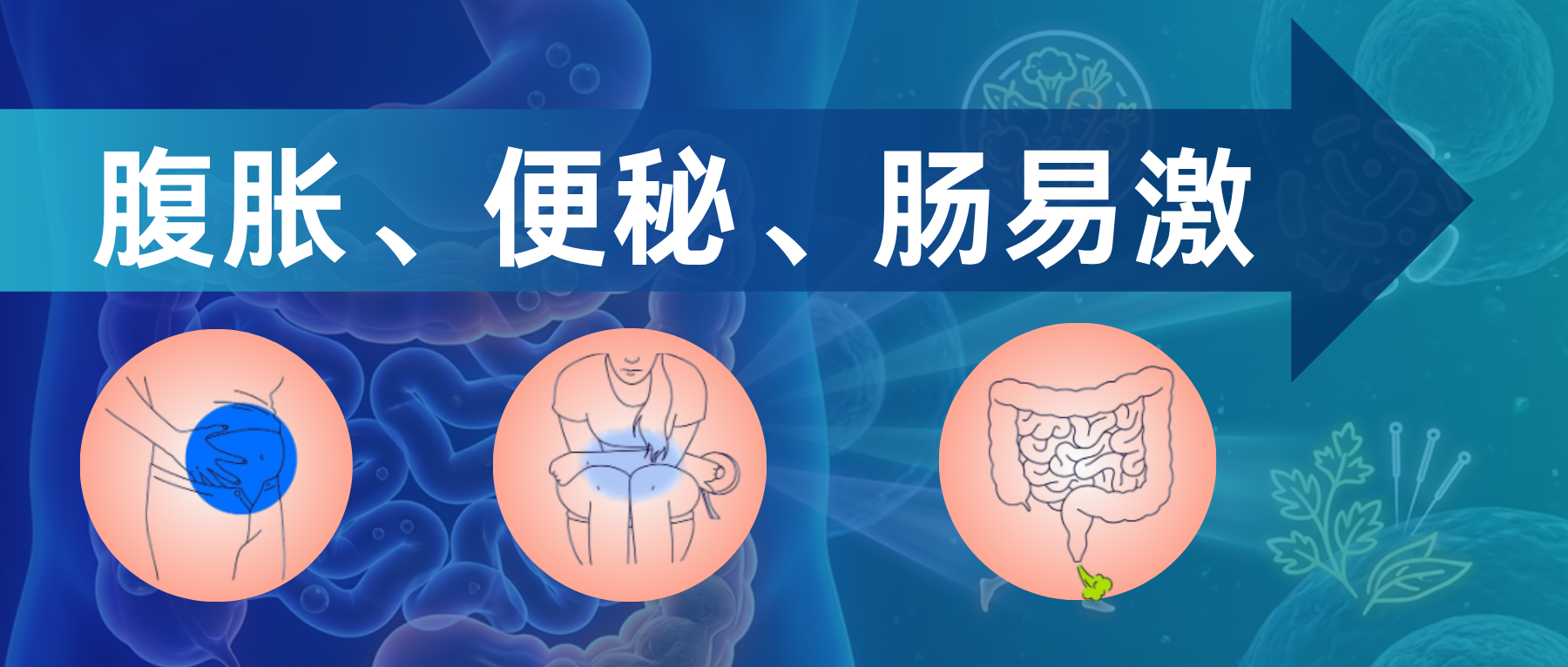
谷禾健康
编辑
在现代快节奏的生活中,消化系统不适已成为困扰全球数亿人的常见问题。其中,腹胀和便秘是最为普遍的胃肠道症状。它们不仅带来身体上的沉重感、疼痛和不适,更深刻影响着患者的心理健康和生活质量。许多患者在经历了长期的腹胀和排便困难后,最终被诊断为肠易激综合征(Irritable Bowel Syndrome, IBS),这是一种以腹痛伴随排便习惯改变为特征的功能性肠病。
过去,这类问题常被简单归因于“消化不良”或“胃肠动力不足”。随着神经胃肠病学和微生物组学的发展,人们逐渐认识到其背后是复杂的脑-肠-微生物轴失调。
研究表明,腹胀和便秘并非孤立的局部问题,而是涉及中枢神经系统、肠神经系统(ENS)、肠道内分泌激素、免疫炎症反应以及数以万亿计的肠道微生物的系统性失调。
本文将基于最新的科学研究和临床指南(如罗马IV标准),深入探讨腹胀与便秘的定义、高发人群、病理生理机制,特别是肠道菌群和肠道激素在其中扮演的关键角色,并为您提供一套涵盖饮食、微生态调节、生活方式及中医药的综合干预建议。
腹胀是一个常见但复杂的症状,原因多种多样,从饮食和压力到肠道微生物群失衡或医疗状况。本文解释了胃肠科医生通常会如何应对腹胀,胃肠专家会诊时应期待的情况,以及消化健康评估如何识别腹部不适背后的模式。帮助判断何时采取基础干预,何时需要进一步检查。
▸ 什么是腹胀和便秘?
▸ 腹胀的定义
腹胀是腹部胀气或膨胀感的统称。普通人吃饭后偶尔会有轻度腹胀,这是正常生理现象。只有频繁发作或持续存在的腹胀才需要关注。
▸ 便秘的定义
便秘主要表现为排便次数减少、粪便干硬及排便困难。一般以每周排便少于3次,并伴有排便费力或不畅作为诊断标准。
注:每个人排便习惯存在差异,有人每日1–2次,也有人2–3天一次。只要规律且粪便不干硬,均不属于便秘。
下面将从临床与医学角度进一步解析腹胀和便秘的定义及其亚型。
▸ 临床与医学定义
(主观感受与客观体征)
在消化系统临床中,腹胀是最常见且困扰患者的症状之一。医学上需区分“Bloating”和“Distension”两个概念。根据罗马IV标准及国际疾病分类(ICD-11),其区别主要如下:
▸主观腹胀(Bloating):指患者主观感受到的腹部饱胀感、紧绷感或气体堵胀感。这是一种感觉,可能伴随也可能不伴随腹部的实际增大。
▸客观腹部膨隆(Distension):指客观上可测量到的腹围增加,通常是肉眼可见的腹部向外突出。
二者可单独或同时存在。普通人群中,主观腹胀患病率约19%,伴客观腹围增加约9%;而在肠易激综合征(IBS)等功能性肠病患者中可高达66%–90%。
★ 腹胀是多系统的复杂失调
罗马IV标准将功能性腹胀和腹部膨隆定义为独立的“肠-脑互动异常”疾病。其诊断要求为:反复出现腹胀或腹部膨隆,平均每周至少1天,症状在诊断前已持续6个月,且近3个月符合标准,并排除IBS、功能性便秘等其他功能性胃肠病。该区分提示腹胀并非单一器质性问题,而是涉及神经、肌肉、微生物及免疫系统的复杂失调。
▸ 腹胀的亚型
腹胀可以分为功能性腹胀和器质性腹胀:
▸功能性腹胀:最常见,无明显的解剖学或生化异常。其发生多与内脏高敏感性、胃肠动力异常、肠道菌群失调及腹膈肌协同失调有关。
▸器质性腹胀:由明确的疾病引起,如肠梗阻、腹水(肝硬化、心衰引起)、胃轻瘫、乳糜泻、卵巢肿瘤等。在重症医学科中,腹胀甚至可能是危及生命的腹腔间隔室综合征(ACS)的先兆。
相比之下,功能性腹胀缺乏常规临床评估可解释的器质性原因。
▸ 腹胀与腹部肥胖的区别
许多患者,尤其是女性,常将功能性腹胀导致的腹围增大误认为“发胖”或“腹部肥胖”,但两者本质上截然不同:
▸基础原因不同:腹部肥胖是由于皮下脂肪或内脏脂肪的长期过度蓄积;而腹胀引起的腹围增大,多由肠道气体、滞留粪便或腹壁肌张力异常导致腹腔内容物前移所致。
▸时间节律不同:肥胖引起的腹围增大相对稳定,短时间内变化不明显;而功能性腹胀呈明显昼夜节律,常表现为“晨轻暮重”:晨起腹部平坦,随进食逐渐加重,至傍晚或夜间达峰,经休息或排气排便后缓解。
▸伴随症状不同:腹胀常伴有肠鸣音亢进、频繁嗳气、排气增多、腹痛或排便习惯改变(如便秘);而单纯的腹部肥胖通常不伴随这些消化道急性症状。
1
总体患病率与风险因素
腹胀和便秘是全球常见的健康问题。流行病学调查显示,全球约有15%-18%的普通人群每周至少经历一次腹胀。在中国,成人慢性便秘的患病率约为4.0%~10.0%,且随着年龄的增长而显著升高,80岁以上老年人的患病率甚至可达38.0%。
两者常相伴发生。在便秘型肠易激综合征(IBS-C)和功能性便秘(FC)患者中,超过90%伴有明显腹胀。
▸ 共同风险因素
这两种症状的共同风险因素包括:
–不良的饮食习惯(如高脂、低纤维饮食,高肉类摄入);
–缺乏体力活动或运动;
–不规律的排便习惯、
–精神心理压力(焦虑、抑郁、睡眠质量差);
–年龄、疾病因素等。
编辑
2
女性更易患腹胀和便秘
需要注意的是,腹胀和便秘的性别差异极为显著,女性患病率通常为男性的1.5~2倍以上。
女性之所以成为腹胀和便秘的高发群体,性激素(雌激素和孕激素)的周期性波动和生命阶段性衰退起到了决定性作用。特别是在围绝经期和围产期,女性的胃肠道面临着巨大的生理挑战。
▸ 围绝经期女性的“肠道危机”
围绝经期是指女性绝经前后的一段过渡时期,其核心特征是卵巢功能衰退,雌激素和孕激素水平剧烈波动并最终显著下降。研究表明,绝经后女性患便秘和IBS的症状严重程度显著高于绝经前女性。其机制主要包括:
1.胃肠动力减弱:雌激素受体(ERα、ERβ)及G蛋白偶联受体(GPER/GPR30)广泛存在于肠神经系统和肌间神经丛。雌激素通过一氧化氮(NO)通路调节平滑肌收缩与舒张;其水平下降会减慢胃肠蠕动、延迟胃排空并延长结肠传输时间,从而引发便秘和腹胀。
2.肠道微生态失衡:肠道菌群与雌激素相互调节。肠内“雌激素代谢菌群”通过分泌β-葡萄糖醛酸酶,将结合态雌激素转为游离态,促进肠肝循环重吸收。围绝经期雌激素下降会降低菌群多样性,减少有益菌、增加有害菌,既降低雌激素利用率,又增强发酵产气,加重腹胀。
3.内脏敏感性增加:雌激素对中枢神经系统的痛觉调节具有保护作用;其缺乏会降低内脏痛觉阈值,使正常肠气或轻度扩张被放大为明显的腹胀和腹痛。
▸ 围产期女性的“双重压迫”
在怀孕期间及产后(围产期),女性同样极易发生便秘和腹胀,患病率可高达40%-52%。其原因包括:
1.激素风暴:孕期孕激素显著升高,其平滑肌松弛作用虽可防止子宫过早收缩,但也降低胃肠道平滑肌张力、减弱肠蠕动,使食物残渣滞留时间延长、水分过度吸收,从而导致粪便干结和气体滞留。
2.机械性压迫:随着胎儿的生长,不断增大的子宫会直接压迫直肠和乙状结肠,形成物理性梗阻,阻碍粪便和气体的排出。
3.产后盆底肌受损:分娩(尤其阴道分娩)可损伤盆底肌及阴部神经,导致排便协同失调,排便时盆底肌反而收缩,进而引起排便困难和继发性腹胀。
1
饮食不当(最常见)
饮食不当是腹胀的首要原因,占比超过60%。具体常见情况主要有以下几种:
▸ 吃太多容易产气的食物
豆类、洋葱、大蒜、韭菜、萝卜、卷心菜、甘蓝这些食物含有较多不易消化的碳水化合物。它们进入结肠后被细菌发酵产生大量气体,易引发腹胀。
许多人饮奶后腹胀,多为乳糖不耐受所致,中国人约70%存在不同程度乳糖不耐受。
▸ 吃饭太快
吃饭速度快会吞入大量空气。这些空气进入胃肠道,加重腹胀感。边吃边说话更容易吞入空气。
▸ 吃太多高脂高糖食物
高脂肪高糖食物减慢胃排空速度,食物在胃内停留时间延长,发酵产气增加。
可能导致腹胀发生的因素
编辑
2
功能性消化不良
功能性消化不良占腹胀原因的15%~20%。这种病无明显胃肠结构异常,主要表现为餐后饱胀、早饱和上腹胀痛。
其发病与胃动力异常及内脏高敏感有关,约半数患者合并幽门螺杆菌感染,根除后部分症状可缓解。
3
本身便秘
便秘患者粪便长时间积聚在结肠内,细菌发酵粪便产生大量气体。便秘本身就会引发严重腹胀。很多便秘患者第一主诉就是腹胀。
4
肠易激综合征
肠易激综合征是一种常见功能性肠病。腹胀是肠易激综合征便秘型最常见症状,大约80%患者有不同程度腹胀。腹胀症状在排便后通常会减轻。
肠易激综合征中的微生物群改变
编辑
Almonajjed MB,et al.Medicina (Kaunas).2025
5
肠道菌群失调
抗生素使用、饮食改变和年龄增长可改变肠道菌群组成;产气菌过度生长会增加气体生成,导致持续腹胀。
▸ 肠道菌群失调会加重腹胀和消化功能障碍
肠道菌群失调还意味着微生物群落失衡。当产气细菌数量超过其他细菌,或有益菌数量被耗尽时,发酵平衡会倾斜。
肠道微生物群的失调会导致中枢和消化生理过程的改变,包括运动性和免疫功能,这些都会加重腹胀和消化功能障碍。
▸ 腹胀可能与菌群失调相关的迹象:
6
胃酸过低(低盐酸)
这里小编要特别强调,胃酸不足是传统医学中常被忽视的根本原因之一。胃酸(HCl)对蛋白质分解、食物杀菌、启动消化级联反应及维持酶活性所需酸性环境至关重要;其不足时,食物尤其蛋白质在进入小肠前难以充分消化。
▸ 低胃酸可致小肠细菌过度生长,从而引起腹胀
文献综述显示,胃酸不足时未消化食物在胃肠道发酵,可能诱发小肠细菌过度生长(SIBO),并促进幽门螺杆菌等进一步增殖。
SIBO与腹胀、吸收不良和腹泻有关,直接与胃酸减少及随后的上小肠菌群失衡相关。关于低盐碱和细菌过度生长的研究发现,在低酸条件下,小肠中的细菌过度生长会导致本应用于重要代谢功能的蛋白质分解,从而产生消化和系统下游的影响。
▸ 腹胀可能与胃酸过低有关的迹象:
7
肠道蠕动缓慢
肠道内存在“清洁运转周期”,对腹胀的发生至关重要。该专业名词为迁移运动复合体(MMC):MMC 是空腹期周期性反复出现的肠蠕动模式,可在餐间“清扫”残留食物残渣、细菌和其他残留物。
缺乏正常 MMC 活性与胃轻瘫、肠道假性阻塞及小肠细菌过度生长有关,从而引起显著腹胀。研究显示,慢传输型便秘患者常出现 MMC 异常(如缺失或延长周期,以及关键阶段蠕动减退)。MMC 受损时,食物停滞并发酵,平时会被清除的菌群得以定植和繁殖,进而导致腹胀、便秘和 SIBO。
▸ 腹胀可能与肠道蠕动缓慢有关的迹象:
8
小肠细菌过度生长(SIBO)
小肠细菌会分解碳水并生成气体及短链脂肪酸;当细菌过多并“吞食”时,气体与其他副产物就会显著增多。它们还会消耗蛋白质和维生素B12,以及脂肪消化所需的胆汁盐。
因此,小肠细菌过度生长(SIBO)导致的腹胀往往特别令人困惑:常见的“健康食物”反而会诱发发作。益生元纤维、洋葱、大蒜、苹果、豆类和发酵食品会为细菌提供养分;若这些细菌在小肠而非结肠内大量繁殖,就会在不合适的时间、地点与剂量下发生发酵,从而引起症状。
SIBO 往往并非独立疾病,而是多因素共同导致:胃酸受损使细菌得以在小肠存活,MMC 功能障碍无法有效清除,并叠加结构性问题形成“藏匿口袋”。
▸ 腹胀可能与 SIBO 相关的迹象:
9
消化酶不足
消化酶是分解食物的关键:淀粉酶处理碳水,蛋白酶处理蛋白质,脂肪酶负责脂肪。若酶分泌不足,食物会以较大且未充分消化的形式进入肠道,随后被细菌分解,从而引发腹胀。
▸ 最常见的是外分泌胰酶缺乏,导致脂肪吸收差
临床上最公认的情况是外分泌胰功能不全(EPI)。EPI的核心特征是外分泌胰酶缺乏,而脂肪吸收不良是最典型表现,常伴腹部不适和腹胀。
不过,腹胀不一定要达到“明显缺乏酶”的程度。亚临床功能不足(常与胃酸过低有关,因胃酸参与触发酶释放的信号级联反应)、压力或衰老,也可能导致碳水与脂肪分解不良。
▸ 腹胀可能与消化酶不足相关的迹象:
10
慢性压力与神经系统失调
这一点常被低估:若你曾在恐惧或压力时感到“胃部下沉”,甚至出现消化仿佛完全停止的体验,你可能已亲身感受到肠-脑轴。
▸ 压力或心情不好会影响食物消化
研究表明,压力会抑制迷走神经,进而损害胃肠道及微生物群,并参与肠易激综合征、炎症性肠病等胃肠疾病的发生发展。
肠易激综合征病理生理学要素
编辑
doi: 10.3390/jcm12072558.
迷走神经是肠道与大脑之间的重要通路。处于交感神经优势时,消化会被“忽略”:胃酸减少、酶分泌下降、肠蠕动减缓,且 MMC 节律受影响。清单中的这些环节都可能因长期且未缓解的压力而加重。
此外,迷走神经对肠道功能具有关键的中枢调控作用;压力可通过该通路直接影响消化动力、分泌与免疫功能。
▸ 腹胀可能与压力相关的迹象:
11
激素波动
这一原因仅限于女性,在功能和传统环境中都被严重低估。
▸ 月经周期可能会影响女性消化和吸收
一项关于孕酮与胃肠蠕动的综述指出:孕酮会减慢胃肠蠕动,并以“剂量依赖”方式影响胃排空,从而促成许多女性在黄体期出现消化迟缓和腹胀。
在月经周期中,卵巢激素的波动会调节胃肠道的收缩力、运输、分泌、内脏敏感性和免疫功能;月经期间雌激素水平下降可能尤其增加腹痛、改变排便习惯并引发腹胀。
此外,研究显示肠道通透性与月经周期存在关联:黄体期孕酮相关的变化可能影响肠道通透性、细菌易位以及乳房肿胀、腹胀等生理症状的严重程度。
▸ 临床实际情况:
▸ 腹胀可能与激素有关的迹象:
12
胃肠道器质性疾病
▸ 慢性胃炎、胃下垂
慢性胃炎患者胃动力减弱,食物排空减慢,容易腹胀。
胃下垂患者胃排空能力明显下降,站立后腹胀加重。
▸ 幽门梗阻
幽门是胃通向十二指肠的出口。溃疡或肿瘤造成幽门梗阻,食物和气体无法顺利通过,会引发上腹部严重腹胀。
▸ 肠梗阻
肿瘤、粪块、粘连性肠梗阻都能阻挡肠道内容物通过。梗阻近端肠道积气积液,腹胀非常明显,同时伴随停止排气排便。
▸ 腹腔肿瘤
卵巢癌、结肠癌、胃癌增长到一定体积,占据腹腔空间,产生持续腹胀。
13
全身性疾病
▸ 糖尿病
糖尿病患者常合并胃肠自主神经病变,胃排空减慢,肠蠕动减慢,引发腹胀便秘。
▸ 甲状腺功能减退
甲状腺激素分泌不足,全身代谢减慢,胃肠道蠕动也减慢,容易腹胀便秘。
▸ 慢性肾功能不全
尿毒症毒素影响胃肠道平滑肌功能,引发腹胀食欲下降。
▸ 心力衰竭
右心衰竭患者腹腔脏器淤血,胃肠道水肿,蠕动减慢,易出现腹胀食欲减退。
1
饮食结构不合理(最常见)
当代饮食越来越精细,膳食纤维普遍摄入不足。纤维能增加粪便体积、促进肠道蠕动;纤维不足则粪便量小、对肠道刺激不够,因而更容易便秘。
中国居民膳食纤维推荐摄入量是每天25~30克。实际调查显示,国人平均摄入量只有10~15克,远达不到推荐标准。
▸ 具体饮食相关因素:
– 吃太多精加工米面,全谷物摄入太少;
– 蔬菜水果吃得不够;
– 喝水太少,粪便水分被过度吸收,变得干硬;
– 吃太多肉类,膳食纤维严重不足。
可能导致便秘发生的因素
编辑
2
不良生活习惯
▸ 缺乏运动
长期久坐不动,肠道蠕动减慢。很多办公室工作者每天坐着超过8小时,便秘发生率明显高于体力劳动者。
▸ 排便习惯不良
很多人因为工作紧张,有便意时憋着不排便。长期反复这样,直肠对粪便刺激敏感性降低,便意减退,引发便秘。
▸ 如厕习惯不好
许多人上厕所玩手机,排便时间延长,容易诱发盆底肌功能紊乱,加重便秘。
▸ 长期精神紧张压力大
压力会通过脑肠轴影响肠道蠕动,进而影响便秘。
3
年龄增长
随着年龄增长,胃肠道蠕动减弱,腹肌与盆底肌力量下降,排便动力随之减弱,因此老年人便秘的发生率明显高于年轻人。60岁以上老年人的便秘发生率约为年轻人的2–3倍。
4
药物副作用
很多常用药物会引起便秘,这是临床便秘重要原因之一。
▸ 常见引起便秘的药物:
5
肠道菌群紊乱
越来越多研究证实,肠道菌群失调在便秘发病中起到重要作用。具体哪些细菌和便秘有关,我们后面专门详细说。
肠道微生物群可能影响便秘的发病机制
编辑
doi: 10.1093/gastro/goab035.
6
功能性疾病
▸ 肠易激综合征便秘型
肠易激综合征便秘型,除了便秘还伴随腹痛腹胀,排便后腹痛腹胀减轻。
▸ 结直肠肿瘤
肿瘤长大堵塞肠腔,造成排便困难。老年人突然发生便秘,一定要首先排除肿瘤可能。
▸ 直肠前突/盆底肌功能障碍
这是女性出口梗阻型便秘常见原因,多次生育女性更容易发生。
▸ 糖尿病
糖尿病神经病变影响肠道蠕动,很多糖尿病患者合并便秘。
▸ 甲状腺功能减退
甲状腺激素不足,全身代谢减慢,肠道蠕动也减慢。
▸ 神经系统疾病
中风、帕金森病、多发性硬化都能影响神经控制,引发便秘。
1
内脏高敏与腹膈肌协同失调
功能性腹胀的发生机制极其复杂,目前认为主要由以下几个核心环节构成:
▸ 内脏高敏感性
首先是内脏高敏感性。这是肠易激综合征(IBS) 和功能性腹胀的关键机制:患者的肠道会对正常生理刺激(如少量气体、食物残渣或正常蠕动)产生过度敏感。
其原因可能包括肠道黏膜的低度炎症、肠道通透性增加(“肠漏”),使潜在有害物质刺激黏膜下神经丛;信号经脊髓传入大脑后,被大脑皮层(如前扣带回、岛叶)错误地放大解读为胀痛感。
▸ 腹膈肌协同失调
其次是腹膈肌协同失调,它解释了为什么腹胀患者的腹围会明显增大。
健康情况下,肠道内容物增多(气体或食物)会触发协调反射:膈肌向上松弛、腹壁肌肉收缩,从而增加腹腔容积,避免腹部向前突出。
但在腹胀患者中,该反射出现异常:进食或肠腔轻度扩张时,膈肌反而出现矛盾性收缩(向下移动),同时前腹壁肌肉异常松弛。这种内脏-躯体反射失调会使腹腔内容物被向下、向前挤压,从而造成可见的腹部膨隆。
2
腹胀与便秘同行的疾病机理与致病因素
腹胀与便秘常常是“难兄难弟”,两者同发的概率很高。在慢传输型便秘和便秘型肠易激综合征(IBS-C)中,二者病理机制相互促进,形成恶性循环。
▸ 粪便滞留与过度发酵
当结肠蠕动减弱(慢传输)或排便出口受阻(排便障碍)时,粪便在肠内停留时间变长。水分被过度吸收使粪便干硬;更关键的是,滞留的食物残渣为肠道菌群提供底物,促使细菌长时间发酵,产生大量气体(如氢气、甲烷、二氧化碳)。
▸ 肠腔扩张与动力抑制
气体增多与粪便堆积会造成肠腔扩张。一方面直接引起腹部胀满;另一方面可触发抑制性反射,进一步减弱肠蠕动,加重便秘。
此外,部分气体(如甲烷)还可能具有神经肌肉毒性,直接抑制肠道平滑肌收缩,减慢肠道传输。
3
肠道激素与神经递质在胃肠动力中的作用
胃肠道不仅负责消化,也被认为是人体最大的内分泌器官。肠道激素与神经递质共同调控胃肠动力、感觉和分泌,起到“指挥官”的作用。
肠道微生物群对神经递质的调节
编辑
•血清素(5-羟色胺):人体约95%的血清素(5-HT)存在于胃肠道,主要由肠嗜铬细胞(EC细胞)合成并释放。5-HT是“脑-肠轴”连接的重要关键分子:当食物或气体刺激肠壁时,EC细胞释放5-HT,作用于肠神经系统的5-HT3与5-HT4受体。
5-HT4受体激活可促进乙酰胆碱释放,增强肠蠕动与分泌;5-HT3受体则参与疼痛信号向中枢传递。便秘患者常见5-HT通路异常(如合成减少、受体表达下调),导致肠道动力不足;而IBS患者则多见异常释放,引发内脏高敏与腹痛。
•胃动素:由小肠M细胞分泌,主要在空腹(消化间期)释放;它激活MMC的第III相,也就是“肠道清道夫”运动,将未消化的食物残渣和细菌推向大肠。若胃动素分泌不足或受体不敏感,便会导致胃排空延迟和小肠细菌过度生长(SIBO),进而引起腹胀。
•胆囊收缩素:主要由小肠I细胞分泌。进食脂肪和蛋白质后,CCK释放增多,促进胆囊收缩并刺激胰酶分泌;同时它显著抑制胃排空、减慢结肠传输。IBS患者可能对CCK更敏感,从而出现进食后夸张的动力抑制与腹胀。
•多巴胺:尽管多巴胺常被视为中枢“快乐递质”,但肠道同样可合成大量多巴胺。肠道多巴胺受体(如D2受体)被激活时,通常会抑制胃肠动力。因此,帕金森病患者往往伴随顽固便秘和腹胀(中枢多巴胺不足,同时外周受体敏感性可能发生改变)。
4
肠道产气机制及其对症状的放大效应
正常成年人每天肠道排气量约为500-1500毫升。肠道气体主要来源于吞咽的空气和肠道细菌的发酵。未被小肠完全吸收的碳水化合物(如膳食纤维、低聚糖、抗性淀粉)进入结肠后,被肠道菌群发酵,产生氢气(H2)、二氧化碳(CO2)以及微量的甲烷(CH4)和硫化氢(H2S)。
编辑
不同菌群代谢产生的气体对肠道功能影响迥异。例如,甲烷(CH4)不仅是一种温室气体,在人体内它被证明是一种神经递质调节剂。
产甲烷菌(如史氏甲烷短杆菌Methanobrevibacter smithii)消耗氢气生成甲烷。高浓度的甲烷能够直接抑制回肠和结肠的平滑肌收缩,显著延长肠道传输时间。临床研究证实,呼气试验中甲烷阳性的患者,其便秘的严重程度和腹胀频率远高于阴性患者。此外,硫化氢(H2S)虽然量少,但具有高度毒性,不仅产生恶臭,还可能损害肠道上皮细胞的能量代谢,加剧内脏高敏。
近年来研究已证实:肠道菌群在便秘与腹胀的发生发展中起关键作用。
便秘患者粪便菌群α多样性通常降低,厚壁菌门/拟杆菌门比例升高。
1
产气菌过度生长易导致腹胀
产气菌过度生长会使肠道内产生的气体增多,从而引发腹胀等不适症状。
–Klebsiella pneumoniae(肺炎克雷伯菌)
克雷伯菌过度生长会产生大量气体,引发明显腹胀。很多小肠细菌过度生长患者,克雷伯菌丰度显著升高。
–Streptococcus属某些物种
链球菌发酵碳水化合物产生大量气体,增加腹胀风险。
-Enterobacteriaceae科整体
肠杆菌科多数细菌发酵碳水化合物产气,丰度升高后气体产生增加。
人类肠道微生物组中主要的产气微生物
编辑
doi.org/10.1016/j.jff.2022.105367
2
为什么产气菌多会腹胀便秘?
产气菌过度增殖会消耗可发酵碳水化合物,生成大量氢气、二氧化碳和甲烷;气体积聚引起腹胀。与此同时,甲烷可减慢肠蠕动、延长结肠传输时间,从而加重便秘,因此不少便秘患者常同时出现腹胀。
细菌发酵中H2、H2S、CH4和CO2的生化途径
编辑
doi.org/10.1016/j.jff.2022.105367
3
哪些细菌增多易导致腹胀?
–Desulfovibrio物种
硫酸盐还原菌代谢产生硫化氢,高浓度硫化氢对肠道平滑肌有毒性作用,抑制蠕动。研究发现,便秘患者硫酸盐还原菌丰度高于健康人。
-黏液降解菌过度生长
Akkermansia muciniphila(嗜黏蛋白阿克曼菌)专门降解肠道黏液层。适量阿克曼菌对健康有益,过度生长会破坏黏液屏障,引发肠道炎症,影响蠕动功能。
微生物群和肠道屏障完整性
编辑
4
哪些细菌减少易导致便秘腹胀?
-产丁酸菌减少
丁酸盐是结肠上皮细胞主要能量来源,同时能够促进肠道蠕动,维持肠道正常节律。
主要产丁酸细菌包括:Faecalibacterium prausnitzii(普拉梭菌)、Roseburia属物种(如Roseburia inulinivorans)、Eubacterium rectale(直肠真杆菌)、Butyricicoccus pullicaecorum。
这些细菌丰度降低,丁酸产生减少,结肠上皮能量不足,蠕动功能减弱,容易便秘。
–Bifidobacterium(双歧杆菌)减少
双歧杆菌能够调节肠道蠕动,降低肠道pH值,促进有益菌生长。很多研究证实,便秘患者双歧杆菌丰度显著低于健康人。
–Lactobacillus(乳杆菌)某些物种减少
Lactobacillus reuteri(罗伊氏乳杆菌)能够改善肠道动力,减少炎症反应。补充罗伊氏乳杆菌能够缩短结肠传输时间。干酪乳杆菌也能调节肠道运动功能,增加排便频率。
近年来多个大型队列研究证实了肠道菌群和便秘腹胀的关系。随机双盲安慰剂对照试验证实:
补充 Bifidobacterium lactis能够增加每周排便频率,改善粪便性状;
补充Lactobacillus reuteri 缩短结肠传输时间15-20%;
益生元菊粉能够增加粪便中双歧杆菌数量,增加排便频率。
212种肠道微生物群与便秘的因果关联
编辑
doi: 10.3389/fmicb.2024.1438778.
5
针对特定细菌的干预方法
根据肠道菌群检测结果,不同菌群异常采用不同干预策略。
▸ 产气菌过度生长(克雷伯菌、肠杆菌科丰度升高)
饮食干预—减少精制碳水化合物摄入:产气菌喜欢利用容易发酵的简单糖产生气体。减少精制糖就是减少底物。
增加膳食纤维,特别是不可发酵纤维:不可发酵纤维能够促进肠蠕动,减少细菌过度生长。不可发酵纤维主要存在于全谷物外壳、蔬菜的纤维素部分。
低FODMAP饮食:低FODMAP饮食减少可发酵寡糖和单糖,减少气体产生。对小肠细菌过度生长引发的腹胀效果很好。
需要注意,低FODMAP饮食不建议长期使用,一般使用4-6周,症状缓解后需要逐步恢复高纤维饮食,避免长期低纤维饮食对菌群造成不良影响。
益生菌:补充产丁酸益生菌可通过竞争占位抑制产气菌生长;如布拉氏酵母菌、嗜酸乳杆菌等还能减少有害菌过度繁殖,改善肠道菌群环境。
抗生素:确诊为小肠细菌过度生长后,可在医生指导下短期使用抗生素(如利福昔明),通常疗程约1–2周;不建议自行用药,也不推荐长期使用抗生素。
▸ 产丁酸菌减少
增加膳食纤维摄入:尤其是菊粉、低聚果糖等益生元,可选择性促进产丁酸菌生长。常见富含益生元的食物包括:菊苣、芦笋、洋葱、大蒜、香蕉、燕麦。
多吃全谷物:全谷物因膳食纤维更丰富,可促进产丁酸菌生长;抗性淀粉同样能促进其增殖,例如冷却后的米饭和青香蕉都富含抗性淀粉。
直接补充:可以补充含有产丁酸菌的益生菌产品;或者补充丁酸栓剂或丁酸衍生物,但是注意直肠局部生产的丁酸效果更好。
▸ 双歧杆菌、乳杆菌减少
多吃含有低聚糖的食物:比如大豆、洋葱,能够促进双歧杆菌生长。
发酵乳制品:比如酸奶,含有活的乳酸菌,能够直接补充,注意选择无糖原味酸奶,避免额外添加过多糖分
直接补充:直接口服含有双歧杆菌或乳杆菌的益生菌制剂;选择明确标注菌株的产品,不同菌株功能不同。比如罗伊氏乳杆菌DSM17938明确研究证实对便秘有效,购买时可以关注菌株编号。
▸ Parabacteroides distasonis 低丰度
目前没有专门针对性食物。研究发现,蘑菇多糖能够促进P. distasonis 生长。克罗恩病患者食用蘑菇提取物后,P. distasonis 丰度升高,症状改善。可以尝试增加蘑菇摄入,每天吃100-200克新鲜蘑菇,有助于促进生长。
胃肠科医生在评估腹胀时通常会先进行细致而全面的检查:明确症状特点,识别风险,找出最可能引发不适的机制。尽管不同医生的具体方案会有所不同,但常见流程可能包括:
⑴ 询问临床病史
起效时间、发作频率及具体时段(如饭后、早晚),并评估可能诱因。需记录饮食习惯、纤维摄入、碳酸饮料,以及乳糖或果糖的暴露情况,和近期抗生素使用史。
抗生素使用可能产生的关联
编辑
同时梳理排便习惯(便秘、腹泻、腹痛急迫、排便不尽)及伴随症状(烧心、恶心、打嗝、疼痛),是否受月经周期影响,体重变化、疲劳与压力。
药物和补充剂使用情况(如二甲双胍、阿卡博糖、GLP-1 受体激动剂、阿片类药物、制酸剂、益生菌)。
⑵ 体格检查
进行腹部体检,观察是否存在压痛、腹胀、肠鸣音异常,以及器官肥大或肿块;同时评估是否有液体积聚(腹水)或疝气迹象。有时还会进行直肠检查,以了解粪便负担、张力及盆底相关问题。
⑶ 腹部不适的诊断与风险评估
症状出现的年龄、家族史(乳糜泻、炎症性肠病、结直肠癌)、旅行史,以及近期感染或手术史(如胆囊切除术后可能出现胆汁酸相关腹泻)。
⑷ 初步测试(如有指示)
基础化验常包括:全血细胞计数、电解质检测、炎症标志物(如C反应蛋白)、肠道菌群检测、甲状腺功能及乳糜泻相关血清学。
必要时可做粪便检测,如粪便钙卫蛋白用于评估炎症,或粪便病原检测以排除感染。呼气测试可评估乳糖不耐受或果糖吸收不良;而针对小肠细菌过度生长(SIBO),有时会采用乳果糖或葡萄糖呼气测试。
⑸ 询问临床病史
若出现警报症状或检查结果需要进一步评估,可能会进行上消化道内镜(EGD)、结肠镜、腹部超声或其他影像学检查,以排查结构性或炎症性疾病;怀疑胃轻瘫时,可考虑做胃排空检查。
通过整合这些数据点,胃肠科医生会制定计划,可能包括饮食调整、肠道习惯优化、微生物调节策略或治疗特定疾病的方案。重要的是,这很少是一个放之四海而皆准的流程。相反,它是迭代的,并由你个人的反应随时间引导。
!
重点:为什么这个诊疗对肠道健康很重要
腹胀不仅仅是烦恼,它常提示消化系统失衡。对部分人而言,可能是快速发酵的碳水化合物刺激了敏感肠道;对另一些人,缓慢排便与便秘会导致气体积聚;还有人则可能与微生物失衡或炎症有关。
由于肠道与免疫、新陈代谢、神经系统和微生物群紧密相连,理解腹胀有助于打开更广泛的消化健康视角。
积极处理持续腹胀可缓解不适,提升生活质量、精力与社交功能,也能避免反复试验带来的“走弯路”,减少误诊或延误评估的风险,防止症状在无意中加重。
若腹胀伴随非自愿体重减轻、便血、持续呕吐、吞咽困难、发热、贫血,或50岁后新出现的症状等警示信号,应尽早重视并进行及时、详细的评估。
▸ 腹胀并非单独出现,常见伴随症状包括:
长期且失控的腹胀会显著降低生活质量,并因饮食受限导致营养不良,进一步加剧肠—脑反馈循环。腹胀本身虽只是症状而非疾病,但可能提示肠易激综合征、功能性腹胀/膨胀、小肠细菌过度生长(SIBO)、乳糜泻、碳水化合物不耐受、胆汁酸吸收不良,或在特定情况下提示更严重的潜在病理。因此,当症状持续时,进行结构化评估并咨询胃肠专科尤为重要。
1
个体对腹胀原因的差异与不确定性
▸ 症状相似但可能由个性化的不同机制引起
每个人的消化系统都不完全相同。遗传、早期暴露、饮食、压力、激素、药物以及微生物群都会影响肠道对日常食物的反应:同样一顿饭,有人舒服,有人却胀气。
即使症状看起来相似,也可能由不同机制引起。例如,夜间腹胀的两个人可能都表现出相近外观:其中一人可能结肠排出缓慢、以产甲烷的菌群为主;另一人则可能主要由压力和快速发酵的碳水化合物导致内脏过敏。
2
仅根据症状本身难以确定根本原因
▸ 症状相似但可能由于不同病因
腹部腹胀表面上可能很相似:无论成因是气体过度产生、气体处理能力受损、肌肉协调障碍、炎症、酶缺乏还是微生物群失衡,都会出现类似表现。
肠易激综合征、功能性消化不良、乳糖不耐受和小肠细菌过度生长(SIBO)等多种疾病中也常有重叠症状。缺少具体背景时,同样的症状可能对应不同病因,甚至导致误诊。
案例一:许多人误以为所有腹胀都源于豆类和十字花科蔬菜,因此完全回避。但真正的主要诱因往往是严重便秘。相较于减少健康食物,改善排便频率与规律更能缓解腹胀。
案例二:有人会把腹胀归咎于麸质。检测已排除乳糜泻;反而,果聚糖和多元醇可能是个体的主要触发因素。通过结构化的重新引入,可更清楚界定耐受水平,从而保留更多饮食多样性。
案例三:有人在使用抗生素后反复腹胀与腹泻,怀疑乳糖不耐受;但呼气测试及症状出现时间提示,更可能是感染后IBS,且可能伴随SIBO。治疗重点应放在促进胃肠动力与调整微生物平衡。
这些例子说明了清晰诊断框架的价值。它减少了不必要的限制,将精力集中在最重要的地方,并为个性化策略打开了大门——尤其是在结合测试和细致跟进的客观见解时。
鉴于腹胀与便秘发病机制复杂,单一治疗常难以见效。现代医学通常建议采用多层次、多靶点的综合干预,包括饮食调整、微生态治疗、生活方式干预以及中西医结合治疗。
1
饮食干预:低FODMAP饮食与膳食纤维的精准应用
▸ 低FODMAP饮食法是针对腹胀的有效干预
对伴有明显腹胀的 IBS 患者而言,低 FODMAP 饮食是目前国际公认的一线干预方案。FODMAP 指“可发酵的寡糖、二糖、单糖和多元醇”。这类短链碳水在小肠中难以充分吸收,进入结肠后易被细菌快速发酵产气,同时具高渗透性,把水分吸入肠腔,从而引起腹胀并加重腹泻。
低FODMAP饮食分为三个阶段:
1.排除期(2-6周):严格去除所有高FODMAP食物(如小麦、洋葱、大蒜、苹果、西瓜、牛奶、豆类及人工甜味剂),以迅速缓解症状并建立消化基线。
2.重新引入期(6-8周):症状控制后,对各类FODMAP食物进行逐一、少量重引,评估耐受情况,明确具体“触发食物”。
3.个性化维持期:依据测试结果制定长期饮食方案,仅避免已证实不耐受的食物,以兼顾营养均衡与肠道菌群多样性。
▸ 膳食纤维的精准补充
膳食纤维是预防和治疗便秘的关键,但若患者同时伴有腹胀,补充需格外谨慎。不可溶性纤维(如麦麸)虽能增加粪便体积,却可能带来较强摩擦且发酵快,容易加重腹胀;因此更推荐可溶性、非黏性或低发酵性纤维。
例如,猕猴桃(每天2个)已在多项临床试验中证实可显著提高排便频率、软化粪便,并因其特异性酶(如猕猴桃碱)和低发酵特性,通常不易引起腹胀,患者满意度高。
西梅(每天100g)富含山梨糖醇和多酚,具有天然的渗透性通便作用;此外,部分水解瓜尔豆胶(PHGG)和洋车前子壳粉也属优选纤维补充剂,可形成凝胶润滑肠道,产气量较低。
2
微生态调节:益生菌与益生元的循证应用
纠正肠道微生态失衡是治本之策。微生态制剂(益生菌、益生元、合生元)通过竞争性抑制有害菌、修复肠黏膜屏障、调节免疫和神经递质分泌,有效改善腹胀和便秘。
▸ 精准益生菌株的选择
益生菌的效果具有显著的菌株特异性。对IBS相关腹胀与腹痛,长双歧杆菌35624®(Bifidobacterium longum 35624)和格氏乳杆菌LA806循证证实可显著降低IBS严重程度评分(IBS-SSS),缓解内脏高敏。
而对慢性便秘,乳双歧杆菌HN019和动物双歧杆菌BB-12可缩短肠道传输时间并提高排便频率。
▸ 益生元协同作用
益生元(如低聚果糖、菊粉)是益生菌的“食物”。对便秘患者而言,合生元(益生菌+益生元)的联合使用常可产生“1+1>2”的效果。需注意的是,部分益生元含FODMAP,摄入过量可能在早期加重腹胀,应从小剂量开始,逐步增加。
3
生活方式与心理行为干预
生活方式的调整与良好的精神状态,不仅是治疗方案的起点,也是所有治疗能够有效开展的基础。
▸ 日常保持一定的运动
避免久坐:每坐1小时起身活动2-3分钟,走动并做轻度伸展即可促进肠蠕动。
规律运动有助于改善便秘。每周约150分钟的中等强度有氧运动(如快走、慢跑)可机械性刺激胃肠蠕动、缩短结肠传输时间。太极拳与瑜伽等特定运动(如膝胸动作)配合腹式呼吸与腹部内脏按摩,既能促进排气,也有助于调节自主神经、降低交感兴奋,缓解压力。
推荐运动项目:
-快走或慢跑:每天30分钟,促进全身血液循环,增加肠蠕动;
-瑜伽:很多扭转体式能够按摩腹部肠道,促进蠕动;
-游泳:全身运动,促进代谢,改善肠道功能;
-腹肌锻炼:适当锻炼腹肌,增加排便动力,老年人特别需要注意腹肌锻炼。
▸ 压力管理
长期压力增大,通过脑肠轴抑制肠道蠕动。你可以尝试:
-每天10-15分钟深呼吸冥想;
– 规律作息,保证每天7-8小时睡眠;
– 找到适合自己的放松方式,比如听音乐、养花、散步。
▸ 环境调整
-旅行环境改变容易便秘,提前做好准备,带够膳食纤维食物,不要完全改变饮食习惯。
–长期精神紧张工作环境,试着拆分任务减少压力,定时起来活动。
▸ 心里干预
鉴于“脑-肠轴”的紧密关联,心理干预对难治性腹胀和便秘尤为关键。认知行为疗法(CBT)、肠道导向催眠疗法和正念冥想可降低焦虑与抑郁水平,纠正大脑对内脏痛觉信号的异常处理,从而从根本上缓解症状。
4
中医药干预:中草药、针灸与内外同治
中医药在治疗功能性胃肠病方面具有独特的“整体观念”和“辨证论治”优势,强调多靶点、双向调节。
▸ 中草药的独特机制
大黄:大黄是治疗便秘的中医经典药。现代研究认为其具有“双向调节”机制:蒽醌类成分(如番泻苷)在肠道菌群作用下转化为大黄酸,可刺激肠神经丛,促进肠液分泌与蠕动,从而通便泻下;而其中的鞣质类成分具收敛作用,可抑制肠道水通道蛋白(AQPs)表达,起到止泻作用。凭借这种动态平衡,大黄在腹泻-便秘交替型IBS的治疗中更具优势。
生姜与薄荷油:生姜含姜辣素和姜烯酚,具有促胃动力、抗炎与抗氧化作用,可加速胃排空,缓解上腹胀和恶心。薄荷油(主要成分薄荷醇)可作为天然的平滑肌钙离子通道阻滞剂,解除肠道痉挛,缓解IBS患者的腹痛和腹胀,因而被多国指南推荐为一线用药。
经典方剂:如白术、白芍、防风、陈皮用于肝郁脾虚型IBS,能抑肝扶脾、解痉止痛;木香、枳壳、乌药则能行气导滞,显著改善便秘型IBS患者的腹胀和排便困难。
▸ 中医外治法:针灸与穴位贴敷
针灸刺激特定穴位(如天枢、足三里、大肠俞、上巨虚),可调节自主神经平衡,促使胃肠激素(如胃动素、VIP)分泌趋于正常,实现肠道动力的双向调节。
穴位贴敷(如用生姜泥或健脾温肾中药贴敷神阙穴/肚脐)则借助药物透皮吸收与经络传导直达病所。临床研究显示,生姜热敷脐部可有效预防并缓解卧床患者(如急性心肌梗死者)的腹胀与便秘,且无全身性副作用。
1
微生物组检测在腹胀便秘背景下能揭示的信息
对于频繁或持续腹胀的人群,粪便微生物组检测报告可提供如下参考:
• 微生物多样性: 慢性胃肠症状患者有时会观察到较低的多样性,但这并非腹胀特有。这可以指导逐渐谨慎地重新引入不同纤维。
• 相对丰度: 甲烷相关信号升高可能与便秘为主的模式相符。某些有益类群(如双歧杆菌)含量偏低,可能反映了饮食、抗生素史或其他因素。
• 发酵潜力: 分类模式可能指示其碳水化合物发酵能力极强;如果与敏感性结合,这有助于塑造饮食试验。
• 潜在失衡的标志: 部分特征显示其易产生炎症或硫化物,因此应密切关注诱因和耐受性。
微生物组检测多以粪便样本为主,用于表征微生物多样性及细菌(有时也包括其他生物)的相对丰度。不同方法差异明显:不少检测采用16S rRNA基因测序分析细菌类群;也有部分采用散弹式宏基因组学,捕捉更广泛的微生物并推断功能通路。结果可能呈现多样性指标、特定群体潜在过度生长,以及与胀气、便秘或腹泻等症状相关的模式(往往为松散关联)。
在16S检测方面,数据库更庞大、模型构建时可纳入的因素更多,因而敏感性与稳定性通常更强。宏基因组则能识别真菌、毒力基因及部分代谢相关基因,从而更全面了解微生物并推断其功能。
需要注意的是,微生物组检测不能用于诊断SIBO或排除其他医疗状况;但当症状持续、且需要反复调整管理方案时,它可帮助进行个体化规划。
2
哪些时候可以考虑微生物组检测?
并非所有腹胀患者都需要做微生物组检测。很多人通过日常调整并在必要时进行简短的临床评估即可改善。但若符合以下情况,补充相关信息可能会更有帮助:
在这些情况下,结合临床评估与个性化肠道微生物组报告的仔细审查,有助于指导有针对性的营养选择和微生物支持策略。目标不是“治疗检测结果”,而是将数据与症状、饮食和生活方式整合成一个连贯的计划。
3
决策支持:微生物组检测何时有意义?
当信息能改变您下一步的行动时,测试最有价值。
• 持续的不确定性: 你尝试过第一线策略(比如慢吃、减少碳酸饮料、优化排便、基础碳水化合物-乳糖试验),但都没有明确的答案。
• 反应不一: 有些改变能稍微改善,但症状会反复出现;目前尚不清楚是否应该增加纤维、更换纤维类型,还是继续避免。
• 便秘为主症状: 甲烷相关模式的证据支持关注运输时间和特定微生物方法。
• 抗生素后或感染后转变: 了解多样性和相对丰度有助于重新引入阶段并支持恢复。
• 复杂的饮食或限制: 客观的洞察有助于战略性地重建饮食多样性,避免不必要的回避。
相反,如果你的腹胀是轻微的,明显与暴饮暴食或汽水有关,并且通过基本调整就能缓解,检测效果可能不会太大。同样,如果出现明显症状,医疗评估优先于任何择期检测。
4
胃肠专科就诊咨询的帮助
胃肠专家咨询将对您的症状模式与可能诱发机制进行评估和对应。典型的就诊和随访可能包括:
• 明确目标:哪些结果最重要——是减少胀气、减少不适日、更明确的诱因,还是改善肠道规律?
• 症状日记/饮食记录: 在预约前后进行简短记录有助于识别时间模式(例如,与每日进食节律或便秘相关的下午-傍晚胀痛)。
• 选择性检测:如前所述,可能会根据风险、病史和检查安排针对性的化验或微生物测试,呼气测试等。
• 管理试验:指导性饮食调整(例如,FODMAP 轻量法并重新引入)、肠道优化(补水、可溶性纤维或渗透剂治疗便秘),或在适当情况下支持肠道蠕动。
• 微生物组及辅助见解:可选的微生物组检测可用于制定下一步,尤其是在反应部分或非典型时。
• 后续与迭代: 在规定的间隔后回顾进展,根据反应调整,避免长时间不必要的限制。
这一流程强调协作与灵活调整,而不只是追逐症状。它要把机制理清:气体来源可能与肠黏膜受损有关,饮食可能导致发酵增加并影响肠道蠕动,微生物失衡则可能与炎症或结构性问题相关。明确这些因素后,腹胀治疗方案便能更有针对性地选择。
5
管理腹胀的循证策略
管理与干预通常不依赖单一因素。相反,结合符合个体生理特点与偏好的多项调整往往效果更佳。潜在组成包括:
• 饮食行为:吃得更慢;彻底咀嚼;均匀间隔的用餐;限制碳酸饮料的使用;避免过量口香糖或硬糖,以免增加吸入空气。
• 靶向饮食试验:短暂、结构化的低 FODMAP 阶段,随后逐步重新引入以确定个人耐受性;如果呼气测试或病史支持不耐受,则要求乳糖或果糖限制;在工艺早期关注糖醇(山梨醇、甘露醇)和高度可发酵的纤维。
• 纤维:如果便秘,应优先选择可溶性纤维(例如部分水解的瓜尔胶、车前子)和充足的水分摄入。高不可溶性纤维可能会加重某些人的症状。
• 粪便调节:对于慢性便秘,建议在临床医生指导下使用循证渗透剂,必要时进行盆底评估,并定期活动以支持肠道蠕动。
• 肠道-大脑支持:减压、膈肌呼吸以及认知行为或肠道导向催眠疗法可以降低内脏敏感度,改善部分患者的症状控制。
• 其他:薄荷油肠涂胶囊有一定证据可能缓解与IBS相关的不适。西甲硅油在某些情况下或可帮助减轻胀气。益生菌可作为试验性选择,但效果会因菌株和个体差异而不同;最好结合临床与微生物组背景来选择合适的菌株。
• 医疗治疗:如果怀疑并确诊 SIBO,有时会与促进肠道蠕动的同时使用靶向抗生素或草药方案;如果发现胆汁酸腹泻,胆汁酸结合剂可能有帮助;如果存在胃排空延迟,可以考虑促动力学策略。
需要强调的是,这并非“一刀切”治疗。关键在于把治疗选择与驱动你症状的机制对齐,并持续、系统地重新评估。
腹胀与便秘,看似是日常生活中微不足道的“小毛病”,实则是人体内部复杂生态系统失衡的警报。从肠道内数以万亿计的微生物,到穿梭于神经突触间的5-羟色胺;从女性生命周期中起伏的雌激素,到内脏高敏、腹膈肌协同失调,从肠道激素的紊乱,再到产甲烷菌等微生态的失衡,每一个环节都相互交织,互为因果。大脑与肠道之间无声的对话,每一个环节的微小偏差都可能演变为令人痛苦的临床症状。
面对这种复杂的疾病网络,仅靠单一的泻药或促动力药物往往只能“治标”,且长期使用可能带来依赖与副作用。未来治疗方向应走向更精准、更综合。
通过低FODMAP饮食减少发酵底物,结合特定益生菌株与益生元重塑微生态;再辅以运动与心理干预,打破脑-肠恶性循环。同时充分发挥中医药多靶点、双向调节的优势,才能真正从“缓解症状”迈向“恢复稳态”的根本转变。
随着微生物组学测序技术普及以及人工智能在医疗数据分析中的应用,我们有望实现真正的精准医疗:基于个人肠道菌群图谱、基因型与代谢特征,制定专属的益生菌组合、饮食方案和药物干预策略。
对广大受困扰的患者,尤其处于特殊生理阶段的女性而言,理解疾病机制是战胜疾病的第一步。保持良好心态,科学调整饮食,合理补充膳食纤维并选择合适的益生菌;必要时及时咨询消化科与心理科医生。通过重建肠道微生态与脑-肠轴的平衡,才能重新获得轻松顺畅的健康生活。
注:本账号内容仅供学习和交流,不构成任何形式的医疗建议。
主要参考文献
中华医学会消化病学分会胃肠动力学组,功能性胃肠病协作组,食管疾病协作组.中国慢性便秘临床诊断与治疗规范[J].中华消化杂志,2024, 44(10): 649-658. DOI:10.3760/cma.j.cn311367-20240930-00373.
Napolitano M, Fasulo E, Ungaro F, Massimino L, Sinagra E, Danese S, Mandarino FV. Gut Dysbiosis in Irritable Bowel Syndrome: A Narrative Review on Correlation with Disease Subtypes and Novel Therapeutic Implications. Microorganisms. 2023 Sep 22;11(10):2369.
Zhang S, Wang R, Li D, Zhao L, Zhu L. Role of gut microbiota in functional constipation. Gastroenterol Rep (Oxf). 2021 Aug 6;9(5):392-401.
Almonajjed MB, Wardeh M, Atlagh A, Ismaiel A, Popa SL, Rusu F, Dumitrascu DL. Impact of Microbiota on Irritable Bowel Syndrome Pathogenesis and Management: A Narrative Review. Medicina (Kaunas). 2025 Jan 13;61(1):109.
Feng C, Gao G, Wu K and Weng X (2024) Causal relationship between gut microbiota and constipation: a bidirectional Mendelian randomization study. Front. Microbiol. 15:1438778.
Shaikh SD, Sun N, Canakis A, Park WY, Weber HC. Irritable Bowel Syndrome and the Gut Microbiome: A Comprehensive Review. J Clin Med. 2023 Mar 28;12(7):2558.
Mamieva Z, Poluektova E, Svistushkin V, Sobolev V, Shifrin O, Guarner F, Ivashkin V. Antibiotics, gut microbiota, and irritable bowel syndrome: What are the relations? World J Gastroenterol. 2022 Mar 28;28(12):1204-1219.
Bercik P. The Brain-Gut-Microbiome Axis and Irritable Bowel Syndrome. Gastroenterol Hepatol (N Y). 2020 Jun;16(6):322-324.
Ohkusa T, Koido S, Nishikawa Y, Sato N. Gut Microbiota and Chronic Constipation: A Review and Update. Front Med (Lausanne). 2019 Feb 12;6:19.
谷禾健康
近年来,越来越多的人开始关注口腔健康。但实际上,很多口腔问题早已在悄悄增加。无论是儿童常见的龋齿,成年人困扰的口臭,还是中老年人高发的牙周病,其发生率都在不断上升。其中,牙周炎因患病人数众多、进展隐匿,已被认为是一个重要的公共卫生问题。
更值得关注的是,口腔健康并不仅仅关乎牙齿本身。过去十多年的研究发现,口腔微生物与全身健康之间存在着密切联系。例如,牙周炎与糖尿病、心血管疾病、类风湿性关节炎,甚至阿尔茨海默病等慢性疾病之间,都存在一定关联。
在口腔微生物组(上)篇中,我们探讨了口腔微生物组的复杂性,口腔菌群的定植与平衡(口腔微生物组指南(上):认识菌群定植、共生与失调,守护全身健康),接下来这篇,我们将深入了解口腔微生物组如何与多种系统性疾病相互作用,揭示其潜在机制和临床意义。通过分析口腔微生物组的失调如何影响心血管疾病、糖尿病、认知衰退等健康问题,我们将为口腔健康管理提供新的视角和干预策略。
龋 齿
为什么爱吃糖会把菌斑推向致病状态
► 龋齿的本质与成因
龋齿,俗称“蛀牙”,本质上并不是某一种细菌单独造成的结果,而是一种由微生物介导、受饮食驱动的牙体硬组织进行性破坏。它的关键,在于牙菌斑生物膜的生态平衡是否被打破。
在健康状态下,牙面菌群与宿主处于动态平衡之中。但当口腔长期受到可发酵碳水化合物,尤其是蔗糖的反复冲击时,这种平衡就会开始偏移。
根据“生态学假说”,龋病并非由单一病原体引起,而是整个龈上微生物组在持续糖压力下发生生态演替的结果。
► 生态平衡的破坏与致龋循环
糖被摄入后,菌斑中的产酸菌会迅速代谢底物,产生大量乳酸,使局部pH值下降。若这种过程反复发生,菌斑环境就会长期处于酸化状态。当pH持续低于约5.5时,对酸敏感的共生菌受到抑制,而更耐酸、也更善于产酸的菌群则被逐渐富集。于是,菌群结构开始从相对稳定的共生状态,转向更有利于致龋的失调状态。
在这一过程中,变形链球菌(Streptococcus mutans)和乳杆菌(Lactobacillus)是典型代表。它们既能产酸,又能耐受酸性环境,因此在低pH条件下更容易获得优势,并进一步强化局部酸化,形成一个自我维持的“致龋恶性循环”。
龈上牙菌斑的不同形态
Kim et al., 2020
► 龋齿的复杂性
其中, S. mutans之所以重要,不只是因为它产酸能力强,还因为它能够利用蔗糖合成大量胞外多糖(EPS)。
EPS作为生物膜基质,一方面促进细菌在牙面的黏附、聚集和生物膜成熟,另一方面形成物理屏障,削弱唾液的缓冲作用,使酸性产物更容易在生物膜深层积聚。结果是,牙釉质长期暴露于低pH微环境中,脱矿逐渐发生,最终形成龋损。
► 细菌、真菌的协同作用
近年来的研究还提示,龋病并不完全是细菌。尤其在严重早期儿童龋中,白色念珠菌(Candida albicans)也可能参与致病过程。它与 S. mutans 之间存在明显的跨界协同: 白色念珠菌的菌丝可为 S. mutans 提供附着支架,而 S. mutans 产生的EPS又能将 C. albicans 稳定嵌入生物膜。两者共同存在时,生物膜的结构稳定性和产酸能力都会增强,从而加重牙体组织破坏。
因此,更准确地说,龋齿是口腔微生物组由共生走向失调的典型结果。当菌斑生态被持续推向一个更酸、更黏、更利于致龋菌生存的方向时,龋病就开始发生了。
牙周病
牙龈发炎、牙槽骨吸收,菌群是怎样失控的
牙周病是一类影响牙齿支持组织的慢性炎症性疾病,主要包括牙龈炎和牙周炎。它的起点并不复杂:龈下菌斑生物膜发生失调。但问题在于,这种失调一旦越过某个阈值,后果就不简单。
牙龈炎是牙周病的早期阶段,也是相对可逆的阶段。它通常由菌斑堆积引发,表现为牙龈红肿、出血。这个阶段,微生物组的整体变化还不算剧烈,但已经开始出现向革兰氏阴性菌倾斜的趋势。如果炎症长期得不到控制,病变就会向深部组织推进,逐渐发展为牙周炎。
► 牙周炎的生态变化与关键病原体(牙龈卟啉单胞菌等)
牙周炎的关键变化,是生态位变了。随着牙周袋形成,龈下环境变得更加缺氧,同时龈沟液渗出增加,局部富含蛋白质、血红素和宿主组织降解产物。这种环境不再适合原本占优势的健康相关菌,反而更有利于严格厌氧、蛋白水解型细菌扩张。
因此,牙周炎是一个多物种协同驱动的失调过程。其中,牙龈卟啉单胞菌(Porphyromonas gingivalis)常被视为关键病原体。它能够通过牙龈蛋白酶等毒力因子破坏宿主免疫监视、操纵炎症反应,为整个致病菌群创造更有利的生存条件。
与此同时,福赛斯坦纳菌(Tannerella forsythia)和齿垢密螺旋体(Treponema denticola)也常与其共同富集,被视为重度牙周炎的核心相关菌群。
► 牙周炎的复杂性与恶性循环
除了这些重要菌,其他厌氧菌也在其中发挥协同作用。比如,具核梭杆菌(Fusobacterium nucleatum)常被称为“桥梁菌”,因为它能够连接早期和晚期定植菌,促进更复杂的生物膜形成。与此同时,Prevotella intermedia、Parvimonas micra、Filifactor alocis等菌也常参与其中,共同塑造出一个更稳定、也更具破坏性的龈下微生态。
► 牙周炎:多样性增加
和龋病不同,牙周炎的微生物变化并不主要表现为少数菌独占优势,而常常表现为多样性增加。这提示牙周袋已经形成了一个更复杂的生态系统:营养来源更多,微环境分层更明显,不同细菌之间的协作也更强。
换句话说,牙周炎不是简单的感染加重,而是一个失调群落逐步成熟的过程。
► 失调菌群与宿主炎症反应相互放大——造成组织破坏
这些菌群通过LPS、蛋白酶和其他代谢产物持续刺激宿主,诱导一种过度而破坏性的炎症反应。最终,胶原纤维被降解,牙槽骨发生吸收,牙齿支持组织逐渐丧失,临床上就表现为牙周袋加深、附着丧失、牙齿松动,甚至脱落。
所以,牙周病更像是口腔微生物组在龈下环境中完成了一次方向错误的重组:菌群越失调,炎症越难控制;炎症越持续,越有利于致病菌继续扩张。牙周炎的进展,正是这种恶性循环的结果。
口腔念珠菌病
当共生真菌变成机会致病菌
口腔念珠菌病,俗称“鹅口疮”,是最常见的口腔真菌感染,主要由白色念珠菌(Candida albicans)过度生长所致。它的特殊之处在于:白色念珠菌本来就是口腔中的常见共生菌。问题在于什么条件下,它会从共生状态转向致病状态。
► 这种转变通常发生在宿主防御被削弱的时候
常见诱因包括免疫力下降、长期使用广谱抗生素、佩戴义齿以及唾液分泌减少。这些因素看似不同,但共同指向一点:它们都会改变口腔原有的微生态平衡,使原本受到限制的念珠菌获得扩张机会。
►口腔念珠菌病的发生,同样嵌在微生物之间的相互作用之中
抗生素的使用就是典型例子。它在抑制细菌感染的同时,也会杀灭那些原本对白色念珠菌具有抑制作用的细菌,从而为真菌过度生长腾出空间。
更复杂的是,口腔中的细菌对念珠菌并不都是拮抗的。一些细菌会与其形成协同关系。例如,口腔链球菌(Streptococcus oralis)可增强白色念珠菌的黏附、生物膜形成和对宿主组织的侵袭能力。
相反,也有一些共生菌具有保护作用。比如乳酸杆菌(Lactobacillus)能够通过产生乳酸、过氧化氢和细菌素等物质,抑制白色念珠菌的生长及其菌丝形成,在维持细菌—真菌平衡中发挥重要作用。
► 从致病机制看,白色念珠菌的关键是毒力状态的切换
其致病性主要依赖于几个环节:
► 白色念珠菌转为致病菌,出现白斑
当这些过程被启动后,白色念珠菌就不再只是“定植者”,而成为能够黏附、侵入并损伤上皮的机会致病者。临床上,最终表现为口腔黏膜上的典型白色斑块。
因此,口腔念珠菌病是一次真菌—细菌—宿主三者失衡。当抑制力量减弱、协同因素增强,而白色念珠菌又完成了从共生型向侵袭型的切换,疾病就发生了。
牙周袋:一个容易被忽视的炎症和细菌储源
如果说龋病和牙周病首先是口腔局部的微生态失衡,那么更值得警惕的是:这种失衡并不总停留在口腔内部。
尤其在牙周炎中,牙周袋可以被视为一个持续存在的炎症和细菌储源。在这个环境里,失调的龈下微生物群长期定植,局部炎症不断维持,牙龈上皮屏障也随之受损,甚至形成溃疡面。这样一来,口腔与全身之间原本相对清晰的边界,就变得不再稳固。
编辑
这意味着,牙周袋更像是一个持续向外释放细菌、细菌产物和炎症介质的源头。日常生活中,咀嚼、刷牙等看似普通的动作,都可能造成短暂菌血症,使口腔中的微生物或其成分进入血液循环。
因此,牙周炎的意义不能只理解为牙龈发炎或牙槽骨吸收。从全身角度看,它还代表着一种持续存在的炎症。也正因为如此,口腔健康与全身健康之间的关系,是可能存在明确的生物学通路。
细菌、毒素和炎症介质,如何从口腔影响全身
口腔微生物组失调之所以可能影响全身,主要有三条路径:细菌本身进入循环、细菌产物进入循环,以及局部炎症介质外溢。
编辑
路径一,细菌本身进入循环
牙周炎时,受损的牙龈屏障为细菌进入血流提供了直接通道。
研究已经在动脉粥样硬化斑块、关节滑液,甚至阿尔茨海默病患者的大脑中,检测到牙周相关致病菌的DNA或菌体本身,如Porphyromonas gingivalis 和 Fusobacterium nucleatum。
这并不意味着口腔细菌必然是这些疾病的单一病因,但至少说明,它们并非始终被限制在口腔局部。
路径二,细菌产物进入循环
细菌结构成分和代谢产物的系统性作用,其中最典型的是革兰氏阴性菌外膜上的脂多糖(LPS)。
LPS进入血液后,可与宿主细胞表面的Toll样受体4(TLR4)结合,激活NF-κB等炎症信号通路,诱导TNF-α、IL-6、IL-1β等促炎细胞因子产生,从而推动或维持一种低度的全身炎症状态。
口腔源LPS驱动的代谢性内毒素血症,被认为是连接牙周炎与胰岛素抵抗、动脉粥样硬化等代谢性疾病的重要机制之一。
路径三,局部炎症介质溢出到全身循环
牙周炎病灶中会持续产生大量TNF-α、IL-6、IL-1β、PGE₂和MMPs。这些分子不仅参与局部组织破坏,也可能进入循环,对远端器官产生影响。
比如,升高的TNF-α和IL-6可干扰肝脏和脂肪组织的胰岛素信号传导,促进胰岛素抵抗;同样,这类炎症因子也会加重血管内皮功能障碍,促进动脉粥样硬化斑块形成与不稳定。
从这个意义上说,牙周炎并不是一个封闭在口腔里的局部事件。它更像是一个持续释放炎症信号的慢性病灶。
为什么口腔炎症和全身慢病常常互相加重
口腔与全身之间的关系,关键不只在于口腔炎症影响全身,还在于它们之间往往是双向作用。
◐ 全身慢性病 👉 口腔
一方面,糖尿病、肥胖、心血管疾病等慢性病常伴随着持续的低度系统性炎症。这种全身炎症状态会改变口腔局部环境,对口腔微生物组施加新的选择压力,促进失调发生。
在2型糖尿病患者中,高血糖可导致中性粒细胞趋化和吞噬能力下降,削弱机体对牙菌斑细菌的清除能力;同时,AGEs积累又会过度激活巨噬细胞,放大TNF-α、IL-1β等促炎反应,使局部组织破坏更重、更难恢复。
◑ 口腔 👉 全身慢性病
另一方面,系统性炎症还会改变龈沟液和唾液成分。牙龈血管通透性增加后,龈沟液流量上升,其中富含血清蛋白、炎症介质和宿主降解产物,为蛋白水解型牙周致病菌提供了更有利的营养条件。
对糖尿病患者而言,唾液和龈沟液中的葡萄糖浓度升高,又进一步为糖酵解细菌提供额外底物,推动致龋菌和部分牙周相关菌扩张。
恶性循环
慢性炎症还常伴随氧化应激增强。过量活性氧不仅会损伤牙周组织,也会进一步改变微生物生存环境。结果是,原本已经失衡的口腔微生物组更容易向致病方向偏移,而失调菌群又反过来加剧牙周炎、龋病等口腔疾病的严重程度。
于是就形成了一个典型的恶性循环:
全身慢病所伴随的低度炎症 → 改变口腔局部环境 → 促进口腔微生物组失调 → 加重口腔炎症 → 进一步推高全身炎症负担。
这也解释了为什么对一些慢性病患者来说,口腔问题往往不是孤立出现的。它既是局部失衡的结果,也可能是全身炎症状态的一部分表现。
越来越多研究提示,口腔微生物组并不是孤立的局部生态。尽管不同疾病的证据强度并不一致,但它与代谢、免疫、炎症以及神经退行性过程之间,可能存在真实的生物学联系。
编辑
1.糖尿病
被高血糖重塑的口腔微生物组
在口腔微生物组与全身疾病的联系中,2型糖尿病(T2D)与牙周炎是目前证据较充分。
菌群为什么会失衡?因为生态位先变了
糖尿病并不会凭空制造某一种致病菌,而是先改变了菌群赖以生存的生态位。高血糖状态下,唾液和龈沟液中的葡萄糖浓度升高,唾液pH下降,龈沟液渗出增多,局部血清蛋白和组织降解产物也更容易进入牙周袋。对微生物而言,这意味着营养底物更丰富、炎症相关分子更多、局部环境更有利于耐炎症和机会致病菌扩张,导致口腔菌群往往会逐渐偏离共生状态,滑向更具炎症驱动性的失调状态。
菌群结构变化
系统综述显示,与健康对照相比,2型糖尿病患者口腔微生物组中常可见厚壁菌门富集,部分研究还观察到梭杆菌门(Fusobacteria)增多,而拟杆菌门在健康人群中更常见。
属水平上,Prevotella、Catonella、Leptotrichia、Rothia 等菌属更常被报道为富集。另一些研究则提示,2型糖尿病患者中还可能出现变形菌门增加,而梭杆菌门和弯曲杆菌门相对减少。
虽然不同研究结果不完全一致,但糖尿病导致的改变反映了菌群结构和功能整体向促炎方向偏移。
菌群功能的重组
关于菌群α多样性的研究结果也并不统一:有的研究发现升高,有的发现降低。这恰恰说明,关键不在于“菌更多还是更少”,而在于菌群之间的关系、代谢分工和生态功能被重新组织了。
已有研究提示,血糖控制较差时,龈下产丁酸菌减少;而糖尿病相关代谢紊乱可伴随琥珀酸积累,进一步增强Fusobacterium nucleatum、Porphyromonas gingivalis 等牙周相关菌的生长优势和毒力表达。
炎症本身也在塑造菌群
糖尿病相关的菌群失调,并不只是高糖“喂出来”的结果,炎症本身也是力量。研究显示,糖尿病状态下,中性粒细胞趋化、吞噬和杀菌功能受损,巨噬细胞更倾向于维持促炎表型,T细胞反应也更偏向Th1/Th17而非免疫调节状态。结果是,机体一方面对菌斑清除不足,另一方面局部炎症又持续放大,最终形成“清除不够、炎症过强、修复不足”的失衡状态。
动物实验发现,糖尿病易感小鼠在疾病进展过程中会出现口腔菌群明显重塑;而阻断 IL-17 通路后,糖尿病背景下口腔菌群的致病性可以被部分削弱。
也就是说,炎症并不只是菌群失调的后果,它反过来也在推动菌群向更具致病性的方向演化。
2型糖尿病与牙周炎之间的双向循环关系
编辑
doi: 10.1080/20002297.2026.2635233
微生物通路与全身健康
口腔微生物组不仅因为它影响牙龈和牙周组织,还可能通过代谢通路参与更广泛的全身调控。
口腔菌群参与的硝酸盐–亚硝酸盐–一氧化氮(NO)通路逐渐受到关注。部分口腔细菌能够将唾液中的硝酸盐还原为亚硝酸盐,后者进一步参与体内NO生成。NO与血管舒张、葡萄糖代谢和免疫调节密切相关,因此这条通路可能是连接口腔健康与心脏代谢健康的重要桥梁。
已有小型实验研究发现,干扰口腔硝酸盐还原菌会影响血压和代谢反应;ORIGINS研究也提示,反映更强亚硝酸盐生成能力的微生物标志,与更低的心代谢风险评分相关。
这意味着,牙周炎相关的菌群失调,可能不仅加重局部炎症,也可能削弱NO相关代谢通路。
牙周炎对糖代谢的影响
牙周病中的细菌、脂多糖(LPS)及炎症介质可持续进入循环,推动低度全身炎症,干扰胰岛素信号,并促进胰岛素抵抗。
纵向研究和荟萃分析提示,牙周炎与新发糖尿病风险升高相关;
一项荟萃分析显示,牙周炎与新发糖尿病风险增加相关,风险比约为1.26(95% CI 1.12–1.41)。
ORIGINS研究进一步将龈下菌群组成与炎症水平、胰岛素抵抗、前驱糖尿病患病率及后续血糖变化联系起来,表明口腔菌群失调可能不仅是糖尿病的结果,也可能参与糖代谢恶化。
治疗牙周,也可能帮助控糖
随机对照试验、系统综述和Cochrane综述显示,牙周治疗后 3–4个月,糖尿病患者的 HbA1c平均可下降约0.43%。
这一变化幅度虽然不算巨大,但已具有明确临床意义,也说明控制牙周炎可能减轻代谢负担。
综上所述,高血糖持续改变口腔生态位,推动菌群结构、代谢功能和宿主反应失衡;失调菌群又通过局部炎症、系统炎症、NO通路受损及潜在的口腔—肠道轴效应,反过来参与糖代谢恶化。因此,牙周管理应被纳入整体代谢管理的一部分。
2.肥胖
口腔微生物组重塑加剧牙周炎风险
肥胖除了与肠道微生物组的稳定关联外,越来越多的证据显示,肥胖状态也伴随口腔微生物组的系统性重塑,并可能通过炎症与代谢通路与牙周炎形成相互促进的双向关系。
肥胖相关的脂肪组织炎症可导致循环中TNF-α、IL-6、CRP等炎症介质升高,诱发或加重胰岛素抵抗。同时,脂肪因子谱系失衡(如瘦素升高、脂联素下降)可影响中性粒细胞与巨噬细胞功能,改变组织修复与免疫稳态,使牙周组织更易处于促炎与破坏性微环境,为牙周相关致病菌的定植与扩张提供生态位基础。
慢性牙周炎与肥胖之间的双向联系机制
编辑
doi: 10.1016/j.jds.2025.07.009
门水平的偏移
研究显示,肥胖个体的唾液中常观察到厚壁菌门相对富集,而拟杆菌门在非肥胖个体中更占优势。这一现象与部分2型糖尿病研究的结果相似,提示口腔微生物组可能对“能量过剩—低度炎症—代谢紊乱”背景存在共同响应模式。这类门水平差异反映了群落生态结构的整体偏移,提示肥胖状态下口腔微生态稳态更易被打破,并向促炎相关的结构迁移。
属水平变化:从“饮食底物信号”到“牙周致病潜能”
在属水平上,肥胖个体唾液中显著富集的菌属包括:
肥胖相关口腔菌群改变,同时包含营养底物改变、免疫炎症偏移与致病潜能上调等多个层面。
肥胖与牙周致病菌群:即便无临床牙周炎,也可能预先定植
除了上述菌属,一些与牙周病相关的“菌团复合体”细菌也与肥胖相关,并且在部分研究中,这种关联可在统计上独立于临床牙周炎的存在。值得注意的是,传统上与重度牙周炎密切相关的核心病原组合(“红色复合体”)包括:
这一现象提示肥胖可能通过系统性炎症与免疫调节改变,先行改变口腔/牙周生态位,使这些牙周相关病原体更易稳定存在,随后在菌斑积累或局部屏障受损等条件下进一步推动牙周组织破坏。
表明肥胖不仅是牙周炎的背景风险因素,也可能通过驱动牙周致病菌群的结构性偏移参与疾病进程。
致肥胖微生物组
致肥胖微生物组(obesogenic microbiota)的讨论框架同样适用于口腔。肥胖的慢性低度炎症与脂肪因子失衡背景下,口腔微生物组更易发生促炎与牙周致病方向的生态重组。这种重组可能通过增强局部与系统炎症、加重胰岛素抵抗相关通路,反过来参与肥胖相关代谢风险与牙周并发症的形成。
因此,在肥胖人群中评估与干预口腔微生态(尤其是龈下/唾液菌群及牙周状态),可能具有超越口腔局部的代谢与炎症管理意义。
3.肝病
牙周炎加重肝纤维化,口腔微生态影响肝功能
近年来,越来越多临床研究支持“口腔—肠道—肝轴”的存在,即牙周炎相关口腔失衡可通过全身炎症、内毒素血症及口源菌经肠道转运等途径影响肝脏。
现有系统综述纳入的 20 项研究总体提示:口腔炎症与肝损伤之间具有生物学合理性,但因果关系尚未建立,且不同研究在样本来源、检测方法和肝病表型上存在较大异质性。
MASLD/NAFLD:以牙龈卟啉单胞菌为代表的牙周致病菌最受关注
在各类肝病中,关于 MASLD/NAFLD 的证据最多。现有研究显示,牙周炎暴露,尤其是牙龈卟啉单胞菌,与 NAFLD 更重的肝脏表型、特别是肝纤维化进展相关。
部分研究发现,唾液中牙龈卟啉单胞菌检出率在 NAFLD 患者中更高,其侵袭性 fimA 基因型与病情进展有关;另有研究提示,牙龈卟啉单胞菌升高与肝脏硬度增加及进展性纤维化风险升高相关。
除牙龈卟啉单胞菌外,Aggregatibacter actinomycetemcomitans、Prevotella intermedia、Tannerella forsythia、Fusobacterium nucleatum也被反复提及。
其中特别是 A. actinomycetemcomitans 与更差的代谢/肝脏表型(如胰岛素抵抗、脂肪肝程度)有关,而 NAFLD 相关口腔失衡中还可见 Porphyromonas endodontalis、Prevotella intermedia、Fusobacterium nucleatum增多。不过,需要注意的是,并非所有研究都支持单一病原体模型。
大样本人群研究显示,血清抗牙龈卟啉单胞菌抗体与 NAFLD 并无明确关联;相反,较新的 16S 研究更支持群落水平信号,即与 MASLD 相关的并非单一菌,而是以 Neisseria、Streptococcus、Fusobacterium、Veillonella 为优势的唾液微生物群结构变化。
口腔-肠道-肝脏轴将牙周炎与非酒精性脂肪性肝病联系起来
编辑
doi: 10.2147/IJGM.S554833
其他肝病类型:病毒性肝炎、肝硬化、酒精性肝炎和 HCC 亦可见口腔微生态异常
除脂肪性肝病外,病毒性肝炎和肝硬化患者也常伴有更重的牙周负担。
研究提示,在 HBV/HCV 相关肝病中,牙周炎与肝纤维化进展存在关联;肝硬化患者口腔中“红复合体”相关菌群,即牙龈卟啉单胞菌、Tannerella forsythia 和 Treponema denticola,可相对增多。另一方面,也有研究指出,肝硬化合并牙周炎患者的龈下菌群并不一定表现为经典红/橙复合体富集,而更像是免疫功能受损背景下的非典型失衡。
在 AAH 中,患者针对牙龈卟啉单胞菌的IgG/IgA/IgM 水平升高,且 IgM 反应与肝损伤指标相关。
在 HCC 尤其是 MASH/NASH-HCC 中,现有小样本研究提示 Fusobacterium nucleatum富集最为稳定,而牙龈卟啉单胞菌、Tannerella forsythia、Prevotella intermedia 也有升高趋势。
另有研究发现,HBV→肝硬化→HCC 进展过程中,唾液菌群多样性下降,并伴随分期相关的口腔菌群变化,提示其可能具有一定的分层或标志物价值。
可能机制:全身炎症、内毒素和口源菌-肠道转运是主要通路
现有证据更支持间接机制而非频繁的直接细菌移位。牙周致病菌及其产物,尤其是 LPS 和毒力因子,可激活 TLR/NF-κB 信号通路,促进 TNF-α、IL-1β、IL-6 等促炎因子释放,诱导氧化应激并激活肝内 Kupffer 细胞,从而推动脂肪变性、炎症和纤维化。
通过吞咽唾液,口源菌及其内毒素还可进入肠道,改变肠道菌群和屏障功能,经门静脉进入肝脏,形成“口腔—肠道—肝”炎症放大回路。值得注意的是, 在失代偿期肝硬化腹水患者中,研究未在腹水中检出牙龈卟啉单胞菌或 A. actinomycetemcomitans,提示临床上更可能是炎症/内毒素介导,而非直接转位至腹水。
4.炎症性肠病
双向关联、口腔—肠道轴
炎症性肠病(IBD)主要包括溃疡性结肠炎(UC)和克罗恩病(CD),是一类以慢性肠道炎症、菌群失衡和免疫异常为特征的疾病。现有研究普遍提示,IBD 与牙周炎存在明确的双向关联:IBD 患者牙周炎的患病率和严重程度更高,而牙周病人群发生 IBD,尤其是 UC 的风险也可能升高。这提示两者可能通过“口腔—肠道轴”相互影响。
微生物机制:口源致病菌与肠道失衡相互牵连
牙周炎相关菌群可通过消化道或血源途径影响肠道生态,其中较受关注的“桥接菌”包括 Klebsiella、Porphyromonas gingivalis、Fusobacterium nucleatum 、 Candida albicans。
其中,P. gingivalis 及其所在的“红复合体”成员 Treponema denticola、Tannerella forsythia 是牙周炎的重要致病菌;F. nucleatum 可破坏肠黏膜屏障;Candida albicans 也可能参与肠道炎症放大。
IBD肠道菌群常表现为多样性下降,厚壁菌门减少,变形菌门增加;放线菌门在部分研究中增加;拟杆菌门的变化各研究结果不一,不同IBD亚型间存在差异。其中产丁酸菌 Faecalibacterium prausnitzii 明显减少,被认为与肠道抗炎能力下降密切相关。
IBD 对口腔微生态的反向影响
IBD 不仅累及肠道,也会反向改变口腔生态。已有研究发现,IBD 患者口腔中 Spirochaetes、Bacteroidetes 、 Prevotella 增多,而健康人口腔中常见的 Streptococcus 减少;部分研究还提示口腔 Firmicutes、Fusobacteria 下降。由于这些变化与牙周炎相关生态失衡存在重叠,IBD 可能通过重塑口腔微环境,进一步促进牙周炎症发展。
牙周炎与炎症性肠病之间的微生物相关性
编辑
doi: 10.3389/fcimb.2023.1132420
共同免疫通路:Th17/Treg 失衡与促炎因子升高
IBD 与牙周炎共享多条免疫炎症机制,其中 Th17/Treg 失衡 是最核心的共同通路之一。口源致病菌可通过 TLR/MyD88/NF-κB 等信号通路激活免疫反应,促进 IL-1β、IL-6、TNF-α、IL-17 等促炎因子升高,并通过 RANKL、M-CSF 和 MMP-9 等途径加重组织破坏和屏障损伤。相对地,Treg 相关的 IL-10 和 TGF-β 保护作用减弱,也会进一步推动口腔与肠道炎症的恶性循环。
总体来看,IBD 与牙周炎之间存在较明确的双向联系,其核心基础在于口源致病菌向肠道迁移、IBD 反向重塑口腔微生态,以及共同的免疫炎症失衡。不过,现有证据仍以相关性研究为主,其因果关系及干预价值仍需更多纵向和机制研究进一步验证。
5.类风湿关节炎
口腔微生物组可能是重要诱因之一
类风湿关节炎(rheumatoid arthritis, 简称RA)是一种以关节滑膜慢性炎症为核心的系统性自身免疫病,通常由遗传易感(如特定HLA-DRB1基因型)与环境因素(如吸烟、感染)共同触发。
牙周炎与类风湿关节炎:关联强、且可能可逆
近年证据提示,牙周炎相关的口腔微生物组失调可能是RA的重要环境触发因素之一。流行病学研究相对一致地显示:RA患者更易合并且更易出现重度牙周炎;反之,牙周炎人群RA风险上升。
更值得注意的是,在RA患者中,牙周炎严重程度往往与疾病活动度(如DAS28)及炎症指标(CRP、ESR)呈正相关;而非手术牙周治疗(NSPT)在改善牙周状况的同时,也被报道可在一定程度上降低RA活动度。
这些现象提示:口腔炎症与RA并非简单“共病”,而可能共享可被干预的炎症—免疫通路。
关键桥梁:异常瓜氨酸化与ACPA,并非一定从关节开始
RA最具代表性的免疫学特征之一是抗瓜氨酸化蛋白抗体(ACPA)。
ACPA常在关节症状出现前多年即可检出,提示RA的自身免疫启动可能发生在关节之外。
“黏膜起源假说”据此提出:口腔、肺、肠道等黏膜部位的慢性炎症与微生物刺激,可能先诱导异常的蛋白瓜氨酸化,继而打破免疫耐受,推动ACPA产生与成熟。牙周炎局部环境中宿主PAD酶活性升高、龈沟液/牙龈组织中瓜氨酸化蛋白增加,也为“口腔先行”的路径提供了生物学可解释性。
类风湿性关节炎发病机制中的口腔-肠道-免疫-营养轴
编辑
doi: 10.3390/ijms27052385
两类机制型关键菌:牙龈卟啉单胞菌直接促瓜氨酸化;伴放线聚集杆菌诱导超瓜氨酸化
高危个体的口腔失调:可能早于关节炎发作
在尚未出现典型关节炎表现、但已ACPA阳性的RA高危人群中,研究者已观察到口腔微生物组失调特征:即使在牙周临床表现不典型的位点,也可出现偏向牙周炎样的生态结构,并更易检出或富集与牙周炎相关的菌群(其中包括上述关键菌)。
与此同时,“双向瓜氨酸化/分子模拟”也被提出:宿主炎症反应既可瓜氨酸化自身蛋白,也可能瓜氨酸化细菌蛋白;部分抗体可能先针对细菌抗原产生,随后因交叉反应转向自身靶点,从而推动自身免疫扩展。
总体而言,现有证据支持:牙周炎相关口腔微生物组失调可能通过异常瓜氨酸化—ACPA轴参与RA的启动与放大,其中牙龈卟啉单胞菌(PPAD)与伴放线聚集杆菌(LtxA)是目前机制链条中最关键的两类病原线索。
因此,从口腔微生物组视角看,RA并不一定是“从关节开始”的疾病;在遗传易感个体中,口腔黏膜可能提供了一个长期、低度但持续的自身抗原生成与免疫驱动场景,也使口腔健康干预成为值得关注的辅助策略之一。
干预方向
非手术牙周治疗(NSPT),包括龈上刮治、龈下刮治和强化口腔卫生指导,是牙周管理的标准措施,也是类风湿关节炎和牙周炎患者最常采用的干预措施。非手术牙周治疗可以显著改善牙周结果,如探诊深度、附着丧失和牙龈出血指数。更重要的是,在接受神经内分泌治疗的类风湿关节炎患者的短期随访中,他们的全身炎症指标(如疾病活动评分和急性期反应物)显示出下降趋势。
然而,这些系统性益处通常仅在3到6个月的随访期内有效,其可持续性尚未通过长期研究得到确认。
抗菌辅助治疗:在牙周临床实践中,阿莫西林与甲硝唑的短期系统应用是非手术治疗的常见强化措施。特别是对于具有侵袭性或难治性牙周炎的患者。
系统性抗生素治疗需要考虑药物耐药性和个体耐受性的风险。因此,抗菌辅助治疗应被视为RA患者严重或难治性牙周炎的个体化选择,并需仔细评估与微生物群破坏相关的潜在系统性影响。
生物制剂:研究表明,生物制剂如抗TNF和抗IL-6与某些患者牙周炎症指标的降低相关,提示全身炎症的缓解可能对牙周健康带来益处。
▸
普遍认为,良好的口腔卫生行为(如定期刷牙、使用牙线和牙间刷)、戒烟以及定期专业维护和口腔菌群检测可以显著降低牙周病复发的风险。对于类风湿患者,纳入口腔健康教育和长期随访可以帮助减少局部炎症负担,从而在一定程度上降低全身炎症水平。
多学科合作是提高治疗效果的关键:类风湿科与牙周医生的联合管理可以实现疾病信息共享和协调治疗计划,避免治疗盲点,促进RA和牙周病的双重控制。未来的临床研究还应将牙周和类风湿结果设定为共同终点,以更全面地评估干预的临床意义。
6.高血压
信号明确,但特征菌尚未定型
高血压(Hypertension)是全球超过12亿人面临的重大健康问题。越来越多的研究表明,它与口腔微生物群密切相关。
与T2D和肥胖相比,高血压与口腔微生物组的结果具有异质性。“两者有关联”这一点明确,但具体是哪些菌升高、哪些菌下降,不同研究之间并不总是一致。
高血压口腔菌群变化
从群落特征看,关于α多样性和β多样性的结果并不统一:有研究发现高血压患者多样性下降,也有研究发现升高,还有一些研究未见显著差异。
具体而言,Atopobium、Prevotella、Bacteroides、Streptococcus、Veillonella等细菌在高血压患者的唾液中丰度显著升高。
牙周相关菌群,可能是更稳定的一条线索
相比“单个属是否升高”,牙周致病相关菌群的富集也许是理解高血压更有意义的切入点。
牙周炎的致病菌和抗体负荷较高与收缩压、舒张压和高血压患病率显著相关。据报道,包括中间普里沃菌(P. intermedia)、牙龈卟啉单胞菌(P. gingivalis)和具核梭杆菌(F. nucleatum)在内的多种牙周细菌与高血压显著相关。
口腔微生物易位参与了高血压的进展
尽管在生理条件下,胃和胆汁酸可以杀死大多数微生物,但口腔微生物仍可通过吞咽直接进入肠道,尤其是在牙周感染的情况下。
来自重度牙周炎患者的卟啉单胞菌科和梭杆菌属可在小鼠肠道中易位并定植。牙龈卟啉单胞菌通过FimA菌毛侵入树突状细胞作为转运载体,从而进入人类主动脉内皮细胞,对高血压构成威胁。
在接受人类口腔微生物移植的高血压患者和高血压小鼠中,韦荣氏球菌属是口腔-肠道间常见的稳定富集的传递菌。
菌群代谢物的调节作用
在重度牙周炎患者和实验性牙周炎ApoE-/-小鼠的血清中,发现三甲基胺N-氧化物(TMAO)水平升高,导致肠道菌群失调和内皮功能受损(图A)。高血压患者的独特代谢簇与龈下菌斑中的微生物群显著相关。
编辑
doi: 10.3389/fcimb.2025.1731845
牙周病原体通过诱导内皮功能障碍来加重高血压
研究发现,牙周炎患者的肱动脉血流介导性扩张(FMD)下降,暗示内皮功能受损。同时,严重感染的牙周组织释放的促炎介质可进入体循环,进而影响内皮功能。宿主免疫系统对微生物感染作出反应,因此不同易感性的宿主个体之间血管内皮损伤程度不同。牙周炎症可通过血管内皮细胞上的受体(包括TLR-2和TLR-4)检测到,从而激活炎症级联反应,释放促炎细胞因子并促进粘附分子的表达。
一氧化氮(NO)水平的调节
口腔微生物组通过调节一氧化氮(NO)水平影响高血压。
NO是由内皮细胞产生的血管舒张剂,具有重要的生理功能。内源性NO由特定的NOS合成,以L-精氨酸为底物,利用氧气、NADPH等(还需BH4等多种辅因子),将L-精氨酸转化为L-瓜氨酸,同时生成NO。
NOS主要分为三类:
神经型NOS、诱导型NOS、内皮型NOS
其中内皮型NOS(eNOS)是维持内皮完整性和血管张力的关键调节因子。
释放的NO可以立即被氧化为亚硝酸盐和硝酸盐。唾液腺和饮食中的硝酸盐可以通过口腔细菌中的硝酸盐还原酶还原为亚硝酸盐,随后再被去硝化细菌转化为NO。
研究表明,正常血压个体的唾液中NO和微生物多样性较高。
多种口腔微生物,包括Neisseria、Haemophilus、Granulicatella、Veillonella、Prevotella、Corynebacterium、Actinomyces、Rothia,能够还原硝酸盐。
具体而言,Streptococcus mutans、Fusobacterium nucleatum、Veillonella dispar具有亚硝酸盐还原能力。然而,感染牙龈卟啉单胞菌的内皮细胞会产生过量的活性氧(ROS),这可能干扰NO通路,并降低NO的生物活性。
值得注意的是,NO的释放还具有抗菌活性,能够对抗与牙周炎相关的细菌,如Aggregatibacter actinomycetemcomitans、F. nucleatum。然而,使用抗菌漱口水可能会导致口腔微生物组的组成变化,从而影响NO的产生。
多项研究表明,使用抗菌漱口水会降低唾液和血浆中的硝酸盐/亚硝酸盐浓度,导致血压升高。相反,含有无机硝酸盐和抗氧化剂的漱口水可以影响口腔微生物组的多样性和结构,促进肠-唾液循环,并在一周内有助于降低血压。
另一项荟萃分析未显示漱口水使用与血压之间存在显著关系。这种不一致的可能原因包括漱口水的成分和使用时间的差异,以及高血压的严重程度不同。
理解这一机制不仅有助于我们更好地认识高血压的病理生理,还可能为其预防和治疗提供新的思路。
心血管疾病(CVDs)中口腔-肠道轴的机制
编辑
doi: 10.3389/fcimb.2025.1658502
7.动脉粥样硬化
口腔菌群参与血管炎症与斑块形成
动脉粥样硬化的核心病理基础是血管内膜脂质沉积、慢性炎症和内皮功能障碍。
口腔菌群在血管炎症与斑块形成中的作用
现有研究提示,口腔微生物组可能通过系统性炎症、菌血症、血管内皮损伤以及代谢紊乱等多条路径参与其进展。牙周炎患者常伴随白细胞计数升高以及 TNF-α、IL-6、IL-8、aMMP-8 等炎症标志物升高,这种低度全身炎症可促进内皮功能障碍,并增加动脉粥样硬化风险。
口腔菌群的变化与动脉粥样硬化的关联
在菌群层面,已有研究发现,动脉粥样硬化患者唾液中 Streptococcus、Lactobacillus、Actinobacteria、Cutibacterium 的丰度较高,而 Bacteroidetes 相对减少。
多项研究在动脉粥样硬化斑块中检测到了口腔来源细菌或其DNA,其中较常被提及的包括 Veillonella、Streptococcus 以及牙周相关病原体牙龈卟啉单胞菌。
有研究甚至提示,牙龈卟啉单胞菌是冠状动脉和股动脉标本中检出的优势菌之一。与此同时,Fusobacterium nucleatum 还被报道可通过促进肝脏糖酵解和脂质生成,加重动脉粥样硬化表型。
8.心肌梗死
可能影响梗死后损伤修复
心肌梗死(MI)患者常表现出更差的牙周状态,且口腔微生物组组成也出现改变。
心肌梗死:牙周健康与口腔微生物组变化
临床研究显示,心肌梗死患者往往具有更深的牙周袋和更重的牙周炎,并伴随牙龈卟啉单胞菌和普雷沃菌属丰度升高;此外,针对 Tannerella forsythia 的较低抗体水平还被报道与心肌梗死患者更高的心血管死亡风险相关。
在更直接的证据层面,研究者已在心肌梗死患者斑块中检测到Viridans Streptococci和 Aggregatibacter actinomycetemcomitans。
口腔微生物组对心肌梗死及其后果的影响
动物实验进一步提示,口腔菌群不仅可能与梗死发生有关,还可能影响梗死后的炎症修复过程:例如,牙龈卟啉单胞菌可侵入缺血心肌,其LPS可激活 CD8+ T细胞、增强 MMP-9 活性并加重心肌破裂风险;其毒力因子 gingipains 还可通过干扰自噬过程损伤梗死后心肌。
另有研究提示,Streptococcus 可沿“口腔—肠道轴”转移并加重实验性心肌梗死。因而,从口腔微生物组角度看,心肌梗死相关改变不仅体现在有无某种菌,更涉及炎症放大、免疫重编程和远隔器官损伤修复失衡。
9.感染性心内膜炎
口腔菌血症具临床指向性的例证
感染性心内膜炎(IE)是一种涉及心脏瓣膜感染的疾病。在几类心血管疾病中,感染性心内膜炎与口腔微生物的联系最为直接。
链球菌属(尤其口腔链球菌)是口腔中最常见、同时也是心内膜炎中重要的一类病原。口腔来源链球菌不仅可进入血流,还可通过影响单核细胞向树突状细胞分化,干扰宿主对病原体的清除。
除链球菌外,口腔微生物中的 HACEK 群也与感染性心内膜炎密切相关,包括 Haemophilus parainfluenzae、Aggregatibacter spp.、Cardiobacterium spp.、Eikenella corrodens \Kingella spp.。
其中,A. actinomycetemcomitans 的关联尤受关注。该菌不仅与牙周感染相关,还可借助黏附因子 EmaA 结合心脏瓣膜中高表达的I型胶原,因此被认为是从口腔到心脏瓣膜传播的代表性病原之一。
总体而言,一个清晰的“口腔—心脏”路径:牙周炎或口腔操作导致菌血症,随后病原在易感瓣膜表面黏附、定植并形成感染灶。
心血管疾病相关口腔-肠道菌群的作用机制
编辑
doi: 10.3389/fcimb.2025.1658502
10.心力衰竭
证据仍有限,但炎症与NO通路值得关注
已有研究提示,较差的牙周状态和口腔微生物多样性变化与心力衰竭发生风险有关;同时,P. gingivalis抗体水平升高也与更高的心力衰竭患病风险相关。
动物研究进一步表明,P. gingivalis 诱导的牙周炎可加速保留射血分数心衰(HFpEF)的进展,其机制可能涉及全身炎症升高和NO耗竭。此外,在射血分数降低型心衰(HFrEF)患者中,还观察到口腔硝酸盐还原能力下降,提示口腔菌群参与的硝酸盐–亚硝酸盐–NO通路可能是心力衰竭的重要调节环节。
由于NO与血管舒张、微循环灌注和心肌功能密切相关,这一发现使调节口腔微生态或补充无机硝酸盐成为潜在干预方向。另一方面,心力衰竭本身也可通过肠道屏障受损加重肠道失调,而口腔菌群向肠道转移可能进一步放大全身炎症,形成“口腔—肠道—心脏”恶性循环。
11.阿尔茨海默
口腔菌群可能参与,不只是伴随现象
如果说心血管疾病中的关键问题,是口腔炎症如何影响血管;那么在阿尔茨海默病(AD)中,更值得关注的则是:口腔微生物组失调,是否会通过慢性炎症、屏障破坏和神经免疫激活,参与神经退行性过程。
现有证据提示,牙周炎、牙齿缺失和较差口腔卫生与认知下降及 AD 风险升高有关。包括 ARIC 在内的一些前瞻性人群研究,以及多项医保数据库研究,都反复给出了相似方向的信号:口腔健康越差,后续认知结局往往也越不理想。
需要强调的是,这类研究首先说明的是“相关”,而不是已经完成严格意义上的因果证明。
口腔菌群失调与阿尔茨海默病
编辑
doi.org/10.1016/j.bcp.2025.117362
牙周相关病原体及其成分在中枢层面的线索
在 AD 相关研究中,最常被提及的菌是牙龈卟啉单胞菌(P. gingivalis)。
Dominy 团队等研究曾在 AD 患者脑组织中检测到 P. gingivalis 及其毒力因子 gingipains;此外,Treponema denticola、其他口腔螺旋体,以及部分口腔来源细菌 DNA,也在部分研究中被报道与 AD 脑组织或外周标志物有关。
从机制上看,关键可能是“牙周炎储源”持续向脑部相关通路施压
牙周炎可反复造成短暂菌血症,并持续释放LPS、蛋白酶和促炎介质。在老龄化、血脑屏障功能下降和全身低度炎症存在的背景下,这些信号更可能通过血行途径放大神经炎症。尤其是 P. gingivalis 的 LPS 和 gingipains,被认为可激活小胶质细胞,诱导 TNF-α、IL-1β、IL-6 等炎症反应,并与 Aβ 沉积、tau 异常磷酸化 等 AD 关键病理过程相关。动物实验也提示,口腔感染或牙周致病菌暴露可加重脑内炎症和淀粉样病理改变。
“口腔-大脑轴”以及牙龈卟啉单胞菌在阿尔茨海默病病理学中的拟议作用
编辑
doi: 10.1080/20002297.2026.2635233
AD 相关的口腔微生物信号
以 P. gingivalis、T. denticola、Tannerella forsythia 等为代表的牙周致病生态位,在长期失调状态下,持续提供炎症刺激、病原成分和免疫扰动;而宿主衰老、屏障脆弱和神经免疫失衡,则决定了这些信号是否会进一步转化为中枢损伤。
牙周炎可能作为一个长期炎症与病原储源,通过系统炎症、病原转位和神经免疫激活,参与 AD 的发生发展。
不过,现阶段证据仍以关联和机制研究为主;要把它真正提升为可操作的风险预测或干预靶点,仍需要更多纵向队列和干预研究支持。
12.帕金森
口腔致病菌增多,牙周病风险升高
多项研究发现,帕金森(PD)患者在唾液、龈下菌斑和口腔软组织的微生物组谱与健康对照组不同。
基于16S rRNA测序的研究显示,PD患者口腔软组织菌群的β多样性显著降低,并表现出更高的致病菌负荷。常见的富集类群包括Prevotella、Streptococcus、Lactobacillus,此外,Streptococcus mutans、Kingella oralis、Veillonella等菌群也有所增加,而Treponema、Lachnospiraceae相对减少。
帕金森的流涎、吞咽困难及唾液pH变化可能重塑口腔微环境,因此这些差异应视为与帕金森相关的微生态信号。
牙周炎与PD风险
牙周疾病被视为帕金森的危险因素,病例-对照研究显示帕金森与探诊出血及炎症标志物升高相关。中国台湾省的回顾性匹配队列显示,牙周病患者随访中的PD新发风险更高(调整后HR≈1.43)。此外,在无牙周病个体中,定期洁治与PD风险降低相关。
13.精神疾病
抑郁与焦虑、自闭症、精神分裂
抑郁与焦虑:口腔菌群改变更多体现为炎症背景下的相关信号
已有研究提示,抑郁或焦虑人群可出现唾液或其他口腔生态位菌群组成改变,部分文献反复提到 Streptococcus、Prevotella、Veillonella、Neisseria、Actinomyces 等常见口腔菌属的丰度差异。与此同时,这类人群常伴随较差的口腔卫生、牙龈炎/牙周炎负担增加以及炎症指标升高,因此口腔菌群变化很可能与慢性低度炎症状态共同出现,而不一定直接参与精神症状的发生。
口腔微生物组在抑郁、焦虑中的研究仍以横断面观察为主,现阶段更适合将其视为一种相关生物学信号,而非已确立的病因机制。
自闭症谱系障碍:可见菌群差异,但受饮食和行为因素影响较大
关于自闭症谱系障碍(ASD)的口腔微生物研究,多集中于儿童人群。现有研究普遍提示,ASD 患儿与神经典型儿童相比,在唾液、牙菌斑或口腔拭子样本中可见菌群组成差异,但不同研究之间的一致性相对有限。
被报道发生变化的通常仍是口腔常见优势菌属,如Streptococcus、Prevotella、Veillonella、Neisseria、Actinomyces 等。
这一领域解释难度较大,主要因为 ASD 儿童往往同时存在饮食偏好明显、口腔清洁配合度差异、龋病或牙周炎风险升高、抗生素或其他药物暴露等情况,上述因素都可能显著影响口腔微生态。
ASD 相关的口腔菌群改变可能反映了行为、饮食、口腔卫生与炎症状态共同作用下的生态偏移,但尚不足以作为独立的诊断标志物。
口腔微生物群影响神经精神疾病的可能机制
编辑
doi.org/10.1016/j.bbr.2024.115111
精神分裂症:菌群差异存在,但药物和生活方式混杂尤为突出
这一领域受到的混杂因素可能比前两类疾病更复杂。
吸烟率升高、抗精神病药物使用、代谢异常、口腔卫生不良及牙周疾病负担增加,均可能显著改变口腔微生态。现有研究提示,精神分裂症患者与健康对照之间可见菌群组成差异,涉及的菌群仍多为 Streptococcus、Prevotella、Veillonella、Neisseria、Actinomyces 等口腔常见类群,以及与牙周炎生态位相关的变化。
精神分裂症中的口腔菌群异常与疾病相关的免疫炎症异常、代谢改变及口腔健康状况相互交织。未来研究尤其需要对吸烟、用药和牙周状态进行严格分层,才能更清楚地判断口腔微生物组改变的临床意义。
14.慢性肾病
口腔微生物组也会发生明显重塑
与健康对照相比,CKD患者口腔菌群的整体结构不同,且微生物多样性往往升高。在菌群组成上,链球菌属、放线菌属(Actinomyces)、纤细菌属(Leptotrichia)以及柠檬酸杆菌属(Citrobacter)等更常见;而普雷沃菌属、Alloprevotella、Haemophilus则相对减少。
CKD患者常伴随尿素代谢异常、唾液成分变化和慢性炎症状态,这些因素都可能改变口腔微环境,从而影响菌群定植与竞争。
功能预测分析还发现,CKD相关口腔菌群在精氨酸代谢和色氨酸代谢方面增强,提示其可能通过炎症放大、NO相关通路受扰以及尿毒症毒素前体增加等机制,参与疾病进展。
一些口腔菌与肾功能指标之间还存在相关性。例如,Prevotella与较好的肾功能指标相关,而部分富集菌则与血肌酐、尿素氮升高及eGFR下降相关。
从口腔微生物组的角度看,CKD不仅伴随局部微生态失调,这些变化还有望成为一种无创辅助筛查和风险评估线索。不过,目前相关证据仍以观察性研究为主,其因果关系和具体机制仍需进一步研究确认。
15.癌症
口腔癌、食管癌、胃癌、结直肠癌、肺癌等
口腔微生物组失调与多器官肿瘤相关,反复出现的关键菌主要是牙龈卟啉单胞菌、具核梭杆菌和齿垢密螺旋体。
共同机制主要包括:慢性炎症(LPS、gingipains、胶原酶、细胞裂解毒素)、直接致基因毒性(乙醛、亚硝化反应、CDT)、免疫逃逸/免疫抑制(TLR/NLR、MDSC、TAN/TAM、TIGIT/CEACAM1)以及信号与表观遗传重编程(NF-κB、MAPK/ERK、PI3K/AKT、JAK/STAT、WNT/β-catenin、Notch、COX-2、CpG高甲基化)。
可能的传播途径包括吞咽进入消化道、菌血症/淋巴播散、微误吸和黏膜直接转移。
✨ 口腔癌
口腔鳞状细胞癌(OSCC)中常见富集菌包括:P. gingivalis、F. nucleatum、
Prevotella intermedia、P. melaninogenica、Tannerella forsythia、Peptostreptococcus、Parvimonas、Capnocytophaga、
Streptococcus anginosus
其中,P. gingivalis可上调miR-21/PDCD4/AP-1、激活PI3K/AKT和NF-κB/MAPK、抑制凋亡并增强侵袭和耐药;F. nucleatum与炎症型菌群特征相关;S. anginosus可趋化并激活中性粒细胞和单核细胞,促进上皮异常增生。
部分研究还发现P. gingivalis可上调肿瘤细胞免疫检查点相关分子。
✨ 食管癌
食管癌,尤其是食管鳞癌(ESCC),最常见相关菌为P. gingivalis;其在食管组织中的检出与分期、病理特征、生存有关,血清IgG/IgA抗-P. gingivalis抗体也被提出可作为诊断/预后线索。
其他相关菌包括F. nucleatum、T. denticola、Prevotella、S. anginosus。
其中,T. denticola偏向通过持续黏膜定植、低度慢性炎症、NF-κB相关细胞因子信号和蛋白水解性组织破坏参与癌变;F. nucleatum可经NLRP3炎症小体—MDSC通路促进顺铂耐药,并可通过自噬相关机制增强化疗耐受;唾液中Prevotella及P. gingivalis/Prevotella比值也被用于风险识别。
✨ 胃癌
胃癌中常见变化包括Streptococcus富集。研究提示其与谷胱甘肽、半胱氨酸、蛋氨酸等代谢通路增强相关。
多种口腔链球菌,如S. mitis、S. gordonii、S. salivarius、S. sanguinis、S. oralis,具有酒精脱氢酶(ADH)活性,可将乙醇代谢为乙醛,增加致癌暴露。整体上,口腔来源菌可能沿口腔—胃轴参与胃内菌群重塑和代谢异常。
✨ 结直肠癌
结直肠癌(CRC)中:
最核心的菌是具核梭杆菌(F. nucleatum),
其次是牙龈卟啉单胞菌(P. gingivalis)。
具核梭杆菌
具核梭杆菌可经菌血症或淋巴播散到达结直肠,也可耐受胃酸后经消化道转移;其表面蛋白FadA与E-cadherin结合,激活β-catenin信号,Fap2可结合肿瘤表面的Gal-GalNAc并与免疫抑制受体TIGIT作用,抑制NK/T细胞杀伤;同时可诱导IL-8、CXCL1分泌,促进迁移,并在MSI-H CRC中与CD68+巨噬细胞增多相关,还可促进CD103+树突细胞和Foxp3+ Treg扩增。
具核梭杆菌促进结直肠癌进展的机制
编辑
DOI: 10.1080/20002297.2026.2635233
牙龈卟啉单胞菌
牙龈卟啉单胞菌可通过gingipains降解细胞外基质、免疫球蛋白、补体和细胞因子,促进免疫逃逸;还可激活NLRP3炎症小体、诱导细胞衰老、上调PI3K/AKT、JAK/STAT3、MAPK/ERK、WNT/β-catenin,抑制caspase-3/9介导的凋亡,并通过调控miR-203/SOCS3维持抗凋亡状态。
部分研究已尝试将粪便F. nucleatum DNA作为结直肠癌标志物。
与结直肠癌相关的口腔微生物丰度
编辑
doi: 10.3390/pathogens14121233
✨ 肺癌
肺癌相关口腔来源菌常见于Streptococcus、Prevotella、Veillonella、Haemophilus、Rothia、Neisseria、Porphyromonas等。
机制上以炎症驱动最突出。P. gingivalis和A. actinomycetemcomitans可通过TLR识别及其下游TNF-α、IL-1β、IL-6、IL-8等炎症因子上调促进肿瘤微环境形成;A. actinomycetemcomitans产生的CDT还可诱导DNA双链断裂和持续炎症。
非小细胞肺癌中TLR4、TLR5、TLR7、TLR8表达升高,部分可能与口腔病原体诱导的DNA甲基化/受体表达重塑有关。F. nucleatum还可通过FadA–E-cadherin–β-catenin轴促进侵袭和转移。
口腔微生物组与肺癌相关性的总结
编辑
doi: 10.3390/cancers18040591
干预方向
口腔保健的重要性:专业的口腔卫生干预对肺癌患者尤其重要。研究表明,良好的口腔卫生可以显著降低误吸性肺炎的发生率,这对于免疫系统受损的肺癌患者尤为关键。定期的口腔护理和牙周治疗可以帮助减少病原体的转移,从而降低肺部感染的风险。
抗微生物肽(AMPs):AMPs在肺癌治疗中也显示出潜力。这些小肽具有广谱的抗菌、抗病毒和抗肿瘤能力,可能通过调节免疫反应和直接杀死癌细胞来改善治疗效果。
将口腔健康纳入整体健康促进策略被认为是一种有效的癌症预防方法。良好的口腔卫生习惯和定期口腔菌群检测被视为降低肺癌及其他癌症风险的重要措施。
在化疗前,特别是在高剂量化疗之前,将牙周治疗纳入治疗方案具有优势。
目前已知的针对口腔微生物群的策略
编辑
doi: 10.3390/cancers18040591
✨ 胰腺癌
唾液中,胰腺癌患者可见Neisseria elongata和Streptococcus mitis减少,Veillonella、Campylobacter、Prevotella较常见;
后续研究又提示Leptotrichia、P. gingivalis、A. actinomycetemcomitans、Streptococcus 升高,舌苔中Leptotrichia、Fusobacterium也可增多。
前瞻性研究发现,抗P. gingivalis抗体水平升高与后续胰腺癌风险增加相关;胰腺癌组织中Fusobacterium spp.相对丰度更高。
机制上,P. gingivalis可激活PAR2/NF-κB、ERK1/2-Ets1和p38/HSP27,诱导proMMP-9表达并促进基质降解;还可促进肿瘤相关中性粒细胞释放中性粒细胞弹性蛋白酶,增强肿瘤发生,并可上调胰腺癌相关基因Reg3A/G。另有研究提出口腔菌可能参与p53相关突变过程。
✨ 其他
肝癌、膀胱癌、宫颈癌、乳腺癌等这些癌症反复被关注的仍是P. gingivalis、F. nucleatum、T. denticola等牙周相关菌,其可能通过慢性炎症、免疫失衡、菌血症/门静脉途径转移参与肿瘤微环境形成。
现阶段以相关性证据为主,缺乏足够机制验证。
编辑
DOI: 10.1080/20002297.2026.2635233
口腔健康是会随着机体内环境的变化而动态波动。其中,一个经常被忽视、却极为关键的因素,就是激素水平的改变。
青春期、妊娠期、更年期以及长期应激状态,都是机体激素谱发生明显变化的阶段。此时,口腔黏膜、牙龈血管、唾液分泌、局部免疫反应,乃至口腔微生物组本身,都会随之发生重塑。
换句话说,许多人在这些阶段更容易出现牙龈出血、口干、口臭、牙周炎加重或念珠菌感染,并不只是“清洁没做好”,而往往是激素变化、宿主反应和微生物生态改变共同作用的结果。
青春期
激素变化为什么会让牙龈更敏感
很多青少年出现牙龈红肿、刷牙出血,并不一定是因为菌斑突然增多,而是因为激素变化先把牙龈的“反应阈值”调低了。
性激素水平上升后,牙龈血管通透性增加,龈沟液渗出变多。GCF 中富含蛋白质、肽类、血红素及宿主组织降解产物,这会使龈下环境更有利于厌氧菌和蛋白水解型菌群生长。结果是,原本相对稳定的龈下微生态更容易向炎症相关方向偏移。
在这一过程中,Prevotella intermedia 是最经典的代表,因为它能够利用孕酮和雌二醇作为维生素 K 的替代物,从而在青春期更容易获得生长优势。与此同时,Capnocytophaga等与青春期牙龈炎相关的菌群也常可增加。如果局部菌斑控制本就欠佳,龈下生态位还可能进一步向更典型的牙周炎相关群落偏移,使 Tannerella 和 Treponema 这类厌氧菌更容易稳定存在。
编辑
此外,青春期也常伴随饮食频率增加、含糖零食摄入上升和口腔清洁不稳定,这会使龋病风险同步增高。在这一背景下,S. mutans 等致龋菌也更容易在菌斑中获得优势。因此,青春期口腔问题增加,往往并不局限于牙龈炎,而是可能同时表现为牙龈炎症易感性上升与龋病风险增加。
妊娠期
为什么孕期更容易出现牙龈炎和菌群波动?
妊娠期是激素波动最显著的生理阶段之一。高水平雌激素和孕激素会明显增加牙龈血流和血管通透性,使龈沟液渗出增多,牙龈组织更容易肿胀、充血和出血。与此同时,局部免疫反应也发生适应性调整,使牙龈对菌斑刺激的炎症反应更强,这就构成了“妊娠期牙龈炎”的基本背景。
从微生物角度看,孕期最典型的仍然是 P. intermedia,因为它可利用孕酮和雌二醇促进生长。
编辑
除此之外,研究还常观察到牙周相关菌增加,包括 Porphyromonas gingivalis、Tannerella forsythia 、 Treponema denticola 等,使龈下微生态更偏向经典牙周致病群落。
越来越多证据提示,口腔菌群本身也在参与局部激素代谢。与牙周病相关的 Tannerella、Treponema、Prevotella 等菌属中存在表达 GUS 酶的成员,理论上可参与结合型雌激素的去结合与再活化;而 S. mutans 和 T. denticola 等细菌则被发现具有 5α-还原酶和羟类固醇脱氢酶等相关活性,可能影响孕激素和雄激素在口腔局部的利用方式。
这也解释了为什么孕期菌群波动常常具有一定可逆性。分娩后随着激素水平回落,牙龈血管通透性、GCF 渗出和局部免疫状态逐渐恢复,孕期被“偏袒”的厌氧菌群也可能部分回落。
因此,妊娠期口腔变化:高激素状态下口腔微生态被暂时推向更促炎、更利于牙周相关菌扩张的方向。
激素-宿主与激素-微生物的相互作用
编辑
Rus et al., 2025
更年期
口干、炎症和机会性感染为何增加
更年期与长期应激:口腔黏膜屏障变弱,唾液防御下降,炎症更容易持续,机会致病微生物也更容易获得优势。
更年期——雌激素下降
雌激素减少后,口腔黏膜可出现萎缩和干燥,唾液分泌也常减少。
唾液不仅是润滑液,更是口腔最重要的“生态稳定器”之一:它负责机械冲刷、缓冲酸性环境,并提供抗菌蛋白和免疫分子。当唾液减少时,口腔清除能力下降,酸性代谢产物更易积聚,黏膜也更容易受到刺激和损伤。
在这种环境下,Candida albicans 等机会致病真菌更容易由共生状态转向致病状态,而 S. mutans 等致龋菌也可能因低唾液、低缓冲状态而更容易获得优势,因此更年期常同时表现为口干、灼口感、龋病风险增加以及机会性感染增多。
与此同时,雌激素下降还会影响牙周组织和牙槽骨稳态,使已有牙周基础问题的人更容易出现炎症持续和附着丧失。在这种背景下,龈下菌群也更容易向牙周炎相关群落偏移,包括 P. gingivalis、Tannerella forsythia、Treponema denticola 等菌更可能持续存在并维持慢性炎症。
应激状态——皮质醇升高
皮质醇可通过调节甚至抑制局部免疫反应,削弱机体对病原体的控制能力,使炎症更容易迁延不愈。研究发现,长期应激与牙周病加重相关,且唾液皮质醇水平升高时,一些牙周相关菌和促炎菌也更容易增加。在这种背景下,Fusobacterium nucleatum、P. gingivalis、Tannerella、Treponema 等菌更可能在牙周袋环境中扩张,并形成“菌群失调—炎症持续—宿主防御下降”的循环。
因此,无论是更年期还是长期应激,口腔问题在于激素改变了唾液、黏膜、免疫和龈下生态位,最终让致龋菌、牙周相关菌和机会致病真菌更容易获得生存优势。
总的来说,激素改变宿主,宿主改变生态位;而生态位一旦改变,微生物组就会随之重排。口腔在这些生理阶段表现出的“敏感”“易炎症”或“易感染”,本质上正是这种激素—微生物互作的临床体现。
鉴于口腔微生物组在健康与疾病中的核心作用,通过主动干预来调节其组成和功能,以恢复或维持微生态平衡,已成为现代口腔医学和预防医学的重要方向。当口腔微生物组从共生状态转向失调状态时,多种干预措施可以用来打破疾病的恶性循环,促进向健康状态的恢复。
传统干预
生物膜控制与化学疗法
在早期阶段,重点是预防和建立独立的护理习惯。传统的口腔健康维护策略主要集中在对牙菌斑生物膜的物理和化学控制上。
机械性菌斑去除
这是最基础也是最重要的干预手段。日常的刷牙和使用牙线(或牙间刷)能够物理性地破坏和清除牙齿表面的生物膜,阻止其成熟和矿化(形成牙结石)。
对于已经形成的龈下菌斑和牙结石,则需要通过专业的牙周治疗,如龈上洁治(洗牙)和龈下刮治及根面平整(SRP),由牙科专业人员使用专门器械进行彻底清除。这些机械性方法是治疗牙龈炎和牙周炎的基石,其目标是尽可能地降低微生物负荷,为牙周组织的愈合创造条件。
抗菌药物与化学制剂
在某些情况下,单纯的机械清创不足以控制感染,需要辅助使用化学制剂。
——漱口水
含有氯己定(Chlorhexidine)或西吡氯铵(Cetylpyridinium Chloride)等广谱抗菌成分的漱口水,可以作为短期辅助措施,用于控制菌斑和牙龈炎。然而,长期使用(特别是Chlorhexidine)可能导致牙齿着色、味觉改变以及对口腔微生物组的非选择性破坏,甚至可能干扰有益的硝酸盐还原通路,因此不推荐常规长期使用。
——抗生素
在重度或侵袭性牙周炎的治疗中,有时会在SRP的基础上联合使用全身性抗生素(如甲硝唑联合阿莫西林)。抗生素能够有效杀灭侵入组织的深层病原体。然而,抗生素的滥用是导致抗菌素耐药性(Antimicrobial Resistance, AMR)全球危机的主要原因。口腔微生物组被认为是耐药基因的一个重要“储存库”(即“口腔抗性基因组”),这些耐药基因可以通过水平基因转移在不同细菌之间传播。因此,抗生素的使用必须严格遵循指南,仅限于必要的病例,以避免加剧耐药性问题。
全身抗生素仅在C级牙周炎病例中适用。
早期筛查
风险评估
通过分析口腔微生物组的组成,可以识别潜在的健康风险。例如,特定的致病菌如牙龈卟啉单胞菌与心血管疾病、糖尿病等慢性病的发生密切相关。
定期的口腔微生物检测可以帮助医生及早发现这些风险,从而进行干预。根据患者的口腔微生物组特征,制定个性化的口腔护理和治疗方案。
为了有效应对这些潜在风险,传统的生物膜控制与机械性菌斑去除是早期干预的基础,日常的刷牙和使用牙线能够有效阻止牙菌斑的形成,降低微生物负荷,为牙周组织的愈合创造条件。这些措施不仅有助于维护口腔健康,还为个性化护理方案的实施提供了支持。
新兴干预
生态学调节策略
新兴的干预方法更侧重于从生态学角度调节微生物组,旨在“扶持益菌、抑制害菌”,恢复微生态的健康平衡。
饮食干预
◎地中海饮食:其抗炎特性可能对改善牙周健康和整体健康有益,尤其是在与类风湿关节炎和其他炎症性疾病相关的背景下。
◎抗氧化剂丰富的食物:富含抗氧化剂的水果和蔬菜(如柠檬和浆果)被推荐用于维持口腔微生物组的健康。
◎多酚和类黄酮:最近的证据表明,食物来源的天然产品,尤其是多酚和类黄酮,作为P. gingivalis毒力因子的强效抑制剂,专门针对牙龈蛋白酶(Rgp和KGP)以减弱细菌的致病性。这些化合物不仅抑制细菌生长和生物膜形成,还通过干扰膜结合的特异性转运酶破坏能量代谢。
◎Omega-3脂肪酸补充剂:已被证明在作为牙周治疗的辅助治疗时改善临床参数(探诊袋深度和临床附着水平)。这种双重作用(减少口腔病原负荷和毒力,同时调节肠道微生物群)强化了营养在干预口腔-肠道轴源头中的作用。
◎姜黄素:作为一种多酚化合物,具有抗炎、抗氧化、抗菌特性。它通过调节关键的炎症通路(如NF-κB)和氧化应激反应,展现出对多种口腔疾病的治疗潜力,如牙周病、龋齿、复发性口腔炎等。
多种口腔疾病中姜黄素制剂的临床试验总结
编辑
DOI:10.3389/FPHAR.2025.1661443
临床上,局部姜黄素已被证明能够减少慢性牙周炎患者的微生物负荷。姜黄素甚至可以在非常低的浓度(20 μg/mL)下抑制P. gingivalis生物膜形成超过80%。
姜黄素的抗菌作用包括对革兰氏阳性菌、革兰氏阴性菌和真菌的抑制,是一种有吸引力的辅助或替代治疗选择。
姜黄素与药物协同治疗
编辑
DOI:10.3389/FPHAR.2025.1661443
益生菌
一些益生菌能产生细菌素、过氧化氢和有机酸,直接抑制致病菌的生长。例如,唾液链球菌K12和M18菌株(Streptococcus salivarius K12, M18)能产生细菌素,抑制引起咽炎的化脓性链球菌和部分致龋菌。
注:唾液链球菌K12菌株(S. salivarius BLIS K12)能产生细菌素,主要抑制引起咽炎的化脓性链球菌(S. pyogenes);M18菌株(BLIS M18)则更侧重于抑制致龋菌 S. mutans,两者功能各有侧重。
调节免疫反应:益生菌可以与宿主免疫细胞相互作用,调节细胞因子的产生,减轻过度的炎症反应。
目前,已有研究表明,含有特定乳杆菌(如Lactobacillus reuteri)或双歧杆菌(如Bifidobacterium animalis subsp. lactis HN019)的益生菌产品,作为牙周治疗的辅助手段,可能有助于改善牙周临床指标。
一项研究表明,作为冷冻干燥片的L. salivarius可使牙周炎患者的下列菌群减少:
P. intermedia、P. gingivalis、T. forsythia、
T. denticola、A. actinomycetemcomitans
另一个潜在的菌株是益生菌Clostridium butyricum MIYAIRI 588(CBM588),其在肠道失调的治疗中显示出疗效。C. butyricum被报道能够降低全身炎症反应,促进稳态维持,并减少肺癌患者化疗引起的腹泻。2024年报告的研究结果强调CBM588作为牙周治疗的治疗辅助剂的有效性。
另一项研究的作者证明,接受ICI治疗的非小细胞肺癌(NSCLC)患者接受MIYAIRI588后,生存期倾向于延长,与未治疗组相比。
然而,关于口腔益生菌的最佳菌株、剂量和应用方式,仍需要更多高质量的临床研究来确定。
益生元
在口腔中,一些不易被宿主或致病菌利用的碳水化合物,如木糖醇和某些寡糖,可以被视为益生元。木糖醇不能被变形链球菌有效代谢,但会被其摄取,从而干扰其能量代谢并抑制其生长。同时,木糖醇还能刺激唾液分泌,有助于中和酸性环境。如前所述,膳食中的硝酸盐也可以被看作一种益生元,它能选择性地促进硝酸盐还原菌的生长,这些菌通过产生亚硝酸盐来抑制致病菌并有益于心血管健康。
噬菌体
噬菌体是感染和杀死细菌的病毒,具有极高的特异性。这使得能够有针对性地消灭单一病原物种,如牙龈卟啉单胞菌,而不会对周围生态系统造成附带损害。
在牙周炎的背景下,噬菌体疗法的目标是诱导失调生物膜群落的解散,从而解决炎症,允许健康的、平衡的群落重新建立。
另一种精准方法涉及针对特定的毒力因子。例如,已经开发出针对P. gingivalis的牙龈蛋白酶的小分子抑制剂。通过中和这一关键毒力因子,这些抑制剂可以使病原体失去能力,减少其造成组织破坏和操控宿主免疫反应的能力。这些化合物目前正在临床试验中,潜在用于治疗牙周炎以及AD,因为牙龈蛋白酶在神经炎症中的作用。
这些高度针对性的策略体现了恢复生态平衡而非无差别杀灭微生物的转变,提供了一种更可持续且潜在更有效的方法来管理与微生物失调相关的疾病。
靶向治疗策略的转化通路
编辑
doi: 10.3389/fcimb.2025.1658502
促进口腔健康和预防癌症的公共卫生干预战略框架
编辑
doi: 10.1007/s12672-026-04527-x
口腔,是我们与外部世界最早接触的界面,也是最容易发生生态失调、却最容易被忽视的部位。
纵观全文,我们看到的不只是一份关于”口腔细菌”的科普,而是一幅口腔微生物组与全身健康深度交织的系统性图景。
龋齿、牙周炎,是菌群失调在口腔局部的最初体现;糖尿病、心血管疾病、类风湿关节炎、阿尔茨海默病,乃至结直肠癌、胰腺癌,是这种失调通过炎症通路、菌血症和神经免疫信号向全身蔓延的可能后果。这条路径,已经从流行病学的相关信号,逐渐向机制层面的清晰证据推进。
和肠道菌群检测相比,口腔菌群检测具有几个独特优势:
换言之,口腔菌群检测不只是牙科问题,它正在成为一种面向全身健康的早期预警工具。
未来,随着组学技术的成本下降、唾液,肠道等多组学检测的普及,以及人工智能辅助菌群解读能力的提升,口腔,肠道联合微生物组有望成为慢性病早筛、个性化健康管理和精准干预不可或缺的一环。
注:本账号内容仅供学习和交流,不构成任何形式的医疗建议。
主要参考文献
Dioguardi M, Lo Muzio E, Guerra C, Sovereto D, Laneve E, Martella A, Aiuto R, Garcovich D, Caloro GA, Cantore S, Lo Muzio L, Ballini A. Liver Disease and Periodontal Pathogens: A Bidirectional Relationship Between Liver and Oral Microbiota. Dent J (Basel). 2025 Oct 31;13(11):503.
Lyu Z, Zhu J, Chen D. Chronic Periodontitis and Non-Alcoholic Fatty Liver Disease: Recent Advances in Mechanisms of Association. Int J Gen Med. 2025 Dec 7;18:7357-7369.
Zhou T, Xu W, Wang Q, Jiang C, Li H, Chao Y, Sun Y, A L. The effect of the “Oral-Gut” axis on periodontitis in inflammatory bowel disease: A review of microbe and immune mechanism associations. Front Cell Infect Microbiol. 2023 Feb 27;13:1132420.
Sulaiman Y, Pacauskienė IM, Šadzevičienė R, Anuzyte R. Oral and Gut Microbiota Dysbiosis Due to Periodontitis: Systemic Implications and Links to Gastrointestinal Cancer: A Narrative Review. Medicina (Kaunas). 2024 Aug 29;60(9):1416.
Nikolić N, Pucar A, Tomić U, Petrović S, Mihailović Đ, Jovanović A, Radunović M. Oral Microbiota and Carcinogenesis: Exploring the Systemic Impact of Oral Pathogens. Pathogens. 2025 Dec 3;14(12):1233.
Hu C, Wang S, Gao Z, Qing M, Tan L, Yang L, Li F. Curcumin in oral health: mechanisms, clinical evidence, and delivery strategies. Front Pharmacol. 2026 Mar 2;16:1661443.
Pertiwi TH, Suraya R, Akashi M, Nagano T. The Role of the Oral Microbiome in Modulating Therapeutic Outcomes in Lung Cancer: Key Commensals and Clinical Implications. Cancers (Basel). 2026 Feb 11;18(4):591.
Sun G, Cheng H, Yin M. Oral hygiene and cancer risk: emerging evidence and public health perspectives. Discov Oncol. 2026 Jan 29;17(1):359.
Guo S, Wu G, Liu W, Fan Y, Song W, Wu J, Gao D, Gu X, Jing S, Shen Q, Ren L, Dong Y, Liu Z. Characteristics of human oral microbiome and its non-invasive diagnostic value in chronic kidney disease. Biosci Rep. 2022 May 27;42(5):BSR20210694.
Guo X, Wang S, Ding J, Liu W, Xu J, Wang M, Sun H, Ma Y, Liu W, Zhang L, Liu M. Periodontopathic bacteria in rheumatoid arthritis pathogenesis: bridging clinical associations to molecular mechanisms. Front Immunol. 2025 Oct 17;16:1681037.
Reytor-González C, Román-Galeano NM, Aules-Curicama LS, Cevallos-Villacis CD, González E, Jima Gavilanes D, Horowitz R, Simancas-Racines D. The Oral-Gut-Immune-Nutrition Axis in Rheumatoid Arthritis: Molecular Mechanisms and Therapeutic Implications. Int J Mol Sci. 2026 Mar 4;27(5):2385.
Yang Y, Wang G, Song Y, Ma J, Liu A. Immunomodulatory effects of oral microbiota in the pathogenesis of rheumatoid arthritis. Front Immunol. 2026 Feb 6;17:1707949.
Tao K, Yuan Y, Xie Q, Dong Z. Relationship between human oral microbiome dysbiosis and neuropsychiatric diseases: An updated overview. Behav Brain Res. 2024 Aug 5;471:115111.
Zhang X, Zhong M, Li Y, Wang H, Xi G, Wang F, Cheng C, Shi Y. Oral microbiota and central nervous system diseases: A review. Neuroprotection. 2025 Feb 9;3(1):79-94.
Tien Vo TT, Tsai MH, Cheng CY, Wang YL, Lee WJ, Lee IT. The oral microbiome-redox-inflammation axis in neurodegeneration: mechanistic insights and therapeutic perspectives. Biochem Pharmacol. 2025 Dec;242(Pt 3):117362.
Zhang W, Li S, Cui Y, Lun Y, Liu H, Li M. The Interface of Oral and Brain Health: Current Insights Into the Bidirectional Relationship Between Alzheimer’s Disease and Periodontitis. CNS Neurosci Ther. 2026 Jan;32(1):e70754.
Pravin V, Vellapandian C, Naveen Kumar V. The oral-gut-brain axis in periodontitis: microbial signaling in systemic and neuroinflammatory disease. Brain Res. 2026 Mar 15;1875:150168.
Miranda V, Laarej K, Cavaliere C. The Oral Microbiota: Implications in Mucosal Health and Systemic Disease-Crosstalk with Gut and Brain. Cells. 2026 Jan 4;15(1):82.
Al-Maweri SA, Al-Mashraqi AA, Al-Qadhi G, Al-Hebshi N, Ba-Hattab R. The association between the oral microbiome and hypertension: a systematic review. J Oral Microbiol. 2025 Feb 2;17(1):2459919.
Pizziolo PG, Silva-Lovato CH, Clemente LM, Carandina A, Salgado HC, Silva TMD, Tobaldini E, Montano N, Ribeiro AB. The role of oral microbiota and tooth loss in cardiovascular disease risk: a systematic review. Biofouling. 2025 Aug;41(7):649-663.
Chen TP, Yu HC, Lin WY, Chang YC. Bidirectional association between obesity and chronic periodontitis: Inflammatory pathways and clinical implications. J Dent Sci. 2025 Oct;20(4):2021-2025.
Graves DT, Levine MA, Aldosary S, Demmer RT. Understanding the Periodontitis-Diabetes Linkage: Mechanisms and Evidence. J Dent Res. 2026 Jan;105(1):21-30.
Frey-Furtado L, Magalhães I, Sampaio-Maia B, Azevedo MJ. Oral microbiome characterization in oral mucositis patients-A systematic review. J Oral Pathol Med. 2023 Nov;52(10):911-918.
Lin CS, Chen TC, Verhoeff MC, Lobbezoo F, Trulsson M, Fuh JL. An umbrella review on the association between factors of oral health and cognitive dysfunction. Ageing Res Rev. 2024 Jan;93:102128.
Guo ZL, Cui MW, Dong YL, Wang S. The oral microbiome as a regulatory hub for systemic health: a systematic review of mechanistic links and clinical implications. J Oral Microbiol. 2026 Mar 3;18(1):2635233.
Wu YC, Chen BY, Duan SZ. Oral microbiota in cardiovascular health and disease. Front Cell Infect Microbiol. 2026 Jan 2;15:1731845.
Su S, Ni X, Lin Y. The oral-gut microbiota axis in cardiovascular diseases: mechanisms, therapeutic targets, and translational challenges. Front Cell Infect Microbiol. 2025 Sep 25;15:1658502.
当我们谈论人体微生态时,最先想到的往往是肠道。但你是否知道,我们的口腔其实是人体内多样性仅次于肠道的第二大微生物栖息地?
近年来越来越多研究提示,口腔作为人体与外界环境交互的重要门户,栖息着高度复杂和动态的微生物群落,即口腔微生物组。这一微生态系统在维持宿主口腔乃至全身健康有着比想象中更深的联系。
现代人常见的龋病、口臭、牙周炎等问题,很多时候都不是某一种“坏菌”突然出现,而是原本受控的微生态关系发生倾斜。
想真正理解口腔菌群,关键不只是记住有哪些细菌,而是理解它如何定植、如何维持平衡,又如何在失调时影响口腔与全身。
本文将从口腔微生物组的组成、建立过程、共生稳态与失调机制出发,帮助大家建立一个更清晰的理解框架。
编者按:
本文是一篇基于近年综述与代表性研究整理的科普性综述,重点讨论口腔微生物组的构成、稳态与失调,以及其与部分全身疾病之间的研究联系。
需要特别说明的是,本文涉及癌症、认知衰退与痴呆、类风湿性关节炎、糖尿病等内容时,所引用的许多证据目前仍以观察性研究和相关性研究为主,不能简单理解为直接因果关系。不同主题的证据成熟度并不一致,相关机制也仍在持续研究中。
全文分上下两部分,本文为上篇;下篇将进一步讨论口腔微生物组与系统性疾病的研究进展、潜在机制、干预策略及临床意义。
从生态学视角来看,口腔微生物组是一个由细菌、古菌、真菌、病毒和原生动物等多种微生物组成的复杂生态系统。这些微生物以生物膜的形式存在牙面、舌背、黏膜、龈沟等不同部位,与宿主长期共处,并共同维持口腔环境的稳定。
换句话说,口腔微生物并不是偶然闯入的污染者,而是从出生起就开始定植的长期居民。
超700种菌:谁构成了口腔群落的核心骨架?
据估计,一般健康的人类口腔及上呼吸消化道中可发现超过700种细菌。
注:其中约58%已被正式命名,16%虽可培养但尚未命名,另有26%仅作为“不可培养的系统发育型”被识别。
在这些细菌中,厚壁菌门(Firmicutes)、拟杆菌门(Bacteroidetes)、变形菌门(Proteobacteria)、放线菌门(Actinobacteria)、梭杆菌门(Fusobacteria)和螺旋体门(Spirochaetes)是六个最主要的门类,占据了口腔细菌群落的绝大部分。
口腔微观:不同位置,住着不同的微生物
口腔独特的解剖结构,包括非脱落的硬组织表面(如牙齿、修复体)和持续脱落的软组织表面(如颊黏膜、舌、腭),以及流动的唾液,为不同微生物提供了多样化的生态位。这些生态位在物理化学条件(如pH、温度、氧化还原电位、营养可得性)上存在显著差异,从而赋予了口腔“空间异质性”,并塑造了具有高度位点特异性的微生物群。
例如:
口腔核心微生物组
尽管个体间差异巨大,但在健康人群中,不同口腔位点存在着相对稳定的“核心微生物组”,这些核心成员负责维持口腔的功能稳定性和生态平衡。例如,研究发现,在健康个体的多个口腔位点中,Streptococcus mitis、Gemella haemolysans、Granulicatella adiacens 几乎普遍存在。
除了细菌,口腔中还存在“核心真菌组”,其中念珠菌属(Candida)最为常见,其次是枝孢属(Cladosporium)、短梗霉属(Aureobasidium)等。
口腔病毒组(virome)则以噬菌体为主,它们通过调节细菌宿主的种群动态和基因交换,在塑造口腔微生物群落结构中发挥着关键作用。
古菌(archaeome)虽然丰度较低,但产甲烷菌如Methanobrevibacter oralis在牙周炎等病理状态下丰度会增加,提示其可能参与疾病过程。
综上,口腔菌群是一个多物种、多领域、高度结构化和功能化的微生态系统。它与宿主共同进化,形成了一个被称为“共生体”的超级有机体。理解这个微生态系统的组成、结构及其与宿主的相互作用,是揭示其在口腔及全身健康与疾病中作用的基础。
健康的口腔里,微生物群落是如何运转的?
在健康状态下,口腔微生物组与宿主之间维持着“共生”或“健康平衡”的和谐关系。
在这个状态下,多样化的微生物群落通过协同作用,共同执行多种有益功能。它们参与营养物质的初步消化与转化,例如,某些口腔细菌能够代谢膳食中的硝酸盐,产生具有血管舒张和抗菌作用的亚硝酸盐和一氧化氮,这不仅有助于调节血压,还能抑制致病菌的生长。
此外,共生微生物通过“定植抵抗”机制,占据口腔内的生态位,与外来病原体竞争营养和附着位点,从而构成了抵御感染的第一道生物防线。
口腔微生物组与宿主健康之间的复杂相互作用
Dame-Teixeira et al., 2025, 第1章
一旦打破生态平衡,问题悄然出现
这种微妙的平衡可能被多种因素打破,导致向“失调”状态的转变。
年龄、感染、环境因素、生活习惯(如高糖饮食、吸烟、饮酒)以及抗菌药物的不当使用,都会对口腔微生物组施加选择压力,破坏其多样性和稳定性。
例如,高糖饮食会促进产酸菌(如变形链球菌)的过度生长,导致口腔环境酸化,从而引发龋齿。同样,宿主的全身性疾病,如糖尿病、高血压、肥胖等代谢综合征,也会通过改变唾液成分、影响免疫应答等方式,加剧口腔微生物的失调。
近年来,随着高通量技术的发展,我们对口腔微生物组的认识正在迅速深化。自2010年以来,相关研究文献数量呈指数级增长(下图)。
1950年~2023年间,以“口腔微生物组”
为主题的已发表论文数量变化趋势
编辑
Dame-Teixeira et al., 2025, 第1章
图中显示,自2005年左右,尤其是2010年以后,相关研究呈现指数级增长。
这些研究从单纯的物种鉴定(它们是谁?)发展到功能分析(它们在做什么?),并整合宏基因组学、宏转录组学、宏蛋白质组学和代谢组学等多组学数据,逐步帮我们理解整个口腔微生态奠定了证据基础。
这些进展不仅揭示了口腔微生物组的复杂性,也为开发基于调节微生物组的个体化预防和治疗策略提供了新的契机。
人类口腔微生物组的建立是一个从生命最早期开始,贯穿一生的动态过程。它并非与生俱来,而是在出生后通过与环境的持续互动逐步获得和演替的。这一过程受到一系列内在(宿主相关)和外在(环境相关)因素的复杂调控,共同塑造了每个个体独特而又具有共性的口腔微生态系统。
人类从胎儿到老年的生命历程示意图
编辑
改编自 DOI: 10.1007/978-3-031-79146-8_2
生命早期的初始定植
出生后,喂养方式成为塑造口腔微生物组的另一个重要因素。婴儿吃进去的不仅是营养,也是微生态的“种子”与“养料”。
母乳:不仅是营养的来源,更是一个动态的生物系统,含有丰富的微生物、人乳寡糖(HMOs)、免疫球蛋白和抗菌蛋白。母乳喂养能够将母亲的特定菌株(如双歧杆菌属)传递给婴儿,为其肠道和口腔“播种”有益菌 。
HMOs作为益生元,能够选择性地促进有益菌(尤其是Bifidobacterium infantis)的生长,同时抑制病原体的黏附 。因此,与配方奶喂养的婴儿相比,母乳喂养婴儿的口腔微生物组通常具有更高的多样性,其中部分变化被认为更接近健康相关模式。
辅食:随着固体食物的引入,婴儿的口腔微生物组会经历又一次重大转变,多样性显著增加,以适应更复杂的碳水化合物和营养物质。
从婴儿期到成年的演替过程
随着个体的成长,口腔的物理环境也在不断变化,这为微生物的演替提供了新的生态位。其中,牙齿的萌出是一个重要阶段。
► 无牙期:以软组织表面为主
在无牙期,口腔内只有软组织黏膜表面,主要定植的是早期定植菌,例如:
► 约 6–10 个月:第一颗乳牙萌出
大约在6-10个月时,第一颗乳牙的萌出为微生物提供了独特的“非脱落表面”,使得能够形成更稳定、更复杂的生物膜(牙菌斑)。
随着乳牙列的完全建立,口腔微生物的多样性显著增加,例如下列菌群开始繁盛:
编辑
► 混合牙列期与恒牙期:厌氧菌定植条件增强
进入混合牙列期和恒牙期,口腔的解剖结构变得更加复杂,为更多厌氧菌的定植创造了条件。
牙龈沟的形成提供了一个低氧、富含蛋白质(龈沟液)的微环境,特别适合严格厌氧和蛋白水解菌(如牙周病相关菌群)的生长。
在这一过程中,微生物群落的组成从以革兰氏阳性兼性菌为主,逐渐向包含更多革兰氏阴性厌氧菌的成熟群落演替。
► 青春期:激素变化驱动的第二次转变
青春期是另一个重要的转变阶段。性激素(如雌二醇、孕酮和睾酮)水平的急剧上升,会直接或间接地影响口腔微环境。
这些激素可以被某些口腔细菌利用作为生长因子,同时也会改变牙龈组织的血管通透性和炎症反应,导致所谓的“青春期牙龈炎” 。
这期间,龈下微生物组中与牙龈炎相关的菌群丰度会增加,例如:
成人口腔内不同生态位及其优势细菌类群
编辑
DOI: 10.1007/978-3-031-79146-8_2
牙菌斑和舌部具有高微生物多样性,而口腔黏膜的多样性相对较低。
影响微生物组获得与建立的其他因素
除了上述生理阶段的变化,还有多种因素持续塑造着口腔微生物组的构成。
✦遗传与宿主防御:身体“筛选”哪些菌容易留下
宿主的遗传背景决定了其免疫系统的特征,如唾液中抗菌蛋白(溶菌酶、乳铁蛋白等)的含量和活性、主要组织相容性复合体(MHC/HLA)的类型等。这会影响哪些微生物更容易定植下来,并长期保留下来。
例如,唾液淀粉酶基因(AMY1)拷贝数较高的人,其唾液中普雷沃菌属和卟啉单胞菌属的水平也更高。
✦社会与环境接触:微生物在人与人/动物之间流动
口腔微生物会通过接触持续传播。
婴儿早期主要受母亲影响,随着成长,兄弟姐妹、照料者、宠物和集体环境的作用也会逐渐增强。
编辑
✦药物使用:抗生素改变菌群起跑线
抗生素的使用,尤其是在生命早期,会对口腔微生物组的建立产生深远影响。
广谱抗生素会无差别地杀灭有益菌和有害菌(好坏一起杀),可能导致菌群失调,并为机会致病菌(如念珠菌)的过度生长创造条件。
✦口腔卫生与生活方式:直接改变牙菌斑与口腔环境
口腔卫生习惯(如刷牙、使用牙线)直接影响牙菌斑的量和组成。
不良的口腔卫生会导致菌斑堆积,促进厌氧菌和致病菌的生长。吸烟和饮酒等生活方式因素也会通过改变口腔的氧化还原电位、pH值和免疫反应,导致微生物组失调,增加牙周病和癌症的风险。
✦成年稳定与老年再变化:从平衡到更易失衡
进入成年期后,健康口腔微生物组通常进入相对稳定阶段。
然而,随着年龄的增长,生理性衰老(如唾液流速下降、免疫功能减退)以及慢性疾病和多重用药的累积效应,会再次改变这种平衡,可能导致机会性病原体(如肠杆菌科、葡萄球菌属)的丰度增加,从而增加老年人患口腔及全身性疾病的风险。
总的来说,口腔微生物组并非一成不变,而是在多种因素作用下不断调整。接下来,需要进一步理解它与宿主之间如何形成并维持这种共生平衡。
经过数千年的共同进化,人类与栖居其身的微生物群落形成了一种深刻的互利共生关系。种共生关系主要通过微生物稳态和宿主稳定性的相互支撑来实现。
微生物稳态:群落内部的协作与拮抗
在健康状态下,口腔微生物并不是各自为战,而是形成了一个相互作用网络,这个网络让菌群具备两种能力:
这种稳态主要靠两类关系共同维持:互利协作与相互拮抗。
↗互利关系:代谢协作与交叉喂养
口腔微生物的主要营养来源于宿主饮食、唾液和龈沟液中的大分子,如糖蛋白和蛋白质。
单个微生物菌种往往“吃不完整”,不具备完全降解这些复杂底物的能力,因此需要不同物种之间通过酶促互补进行代谢合作。这种现象被称为“交叉喂养”,即一个物种的代谢产物成为另一个物种的营养底物。
——典型例子:产酸菌 × 耗酸菌
编辑
这个协作带来两点好处:
整体上,这类代谢网络会提升群落效率,增强了其稳定性。
↘拮抗关系:竞争与抑制
口腔生态位很拥挤,共生菌为了守住地盘,也会用各种方式压制竞争者,尤其是潜在致病菌。这些拮抗作用是维持微生物平衡的关键。
——产生细菌素:定向“抑菌武器”
许多共生菌能够产生小分子的抗菌肽,即细菌素,它们可以特异性地杀灭或抑制亲缘关系较近的细菌。
例如,唾液链球菌(Streptococcus salivarius)产生的Salivaricin(一种细菌素)可以抑制引起咽喉炎的化脓性链球菌(S. pyogenes)。
从无龋个体中分离出的Streptococcus dentisani也能产生细菌素,有效抑制变形链球菌(S. mutans)和具核梭杆菌(F. nucleatum)等致病菌的生长。
——产生过氧化氢:用“氧化压力”压制对手
多种共生链球菌(如S. sanguinis, S. gordonii, S. oralis)能够通过丙酮酸氧化酶产生过氧化氢。
过氧化氢(H₂O₂)对许多无法有效应对氧化应激的厌氧致病菌(如P. gingivalis)和产酸菌(如S. mutans)具有强大的抗菌活性。这种机制在早期菌斑形成和应对糖冲击时尤为重要。
——产碱缓冲 pH:把酸中和掉
除了交叉喂养,一些细菌还能通过代谢氨基酸(如精氨酸)和尿素来产生氨(NH₃),从而中和酸性产物,提高局部pH值。
例如,精氨酸脱亚胺酶系统(ADS)在多种链球菌和放线菌中存在,它降解精氨酸产生氨,不仅能缓冲pH,其代谢过程本身还会消耗精氨酸,从而抑制依赖精氨酸的S. mutans的毒力表达。
——群体感应信号的干扰
微生物通过释放和感知被称为“自诱导物”的信号分子来进行种内和种间的通讯,这一过程称为群体感应(QS)。QS系统调控着生物膜形成、毒力因子产生等多种群体行为。
一些共生菌能够干扰致病菌的QS信号。例如,部分S. gordonii菌株可以抑制S. mutans的CSP信号肽,从而削弱其形成生物膜和产生细菌素的能力。
口腔健康条件下微生物间的相互作用机制
编辑
doi: 10.1007/978-3-031-79146-8_3
维持口腔稳态的关键:定植抵抗与免疫稳态
宿主与口腔微生物组之间的共生关系,不仅体现在微生物群落内部的自我调节,更体现在其与宿主免疫系统的协同作用上,共同维持宿主的稳定与健康。
◉ 定植抵抗:常驻菌先占位,外来菌难落脚
健康的常驻微生物群通过“占位”效应,物理性地占据了口腔黏膜和牙齿表面的附着位点,从而阻止了外来病原体的入侵和定植。这种现象被称为“定植抵抗”。
它主要通过两种方式实现:
这种由微生物组提供的天然屏障,是宿主抵御感染的第一道防线。
◉ 免疫稳态与耐受:不是不管,而是受控共存
口腔黏膜持续暴露于大量的微生物及其抗原之下,但通常不会引发剧烈的炎症反应,这表明宿主免疫系统已经进化出一种与共生微生物和平共处的机制,即免疫耐受。
这种耐受并非免疫系统的“无视”,而是一种主动的、精细调控的平衡状态:既控制微生物,又避免过度炎症伤害自己。
共生菌如何帮助维持这种平衡?
共生微生物的某些分子信号(MAMPs,可理解为“微生物特征信号”)会被宿主的模式识别受体识别(如Toll样受体,TLRs),持续地“教育”和“校准”宿主的免疫系统。在健康状态下,这种相互作用倾向于诱导一种“促耐受”的免疫环境,主要体现在:
——诱导调节性T细胞(Treg)
共生菌及其代谢产物(如短链脂肪酸)可以促进Treg细胞的分化。
Treg细胞通过分泌如IL-10、TGF-β等抗炎细胞因子,来抑制过度的炎症反应,防止对共生菌产生有害的免疫攻击。
——促进分泌型IgA(sIgA)产生
sIgA是黏膜免疫的主要抗体。它能够“包裹”微生物,阻止其黏附到上皮细胞表面并侵入组织,同时不激活强烈的炎症反应。
这种机制常被理解为温和的免疫排斥:既能控制微生物数量,又能避免组织损伤。
——强化上皮屏障
共生菌能够刺激上皮细胞产生黏蛋白和抗菌肽(AMPs),增强黏膜的物理和化学屏障功能,进一步限制微生物与宿主免疫细胞的直接接触。
口腔微生物群与宿主免疫系统的相互作用机制
编辑
doi: 10.1007/978-3-031-79146-8_3
口腔微生态具备定植抵抗、免疫耐受和屏障防御等多重稳定机制,但并不是建立起来就能一直维持下去,真正日复一日改写这套系统的,往往是那些看似普通的生活因素——尤其是饮食。下一章我们来具体了解。
人类饮食变迁与口腔微生物组的共同进化
🪨 旧石器时代
高纤维、低精制糖
→ 菌群多样性更高,龋病较少
🌾 新石器时代
谷物进入主食,碳水摄入增加
→ 产酸相关菌增多,龋病开始上升
🏭 工业革命至今
精制糖和加工食品大量增加
→ 产酸、耐酸菌占优势,微生态更易失衡
人类历史进程中饮食结构的变化与口腔微生物组的演变
编辑
doi: 10.1007/978-3-031-79146-8_4
宏量营养素与微量营养素调控口腔微生态
食物中的宏量营养素(碳水化合物、蛋白质、脂肪)和微量营养素(维生素、矿物质)通过多种途径影响口腔微生物组和宿主反应。
◯ 碳水化合物:一把双刃剑
碳水化合物是口腔微生物最主要的能量来源,但其类型和加工程度决定了其对口腔健康的影响。
——可发酵碳水(尤其精制糖/蔗糖):更易推动失调
可发酵碳水化合物,特别是蔗糖等精制糖,能被口腔中的糖酵解细菌(如变形链球菌、乳杆菌)迅速代谢,产生大量有机酸(主要是乳酸),导致牙菌斑pH值急剧下降。
频繁的酸攻击会破坏牙釉质的矿物质平衡,引发脱矿,最终形成龋洞。
——蔗糖:不仅产酸,还让菌斑黏更厚难清除
变形链球菌还能利用蔗糖合成不溶性的胞外多糖(EPS),这些黏性的葡聚糖构成了生物膜的骨架,不仅促进了细菌的黏附和聚集,还形成了一个扩散屏障,使酸性产物更难被唾液中和,从而加剧了局部的酸性微环境。
——复杂碳水/膳食纤维:整体更友好
复杂碳水化合物,如全谷物、蔬菜和豆类中富含的膳食纤维,对口腔健康则更为有益。这些纤维不易被口腔细菌直接发酵,因此不会引起pH的剧烈下降。
更重要的是,富含纤维的食物通常需要更多的咀嚼,这能刺激唾液分泌,增强唾液的冲刷和缓冲作用,有助于清除食物残渣和中和酸性物质。
编辑
◯ 脂肪和蛋白质:来源与平衡是关键
脂肪和蛋白质对口腔微生物组的影响相对复杂,且与其来源密切相关。
——脂肪:不同类型,炎症倾向不同
饱和脂肪和反式脂肪,常见于加工食品、动物脂肪和部分植物油中,被认为是促炎性的。
不饱和脂肪酸,特别是来自鱼油、坚果和橄榄油的ω-3多不饱和脂肪酸,具有抗炎特性。
——蛋白质:既是宿主营养成分,也可能被某些致病菌利用
蛋白质是构成唾液和龈沟液的重要成分,也是许多蛋白水解型牙周致病菌(如P. gingivalis)的主要营养来源。
在牙周炎状态下,富含蛋白质的龈沟液为这些病原体的生长提供了有利条件。
然而,饮食中的蛋白质来源同样重要。有研究指出,过量摄入动物蛋白可能与更高的炎症水平相关,而植物蛋白则可能更有益于健康。
编辑
doi: 10.1007/978-3-031-79146-8_4
◯ 微量营养素:免疫与修复的调节剂
维生素和矿物质等微量营养素虽然需求量小,但在维持口腔组织完整性和调节免疫功能方面发挥着不可或缺的作用。它们的缺乏会直接损害口腔健康。
——维生素C(抗坏血酸)
是胶原蛋白合成所必需的,对维持牙龈和牙周膜的结构完整性至关重要。
严重缺乏维生素C会导致坏血病,其典型症状之一就是牙龈出血和牙齿松动。
维生素C还是一种重要的抗氧化剂,有助于中和炎症过程中产生的活性氧(ROS),保护牙周组织免受氧化损伤。
——维生素D
在钙磷代谢和骨骼健康中起着核心作用。它能促进牙槽骨的矿化,并具有重要的免疫调节功能,可以抑制过度的炎症反应。
维生素D缺乏与牙周炎的严重程度增加和骨质疏松相关。
——B族维生素
参与多种细胞代谢过程,其缺乏可导致多种口腔黏膜疾病,如舌炎、口角炎和复发性阿弗他溃疡。
——抗氧化剂
抗氧化剂(维生素A、E、类胡萝卜素、多酚等)广泛存在于水果、蔬菜、茶和红酒中。这些物质能够清除自由基,减轻氧化应激,从而保护细胞免受损伤,对预防牙周炎和口腔癌具有积极意义。
综上所述,饮食不仅是口腔疾病(尤其是龋病和牙周炎)发生的重要风险因素,也是维持口腔微生态平衡和宿主健康的关键调控手段。一个均衡的、富含全食品、纤维、维生素和抗氧化剂,同时限制精制糖和饱和脂肪的饮食模式,是促进口腔共生、预防疾病的基石。
胃肠道是人体内最庞大、最复杂的微生物生态系统,从口腔延伸至肛门。传统上,口腔和肠道被视为两个相对独立的微生物栖息地,由胃酸、胆汁和物理距离等屏障隔开。然而,近年的研究揭示了两者之间存在着密切而动态的联系,这一联系被称为“口腔–肠道轴”(Oral-Gut Axis)。
通过该轴,口腔微生物及其代谢产物能够影响肠道微生态和宿主健康,反之亦然。理解口腔–肠道轴的机制,有助于我们从更整体的角度看待多种慢性炎症性疾病、代谢性疾病乃至癌症的相关研究线索。
口腔与肠道微生物组的生物地理学特征
——口腔和肠道的菌群,并不一样
在健康状态下,胃肠道沿线的微生物群落表现出显著的体位特异性。每一段消化道的环境都不一样,比如:
这些差异决定了什么样的细菌适合住在哪里。
比如,胃里的环境酸性很强,细菌数量相对较少;而到了结肠,细菌数量就会大幅增加,种类也更加丰富。
胃:约10³-10⁴/mL 结肠:10¹¹-10¹²/g
人体消化道不同部位的微生物群落分布示意图
编辑
doi: 10.1007/978-3-031-79146-8_5
——但它们之间并不是完全隔绝的
尽管存在显著的体位特异性,但在健康个体中,口腔和肠道之间仍存在一定程度的微生物重叠。研究发现,一些常见的口腔细菌,如:
也可能在肠道中被检测到,尽管丰度通常很低。这表明,口腔微生物向肠道的“转位”是一个生理现象。
问题不在于会不会进入,而在于它们进入后能不能存活、定植和扩张。
口腔微生物向肠道的转位机制
——口腔微生物主要通过两条途径到达肠道
编辑
——为什么健康人通常没事?
虽然口腔细菌确实可能到达肠道,但健康人的肠道并不会轻易让它们安家。
在健康状态下,肠道自身的防御机制——包括完整的黏膜屏障、强大的免疫系统(肠道相关淋巴组织,GALT)以及由常驻肠道菌群构成的“定植抵抗”
这些机制会共同作用,限制外来细菌在肠道中长期生存。因此,口腔菌在健康肠道中通常只是匆匆过客。
口腔–肠道轴失调与疾病
当宿主健康状况发生改变时,口腔–肠道轴的平衡可能被打破,导致口腔细菌在肠道中的“异位定植”,这与多种疾病的发生发展密切相关。
✦ 肠道疾病:口腔细菌可能火上浇油
在炎症性肠病(IBD,包括克罗恩病和溃疡性结肠炎)患者中,肠道黏膜屏障功能受损,通透性增加。
研究发现,IBD患者的肠道中,常常能检测到更多口腔来源的细菌,例如:
具核梭杆菌不仅在口腔中发挥作用,还可能通过口腔–肠道轴影响全身健康。
编辑
<来源:谷禾口腔菌群检测数据库>
动物实验也提供了进一步证据。
如果把患有牙周炎小鼠的口腔菌群,移植给无菌小鼠,后者会出现更明显的肠道炎症,尤其是与Th17细胞相关的免疫反应增强。而这种反应,正是IBD的重要发病机制之一。
提到口腔细菌与疾病的关系,具核梭杆菌(F. nucleatum)是一个非常典型的例子。
研究发现,具核梭杆菌在结直肠癌(CRC)组织中高度富集,并通过其黏附素FadA与肿瘤细胞的E-cadherin结合,进一步激活Wnt/β-catenin信号通路,促进肿瘤增殖。
简单说,就是它可能帮助肿瘤细胞长得更快、扩增得更多。这也说明,某些原本生活在口腔里的细菌,一旦进入异常环境,可能不只是旁观者。
✦ 肝脏疾病:口腔细菌也可能影响肝脏
在肝硬化和非酒精性脂肪性肝病(NAFLD)患者中,也观察到肠道中口腔细菌的富集。
这可能与肝功能受损导致胆汁酸代谢异常和肠道屏障功能下降有关。口腔细菌及其产物(如LPS)通过门静脉系统进入肝脏,可能加剧肝脏的炎症和纤维化进程。
✦ 系统性炎症性疾病:可能牵动全身免疫
在类风湿关节炎(RA)等自身免疫病中,口腔–肠道轴也可能扮演了重要角色。
牙周炎是类风湿关节炎的已知风险因素。口腔病原体(如P. gingivalis)不仅能在口腔局部诱导产生针对瓜氨酸化蛋白的抗体(ACPA),还可能转位至肠道,进一步扰乱肠道免疫系统,加剧全身性自身免疫反应。
口腔中的慢性炎症,可能通过肠道这个“中转站”,放大成全身性问题。
✦ 口腔+肠道菌群:可能提示病情轻重
研究通过对唾液和粪便样本的微生物组成进行分析将牙周炎患者分为不同的口腔-肠道谱型。
牙周炎患者的口腔-肠道微生物谱聚类分析
编辑
doi: 10.1007/978-3-031-79146-8_5
某些谱型与更严重的临床症状(如高比例深牙周袋、牙齿缺失)相关,提示口腔与肠道微生物的特定组合可能预示着疾病的严重程度。
临床意义与展望
口腔–肠道轴的发现,为我们从一个更宏观、更系统的角度理解健康与疾病提供了新的框架。它强调了口腔健康对于维持肠道微生态平衡和预防相关疾病的重要性。
临床上,牙周炎患者的肠道微生物组中,与牙周病相关的口腔类群数量明显更高,且某些口腔菌(如F. nucleatum)在粪便中的丰度与牙周病的严重程度相关。
这提示,粪便中的口腔微生物特征可能成为评估口腔健康状况和预测相关系统性疾病风险的非侵入性生物标志物。
近年来,口腔微生物组的一个特定代谢功能受到了科学界的广泛关注:
将膳食中的硝酸盐(NO₃⁻)
还原为亚硝酸盐(NO₂⁻)
这一过程不仅是口腔微生态系统内部复杂代谢网络的一部分,更通过一个被称为“肠-唾液循环”的途径,对人体的口腔及全身健康产生深远的有益影响。
口腔细菌不仅是潜在的致病者,更是维持宿主生理稳态的重要合作伙伴。
硝酸盐的来源、肠–唾液循环
硝酸盐是自然界中广泛存在的含氮化合物,我们日常饮食中的绿叶蔬菜,如菠菜、生菜、芝麻菜、甜菜根,是其最主要的来源。
当我们摄入这些富含硝酸盐的食物后,约25%的硝酸盐会被肠道吸收进入血液循环。随后,一个有趣的循环开始了:血液中的硝酸盐被唾液腺主动摄取并浓缩,其浓度可比血浆高出10倍以上,然后随唾液分泌回口腔。
编辑
在口腔中,这些来自饮食和唾液的硝酸盐,被特定的口腔细菌利用。这些细菌拥有效率极高的硝酸盐还原酶,能将硝酸盐(NO₃⁻)还原为亚硝酸盐(NO₂⁻)。随后,含有亚硝酸盐的唾液被吞咽,进入胃肠道。在胃的酸性环境中,亚硝酸盐可以被非酶促地转化为一氧化氮(NO)和其他活性氮物种(RNS)。
部分未被转化的亚硝酸盐被小肠吸收进入血液,成为全身NO的循环储备库。最终,血液中大部分的NO和亚硝酸盐会被重新氧化为硝酸盐,再次进入肠-唾液循环。这一完整的通路确保了口腔和全身持续获得由微生物介导产生的亚硝酸盐和NO。
硝酸盐-亚硝酸盐-一氧化氮通路在口腔和全身健康中的作用示意图
编辑
Washio and Takahashi, 2025
这个关键步骤,人体自己做不了→依赖菌群
这里有一个很重要的点:
哺乳动物细胞自身并不擅长把硝酸盐变成亚硝酸盐,因此,这一关键步骤完全依赖于口腔中的微生物。
研究表明,口腔中的亚硝酸盐产生能力存在显著的个体间差异,这主要取决于口腔微生物组中“亚硝酸盐产生菌”的数量和活性。
口腔中的亚硝酸盐产生菌
通过琼脂覆盖培养法等技术,研究人员从唾液、牙菌斑和舌苔中分离并鉴定出多种具有硝酸盐还原能力的细菌。这些细菌大多是口腔中的常见共生菌,而非罕见物种,这表明亚硝酸盐的产生是健康口腔微生态的一项普遍功能。主要的亚硝酸盐产生菌包括:
例如奈瑟菌属,是口腔微生物组中的一种重要细菌,在硝酸盐还原过程中发挥关键作用。通过谷禾口腔菌群检测报告,可以了解到其具体丰度及其在口腔微生物组中的表现,部分奈瑟菌属如下:
编辑
<来源:谷禾口腔菌群检测数据库>
——舌苔后部是“核心工作区”
这些菌主要定植在舌背后部的低氧区域,这里为硝酸盐还原酶的活性提供了理想环境。研究发现,舌苔的亚硝酸盐产生活性通常高于牙菌斑,凸显了舌在这一通路中的核心地位。
编辑
持续摄入富含硝酸盐的饮食,不仅能提供底物,还能作为一种益生元,选择性地增加口腔中硝酸盐还原菌(如奈瑟菌属和罗氏菌属)的丰度,从而正向增强整个系统的亚硝酸盐生成能力。
亚硝酸盐对口腔健康有什么好处?
口腔微生物产生的亚硝酸盐及其下游产物一氧化氮,对维持口腔健康具有多重益处,尤其是在预防两大主要口腔疾病——龋齿和牙周炎方面。
◉ 预防龋齿
亚硝酸盐的防龋机制是多方面的:
——直接抗菌作用
亚硝酸盐在酸性条件下可质子化形成亚硝酸(HNO₂),后者能穿透细菌细胞膜,干扰其能量代谢和关键酶的功能。这对主要的致龋菌——变形链球菌(S. mutans)具有显著的抑制作用。
——抑制产酸
糖吃进去后,细菌产酸的能力可能会被部分压住。
龋齿的一个关键过程,就是细菌分解糖分后产酸,使口腔环境变酸,进而腐蚀牙齿。亚硝酸盐能够抑制牙菌斑微生物的糖酵解过程,从而减少有机酸的产生,减缓菌斑pH值的下降。研究表明,摄入富含硝酸盐的蔬菜汁可以显著缓冲食糖后的口腔酸性环境。
龋齿的发生与口腔内多种细菌的活动密切相关,通过监测口腔内的龋齿相关菌群丰度及是否超标,可以更好地掌握口腔微生物组的动态变化。
编辑
<来源:谷禾口腔菌群检测数据库>
——帮助菌群恢复平衡
亚硝酸盐对不同细菌的敏感性不同,它能抑制产酸/耐酸菌的生长,同时可能对硝酸盐还原菌影响较小或无影响。这种选择性压力有助于将失调的、致龋的微生物组向更健康的、共生的方向逆转。
——与乳酸代谢的生态协同
韦荣菌属等细菌在利用乳酸的同时进行硝酸盐还原。这一过程既消耗了致龋的乳酸,又产生了具有保护作用的亚硝酸盐,形成了一个精妙的负反馈调节环路,有助于维持口腔pH稳态。
◉ 预防牙周炎
亚硝酸盐和一氧化氮(NO)对牙周健康同样有益。
首先,它们对多种主要的牙周致病菌,如牙龈卟啉单胞菌(P. gingivalis)、中间普雷沃菌(P. intermedia)、具核梭杆菌(F. nucleatum),也具有抗菌活性。
其次,NO作为一种重要的信号分子,具有强大的血管舒张和抗炎作用。通过改善牙龈组织的微循环血流,NO有助于输送营养和免疫细胞,清除代谢废物,从而促进牙龈炎症的消退和组织修复。临床研究已表明,摄入富含硝酸盐的莴苣汁可以减轻慢性牙龈炎患者的牙龈炎症症状。
亚硝酸盐的影响,不止停留在口腔
通过肠-唾液循环,口腔微生物组的硝酸盐代谢功能超越了口腔局部,对全身健康,特别是心血管系统,产生了系统性的积极影响。
最核心的作用是调节血压
由口腔产生的亚硝酸盐进入循环系统后,在缺氧组织中被有效还原为NO。NO是内皮细胞产生的一种关键的血管舒张因子,能够松弛血管平滑肌,降低外周血管阻力,从而降低血压。
大量临床试验证实,补充膳食硝酸盐(如饮用甜菜根汁)能够显著降低高血压患者的血压。
编辑
一个很关键的证据:抗菌漱口水打断这条通路
重要的是,当使用抗菌漱口水清除口腔中的硝酸盐还原菌后,这种降压效应会明显减弱甚至消失,这证明了口腔微生物组在这一过程中的核心地位。因此,一个功能活跃的口腔硝酸盐还原菌群,可以作为内源性NO生成途径(依赖内皮NO合酶)的一个重要补充,尤其是在衰老或疾病状态下内皮功能受损时。
也就是说,口腔微生物组的功能状态,可能真的会影响全身血压调节。
其他潜在好处也在被关注
NO还具有改善运动表现、保护胃黏膜、增强认知功能等多种潜在益处。
例如,补充硝酸盐可以降低运动时的耗氧量,提高肌肉收缩效率,从而提升耐力运动表现。
那硝酸盐会不会有风险?
很多人会担心一个问题:硝酸盐和亚硝酸盐会不会形成致癌物,比如亚硝胺?
这个担忧曾经非常常见。
然而,目前的主流科学观点认为,对于通过蔬菜摄入的硝酸盐,这种风险极低。因为蔬菜中富含维生素C和多酚等抗氧化剂,能够有效抑制亚硝胺的形成。因此,从健康角度看,摄入富含硝酸盐的蔬菜所带来的益处远大于其潜在风险。
总之,口腔微生物组的硝酸盐代谢功能是宿主-微生物共生关系的一个典范。它不仅通过局部抗菌和pH调节作用维护口腔健康,还通过肠-唾液循环系统性地贡献于心血管健康。
这也带来了一个很有意义的启发:
将硝酸盐视为一种重要的益生元,将硝酸盐还原菌视为潜在的益生菌,为通过饮食和微生态干预来预防和管理慢性疾病提供了新的思路。
唾液是口腔环境的命脉,它不仅仅是水的简单混合物,而是一种复杂的生物流体,富含多种有机和无机成分。
在维持口腔微生物组与宿主之间的共生关系中,唾液扮演着不可或缺的守护者角色。它通过机械清洁、免疫防御、缓冲调节和组织保护等多重机制,主动塑造和调控着口腔微生态的平衡,是抵御口腔及全身感染的第一道关键防线。
唾液的物理与机械功能
冲洗与清洁
我们每天都在不断分泌和吞咽唾液,伴随着吞咽动作,唾液能够有效地将食物残渣、脱落的上皮细胞以及未牢固附着的微生物从口腔中清除,这一过程被称为“唾液清除”。
据估计,一个健康成年人每天仅通过吞咽唾液,
就能清除掉大约 800亿(8×10¹⁰)个细菌!
唾液流速是关键
高流速的唾液能更快地稀释和冲走食糖后产生的酸,以及减少细菌在牙面的停留时间,从而降低龋病的风险。相反,唾液分泌减少(口干症)的患者,其口腔细菌载量通常更高,患龋齿、牙周病和口腔念珠菌病的风险也显著增加。
润滑口腔,帮助吞咽和说话
除了冲洗功能,唾液还是口腔的重要润滑剂。
唾液中的黏蛋白等糖蛋白赋予了唾液黏弹性,使其能够在口腔软硬组织表面形成一层润滑膜。这层膜有几个非常实际的作用:
唾液作为口腔“守护者”的多重功能
编辑
DOI: 10.1007/978-3-031-79146-8_7
唾液的化学防御:不让口腔变酸
缓冲pH调节
细菌导致蛀牙的一个核心武器是:产酸。而唾液的一个重要任务,就是把口腔的酸碱度(pH值)维持在中性范围(大约 6.5 – 7.5 之间)。
唾液里自带了一套碳酸氢盐缓冲系统。当我们吃了酸性食物,或者细菌开始大量产酸时,这套系统就会迅速出动,把酸中和掉。
有意思的是,当你咀嚼食物时,唾液流速会加快,里面的碳酸氢盐浓度也会跟着升高,中和酸的能力就变得更强。
此外,一些口腔细菌也能通过分解唾液中的尿素产生氨,辅助提高pH值,共同维持口腔环境的稳定。
唾液和口腔微生物之间并不是简单的“防守与进攻”关系,很多时候更像是一种动态协作。
免疫防御:非特异性与特异性免疫
唾液并不是机械地“冲走细菌”,它本身还含有大量具有抗菌和免疫作用的分子,它们协同作用,构成一个强大的防御网络。
◖溶菌酶:直接破坏细菌细胞壁
溶菌酶能够水解革兰氏阳性菌细胞壁的肽聚糖,导致细菌裂解死亡。它还具有非酶依赖的抗革兰氏阴性菌活性,并能促进细菌聚集,便于清除。
◖乳铁蛋白:不给细菌“吃铁”
乳铁蛋白是一种铁结合蛋白,通过螯合游离的铁离子,剥夺了细菌生长所必需的铁元素,从而发挥抑菌作用(营养免疫)。此外,它还具有不依赖铁的直接杀菌和抗病毒活性。
◖乳过氧化物酶系统:抑制细菌产酸和代谢
该系统利用细菌代谢产生的过氧化氢(H₂O₂)和唾液中的硫氰酸盐(SCN⁻)作为底物,生成具有强氧化性的次硫氰酸(OSCN⁻),后者能抑制多种细菌的糖酵解酶,从而阻断其产酸和生长。
简单来说,就是让细菌“没法高效产酸、没法顺利生长”。
◖组蛋白、抗菌肽:广谱抗菌抗真菌
组蛋白是一类富含组氨酸的阳离子肽,具有广谱的抗真菌(特别是抗白色念珠菌)和抗菌活性。它们通过破坏微生物细胞膜的完整性或干扰其内部代谢来发挥作用。
防御素和 Cathelicidin (LL-37) 这些是小分子的阳离子抗菌肽,主要由上皮细胞和中性粒细胞分泌。它们能像分子飞镖一样插入微生物的细胞膜,形成孔道,导致细胞内容物泄漏而死亡。
◖sIgA:温和但精准的免疫控制者
分泌型免疫球蛋白A (sIgA)是唾液中最主要的抗体,也是黏膜适应性免疫的核心。
它的特别之处在于:不是通过强烈炎症来“消灭一切”,而是通过更温和的方式控制微生物。
sIgA能够特异性地结合入侵的微生物和毒素,通过免疫排斥机制阻止它们黏附到宿主细胞表面,并促进它们以聚集体的形式被唾液清除。sIgA在不引发强烈炎症反应的情况下控制微生物,是维持黏膜稳态的关键。
唾液中的非特异性与特异性免疫防御
编辑
DOI: 10.1007/978-3-031-79146-8_7
唾液会给口腔表面加上一层保护膜
唾液中的蛋白质和糖蛋白(如黏蛋白、富含脯氨酸蛋白、软蛋白等)会选择性地吸附在牙齿和口腔黏膜表面,形成一层厚度仅为几百纳米的无细胞有机薄膜,分别称为获得性釉质膜和黏膜膜。
这层唾液膜具有双重功能:
它作为一道物理屏障,保护牙釉质免受酸的直接侵蚀,减缓脱矿过程。同时,膜中的钙结合蛋白(如软蛋白)有助于将钙和磷酸盐离子富集在牙齿表面,促进再矿化(相当于帮助牙齿进行微小损伤的自我修复)。对于软组织,黏膜膜则提供了润滑和保湿,防止干燥和机械损伤。
这层膜还有一个非常有意思的生态学功能:
它会影响微生物最初如何在口腔表面定植。
不同的唾液蛋白会选择性地结合不同的细菌,相当于为特定微生物提供落脚点。这意味着,唾液膜并不是单纯被动地让细菌附着,而是在一定程度上选择性地决定谁先成为第一批定植者。
例如,α-淀粉酶可以结合某些链球菌,而富含脯氨酸蛋白则为另一些细菌提供受体。
而最早附着的那批细菌,往往会影响后续整个牙菌斑生物膜的形成顺序和组成。
总的来说,唾液是维持口腔微生态平衡的重要保障。可一旦唾液分泌减少或功能受损,原本稳定的菌群关系就可能被打破,口腔也会由健康走向失衡。接下来,我们就看看口腔微生态失调是如何发生的。
在健康状态下,口腔微生物组与宿主维持着一种动态的共生平衡。然而,当这种平衡被宿主、环境或微生物自身的因素打破时,就会导致失调。
失调并非简单地指有害细菌的增加,而是一个更复杂的生态学概念,指微生物群落的组成、结构和/或功能发生有害的改变,从而增加了宿主患病的风险。
口腔微生态失调是龋病、牙周炎等主要口腔疾病的核心驱动力,并与多种系统性疾病密切相关。
失调的生态学假说
理解口腔失调的关键在于“生态学假说”(Ecological Plaque Hypothesis)。该假说由Philip Marsh提出,这个假说认为,口腔疾病并不是单纯由菌太多引起,也不只是由某一种坏菌单独造成,而是局部环境发生改变后,整个微生物群发生了致病的重组。
简单来说,它强调三件事:
1
疾病的本质是生态失衡
在健康的口腔里,一些潜在致病菌其实本来就可能存在,只是数量很低,而且被共生菌群和宿主防御系统共同压制着。
问题不在于它们有没有,而在于潜在致病菌会不会在合适的环境下突然占上风。
2
环境变化是失调的起点
当口腔局部环境出现明显变化时,比如:
原有的微生物平衡就会被打破。某些原本不占优势的微生物,会因此获得更适合自己的生存条件。
1
菌群演替会推动疾病不断加重
一旦新的环境开始偏向某些致病相关微生物,它们就会逐渐增殖,并进一步改变周围环境。
这种改变又会继续压制共生菌、扶持致病菌,形成一个自我强化的恶性循环,最终推动疾病发生和进展。
龋病和牙周炎,就是两个典型例子。
——在龋病中:高糖让口腔变酸
如果长期频繁摄入糖分,口腔中的细菌就会不断把糖发酵成酸。于是,口腔环境反复酸化。
这种酸性环境抑制了对酸敏感的共生菌(如S. sanguinis),同时促进了耐酸和产酸菌(如变形链球菌和乳杆菌)的生长。这些菌的增殖又会产生更多的酸,使失调状态加剧,最终导致牙齿脱矿,龋病发生。
编辑
——在牙周炎中:缺氧和炎症改变生态位
牙周炎的过程则有所不同。
当菌斑长期堆积,牙龈沟内环境会逐渐变得更缺氧,同时炎症反应会带来更多组织分解产物和龈沟液。
这就为一些厌氧、蛋白水解型细菌创造了有利条件,比如牙龈卟啉单胞菌等。
这些细菌的毒力因子又会进一步刺激和放大宿主炎症反应,造成组织破坏、牙周袋加深,进而为更多厌氧菌提供栖息空间。
于是,牙周炎也进入了“环境改变—菌群演替—炎症加重”的循环。
牙周炎的病理过程不仅涉及到常见的细菌,还包括一些厌氧菌和蛋白水解型细菌的增殖等。这些细菌的丰度和比例变化能综合辅助评估牙周炎的风险。
编辑
<来源:谷禾口腔菌群检测数据库>
口腔微生态失调有哪些表现?
口腔微生物组的失调可以体现在多个层面,这些改变共同构成了疾病的微生物学基础。
◎ 物种组成改变
这是失调最直观的表现。
在龋病中,可见变形链球菌、乳杆菌和部分放线菌的丰度增加。
在牙周炎中,则表现为“红色复合体”(P. gingivalis, T. forsythia, T. denticola)和“橙色复合体”等厌氧菌群的富集,而与健康相关的链球菌、放线菌等共生菌比例下降。
◎ 多样性变化
失调对微生物多样性的影响因疾病而异。
在龋病中,由于强烈的酸性选择压力,微生物多样性通常会降低,少数耐酸菌占据主导地位。
在牙周炎中,由于牙周袋的形成创造了新的生态位,并且炎症反应提供了更丰富的营养,微生物多样性反而会增加。
◎ 功能代谢改变
失调不仅是物种的替换,更是整个群落功能的转变。
在龋病相关的生物膜中,糖酵解和产酸相关的代谢通路被上调。
在牙周炎相关的微生物组中,则表现为蛋白水解、氨基酸发酵、毒力因子(如LPS、牙龈蛋白酶)合成以及炎症诱导相关通路的富集。
◎ 生物膜空间结构改变
失调也会改变生物膜的物理结构。
例如,在致龋生物膜中,大量的胞外多糖(EPS)形成了一个致密的、酸性的微环境。
在牙周炎中,生物膜结构变得更加疏松,但更具侵袭性,细菌更容易从生物膜中脱落并侵入宿主组织。
这说明,疾病并不只是菌种名单变了,连它们在口腔里如何组织起来也变了。
是什么在推动口腔失调?
多种因素可以作为触发或加剧口腔失调的“生态压力”。
► 宿主因素
饮食:如前所述,高糖饮食是龋病失调的主要驱动力。频繁吃糖,比偶尔一次大量摄糖更容易反复制造酸性环境。
免疫状态:宿主免疫功能低下或免疫反应失调(如过度炎症)会无法有效控制致病菌,从而导致失调。例如,糖尿病患者的高血糖状态和受损的中性粒细胞功能,使其更容易发生严重的牙周炎。
唾液功能:唾液流速降低和缓冲能力下降会削弱口腔的自洁和中和酸的能力,为产酸菌的生长创造有利条件。
激素水平:青春期、妊娠期和更年期的激素波动会改变口腔环境,影响特定微生物的生长,可能暂时性地导致失调。
► 环境与行为因素
口腔卫生不良:不良的口腔卫生导致菌斑大量积聚,是牙龈炎和牙周炎失调的始动因素。
如果清洁不到位,微生物生物膜就会越来越成熟,生态环境也会越来越偏向致病状态。
吸烟与饮酒:吸烟会造成口腔局部缺氧,抑制有益的需氧/兼性厌氧菌,促进厌氧病原体的生长。酒精则可能直接影响微生物代谢并损害黏膜屏障。
药物使用:广谱抗生素会破坏正常的微生物群落,为机会性感染(如念珠菌病)打开大门。其他药物(如抗高血压药、抗抑郁药)可能通过引起口干,而间接导致失调。
►关键病原体:不是数量最多,却可能最会“带节奏”
“关键病原体假说”是生态学假说的延伸,它强调某些低丰度的微生物能够通过不成比例的巨大影响力来操纵宿主反应和微生物群落,从而引发失调。
牙龈卟啉单胞菌(P. gingivalis)是典型的关键病原体。
它虽然在菌斑中数量不多,但能通过其独特的毒力因子(如牙龈蛋白酶)破坏宿主的补体系统和中性粒细胞功能,造成局部免疫抑制。这种“被操纵”的炎症环境既能保护牙龈卟啉单胞菌自身免于被清除,又能为整个厌氧菌群落的生长提供营养(源自组织破坏和炎症渗出),最终导致一个协同致病的失调群落的形成。
它不只是自己受益,还把整个微生态都往致病方向带偏了。
总之,口腔微生态的失调是一个由多因素驱动的、复杂的生态学过程。它不仅仅是“坏细菌”战胜“好细菌”的简单故事,而是整个微生态系统在内外压力下失去稳态、功能转向致病方向的系统性失败。
未完待续,下期我们将继续讨论:
注:本账号内容仅供学习和交流,不构成任何形式的医疗建议。
主要参考文献
Dame-Teixeira N, Do T, Deng D. The Oral Microbiome and Us. Adv Exp Med Biol. 2025;1472:3-9.
Azevedo MJ, Kaan AM, Costa CFFA, Sampaio-Maia B, Zaura E. Acquisition of the Oral Microbiome. Adv Exp Med Biol. 2025;1472:13-29.
de Barros Santos HS, Pagnussatti MEL, Arthur RA. Symbiosis Between the Oral Microbiome and the Human Host: Microbial Homeostasis and Stability of the Host. Adv Exp Med Biol. 2025;1472:31-51.
Santonocito S, Polizzi A, Isola G. The Impact of Diet and Nutrition on the Oral Microbiome. Adv Exp Med Biol. 2025;1472:53-69.
Colombo APV, Lourenço TGB, de Oliveira AM, da Costa ALA. Link Between Oral and Gut Microbiomes: The Oral-Gut Axis. Adv Exp Med Biol. 2025;1472:71-87.
Washio, J., Takahashi, N. (2025). Nitrite Production from Nitrate in the Oral Microbiome and Its Contribution to Oral and Systemic Health. In: Dame-Teixeira, N., Deng, D., Do, T. (eds) Oral Microbiome. Advances in Experimental Medicine and Biology, vol 1472. Springer, Cham.
Heller D, Nery GB, Bachi ALL, Al-Hashimi I. Positive Role of Saliva in the Oral Microbiome. Adv Exp Med Biol. 2025;1472:103-118.

谷禾健康
编辑
在全球范围内,乳腺癌已成为女性发病率最高的恶性肿瘤,同时也是导致全球癌症相关死亡的第四大原因之一,严重威胁着女性的生命健康。为了最大程度降低复发风险、延长生存期,辅助化疗几乎是绝大多数乳腺癌患者的必经之路。然而,化疗在带来生存希望的同时,也常常伴随着一系列令人痛苦的副作用。其中,癌症相关疲劳便是最为常见、也最容易被忽视的问题之一。
对于许多乳腺癌患者来说,化疗的艰辛不仅在于治疗本身,相比于脱发、恶心等常见副作用,更在于一种挥之不去的、深入骨髓的疲惫感。这种疲惫并非休息一下就能缓解,它有一个专业名称——癌症相关疲劳(Cancer-Related Fatigue,简称 CRF)。它正悄然影响着无数患者的生活质量,甚至可能比疾病本身更令人困扰。
很多人会觉得,“化疗累一点不是很正常吗?休息休息就好了。” 但事实上,癌症相关疲劳绝非简单的身体劳累,它有着明确的医学定义。
美国国家综合癌症网络(NCCN)将其描述为一种持续存在、令人痛苦的身体、情感和 / 或认知层面的疲惫感,这种疲惫与近期的活动量不成比,会严重干扰正常生活,比如无法完成家务、难以专注交流、甚至连起床都觉得费力。
更关键的是,这种疲劳很难通过常规休息缓解。哪怕患者睡足 10 小时,依然会觉得浑身无力、精神涣散,这种持续性的消耗,比身体上的疼痛更易击垮患者的心理防线。
研究数据显示,超过90%的化疗患者会经历癌症相关疲劳,而在乳腺癌患者群体中,由于常需要接受多种治疗手段(如手术、放化疗、内分泌治疗等),疲劳的发生率和严重程度更高。高达30%的乳腺癌幸存者在治疗结束后数年,仍被重度疲劳所困扰。
这种长期疲惫不仅限制了她们的身体活动能力,还阻碍了重返工作岗位的步伐,对心理和社交生活造成了深远影响。癌症相关疲劳的危害远不止于主观感受上的不适。研究证实,疲劳的严重程度与患者的生活质量呈显著负相关,甚至可能影响生存结局。
尽管如此,目前医学界对癌症相关疲劳的内在发生机制仍知之甚少,这也导致临床上缺乏足够有效的、基于循证医学的干预手段。传统的治疗手段,如睡眠指导、运动疗法、心理干预等,虽然有一定帮助,但对于中重度疲劳的患者往往效果有限。这也使得寻找有效的治疗靶点、开发针对性干预措施变得尤为迫切。
随着人体“第二基因组”——肠道菌群研究的深入,科学家们开始将目光投向了我们的肠道。肠道中庞大的微生物生态系统不仅在消化吸收中发挥重要作用,更是人体最大的免疫器官,并通过复杂的神经-免疫-内分泌网络与大脑进行双向沟通,形成所谓的”肠-脑轴”。越来越多证据表明,肠道菌群可能在癌因性疲劳的发生发展中扮演关键角色。
化疗药物在杀死癌细胞的同时,往往也会攻击肠道中的有益菌群,导致菌群结构失衡。当肠道菌群失去平衡,肠道这道天然屏障就会受损出现“肠漏”,原本被“关”在肠道内的微生物产物和某些神经活性代谢物质,就可能进入血液循环。这些物质会沿着“肠-脑轴”激活神经、免疫和内分泌等多条通路,促使体内释放出一系列炎性因子和活性物质(比如IL-6、脂多糖等)。这些物质可能引发神经炎症,对中枢神经系统产生影响,最终表现为一种难以缓解的疲惫感。与此同时,这种内在变化还可能影响患者的行为,比如让人不自觉地减少活动、不愿社交,从而进一步加重疲劳的恶性循环。
2026年,发表在国际知名肿瘤学期刊《Frontiers in Oncology》上的一项最新前瞻性队列研究,该研究不仅证实了肠道菌群在化疗疲劳中的核心作用,最新的研究发现,我们或许可以通过分析患者化疗前的肠道菌群特征,提前预测她们在治疗过程中可能出现疲劳的风险。这为早期干预、预防严重疲劳的发生开辟了全新的思路。通过该研究深入探讨肠道菌群如何成为预测化疗疲劳的”生物密码”。
编辑
为了探究肠道菌群与癌因性疲劳之间的真实因果与预测关系,苏州大学的研究团队设计了一项严谨的前瞻性队列研究。与以往只能观察“当前状态”的横断面研究不同,前瞻性研究的特点在于它能预见未来,通过在事件发生前就开始收集数据,而不是回顾过去发生的事情。这样的设计能够避免回忆偏差,更准确地揭示因果关系和预测模型。
研究团队在2021年期间,从两家三甲医院招募了100名即将首次接受辅助化疗(共四个周期)的女性乳腺癌患者。为了确保研究的纯粹性,采用严格的入组标准:患者在入组前均没有精神疾病、胃肠道疾病、血液或内分泌系统疾病,且在过去三个月内未使用过抗生素或益生菌。这些严格的排除标准,旨在最大程度地保证所采集到的肠道菌群样本能反映其“原生态”,排除其他疾病或药物的干扰。所有患者均在开始辅助化疗之前入组,确保可以获得她们的”基线数据“——也就是在化疗前还未对肠道菌群产生影响的状态。采用配对粪便和血液样本,以探索潜在的神经免疫-内分泌途径。
为什么特别关注“第三个化疗周期”?
临床经验和既往纵向研究表明,接受四个周期化疗的乳腺癌患者,其疲劳感往往在第三个周期达到顶峰。此时,化疗药物的累积毒性最大,患者的生理和心理承受力往往处于临界点。
如果在这个阶段出现中重度疲劳,极易导致化疗减量甚至中断,直接影响患者的生存预后。因此,精准预测并干预第三周期的疲劳,具有极高的临床价值。
研究人员在患者化疗前(基线)收集了她们的粪便样本、血液样本,并进行了详细的疲劳量表评估(包括癌症疲劳量表CFS和视觉模拟疲劳量表VAFS)。
1
癌症疲劳量表
癌症疲劳量表(Cancer Fatigue Scale, CFS)由日本学者Okuyama于2000年开发的,广泛用于评估癌症患者疲劳症状,该量表包括躯体疲劳、情绪疲劳、认知疲劳三个维度,共15个条目。每个项目均采用5分制李克特量表(1-5)进行评分,总分范围为0-60。分数越高,表示疲劳越严重。
2
视觉模拟疲劳量表
视觉模拟疲劳量表(Visual Analog Fatigue Scale, VAFS)用于使用具有11个等级的标尺评估患者的疲劳水平,从“我不觉得累”到“我觉得筋疲力尽”。VAFS采用0-10的数值评分。
0表示无疲劳,1-3表示轻度疲劳,4-6表示中度疲劳,7-10表示重度疲劳。
随后,在患者进行到第三个化疗周期时,再次进行随访评估。根据第三周期的VAFS评分,患者被分为两组:
16S rDNA测序和微生物多样性数据分析
该项目所有患者的粪便样本,使用杭州谷禾提供的专用常温保存管进行采集和储存,并在谷禾实验室完成了16S扩增子测序及生物信息分析。
样本采用专用的DNA提取试剂盒(GHFDE100 DNA isolation kit)从粪便样本中提取细菌总基因组DNA,对16S rDNA基因的V4高变区进行PCR扩增。采用Illumina NovaSeq 6000高通量测序平台测序,结合生物信息学分析流程(如VSearch聚类、SILVA数据库注释等),计算得到从门到属各个分类层级的微生物相对丰度。
16S扩增子测序的临床转化价值
在众多微生物组学技术中,16S扩增子测序在临床队列探索中具有不可替代的价值。其优势主要体现在:
研究结果发现:
基线时,Y0组和Y1组在躯体疲劳、认知疲劳、疲劳总评分(CFS评分)和疲劳程度(VAFS评分)方面无显著差异(P >0.05)。
然而,在第三个化疗周期,观察到两组之间在身体疲劳、情绪疲劳、疲劳总评分和疲劳程度方面存在显著差异(P< 0.05),Y1组的所有评分均高于Y0组。而在化疗开始前,那些后来在第三周期发展为中重度疲劳(Y1组)的患者,其肠道菌群结构已经与轻度疲劳(Y0组)患者出现了显著差异。这表明,肠道菌群并非在化疗后才被动改变,而是在治疗开始前就可能决定了个体对化疗毒副反应的易感性。
1. 菌群失衡的早期信号:
B/F比值升高与变形菌门增殖
在门水平上,肠道菌群主要由拟杆菌门 、厚壁菌门 、变形菌门、梭杆菌门 (Fusobacteria) 构成,这四大菌门的总和占比均超过了菌群总数的95%。进一步研究发现,中重度疲劳组患者在化疗前的拟杆菌门/厚壁菌门比值(B/F比值)显著高于轻度疲劳组(1.15 vs 1.03, P = 0.043)。
注:B/F比值长期以来被视为衡量肠道菌群健康状况的宏观指标之一,其升高通常与肥胖、炎症性肠病等多种代谢和炎症性疾病相关,暗示着肠道微生态的失衡。
更值得注意的是,中重度疲劳组患者肠道内的变形菌门丰度显著增加(8.03% vs 5.01%, P = 0.039)。变形菌门被誉为“微生物界的杂草”,其在肠道中的过度扩张是菌群失调的典型标志。变形菌门下包含了大量潜在的致病菌,在大肠杆菌、沙门氏菌、幽门螺杆菌等常见致病菌中都占主导地位,它们大多属于革兰氏阴性菌,其细胞壁外膜富含脂多糖(LPS,即内毒素)。
注:LPS是引发人体全身性炎症反应的强烈触发器,其水平的升高意味着肠道内存在一个潜在的“炎症火药库”。
在菌群多样性方面,两组之间的α多样性无显著差异(Shannon指数P = 0.505,Simpson指数P = 0.820,Chao1指数P = 0.326);β多样性分析也显示两组菌群结构整体相似(ANOSIM及PERMANOVA分析P值均大于0.05),主坐标分析结果见图1。
图1:两组患者肠道菌群的α多样性与β多样性分析
编辑
虽然整体多样性指数(如Shannon, Simpson, Chao1)无显著差异,但PCoA图显示两组样本在空间分布上略有分离趋势,预示着特定菌群的丰度已悄然改变。
2. 产短链脂肪酸菌匮乏
通过LEfSe(线性判别分析效应量)深度挖掘,研究人员发现,轻度疲劳组(Y0组)患者肠道内富含考拉杆菌属(Phascolarctobacterium)和瘤胃菌科(Ruminococcaceae)等有益菌。这些细菌是肠道内著名的产短链脂肪酸菌(尤其是丁酸)。
注:丁酸不仅是肠道黏膜细胞的主要能量来源,能维护肠道屏障的完整性,还能穿透血脑屏障,发挥强大的抗炎和神经保护作用。
相反,中重度疲劳组(Y1组)患者体内这些“护城河”细菌的丰度显著降低,取而代之的是韦荣球菌属(Veillonella)、巨球型菌属(Megasphaera)以及肠杆菌目(Enterobacteriales)等条件致病菌的增多。这些菌群的富集,进一步加剧了肠道微环境的促炎状态(图2)。
图2:LEfSe分析揭示了两组患者在化疗前肠道菌群的特征性差异
编辑
红色代表中重度疲劳组(Y1)富集的菌群(如变形菌门下的γ-变形菌纲、肠杆菌目、肠杆菌科),绿色代表轻度疲劳组(Y0)富集的菌群(如瘤胃菌科下的RFN20、考拉杆菌属)。
3. 构建高精度预测模型:AUC高达0.82
基于上述发现,研究团队将基线肠道菌群特征与临床协变量结合,构建了一个多因素逻辑回归预测模型。这个模型的强大之处在于,它不仅仅依赖于单一指标,而是综合了多个维度的信息,模拟了临床决策的复杂性。
模型变量的协同解读
最终进入模型的变量包括:
◐ 菌 群 变 量
◐ 临 床 变 量
◐ 其 他 临 床 协 变 量
包括年龄、BMI、癌症分期等,尽管这些因素在单变量分析中可能与疲劳有关,但在加入了强大的菌群变量后,它们的独立预测能力被削弱。这反过来证明,肠道菌群可能是连接这些临床因素与疲劳结果的更底层的生物学中介。
结果显示,该模型预测患者在第三周期是否会发生中重度疲劳的准确率(AUC值)达到了0.82(95% CI: 0.73 – 0.90, P < 0.001)。 AUC值是衡量模型预测能力的标准,0.82意味着该模型具有“良好”到“优秀”的区分能力。
在最佳截断值下,该模型的敏感性为82.8%(能正确识别出82.8%的未来高疲劳患者),特异性为71.4%(能正确识别出71.4%的未来低疲劳患者)。这意味着,通过化疗前的一次粪便16S测序,结合简单的疼痛评估,医生就能以较高的准确率找出那些容易在化疗中严重疲劳症的高危患者,从而提前进行干预。
为了弄清肠道里的细菌究竟是如何影响大脑产生疲劳感,研究人员从100人的大队列中随机抽取了13名患者组成子队列:CRF无显著变化组(n=6)和CRF增加组(n=7)。
先导队列(n = 13)与整体队列(n = 100)在社会人口学或临床特征上无显著差异(均为 P > 0.05),验证了随机抽样子样本的代表性。对比了她们化疗前后肠道菌群和血液中神经免疫内分泌指标的动态变化。结果揭示了一条清晰的“肠-脑轴”致病链。
1
屏障破损:产丁酸菌Blautia减少与LPS泄漏
在化疗过程中,随着疲劳感加剧的患者,其肠道内的厚壁菌门,特别是经黏液真杆菌属(Blautia)的丰度出现了显著下降(P = 0.011)。Blautia是肠道内极其重要的产丁酸菌。当它减少时,肠道内短链脂肪酸(SCFAs)的产量锐减。这不仅导致肠道黏膜细胞失去营养支持,发生肠漏,还使得变形菌门产生的内毒素(LPS)趁机大量进入血液循环。相关性分析也证实,在化疗前,Blautia的丰度与血清LPS浓度呈显著负相关(图3)。
图3:肠道菌群与神经免疫内分泌指标
及疲劳评分的相关性热图
编辑
化疗前(a),Blautia与BDNF呈显著正相关,与疲劳评分(CRF)和LPS呈显著负相关。而巨球型菌属(Megasphaera)则与CRF、IL-6和NGF呈正相关,清晰地展示了“好菌”与“坏菌”的不同角色。
2
神经炎症:LPS如何抑制“大脑肥料”BDNF?
——LPS引发的免疫反应与神经炎症
进入血液的LPS会激活免疫系统(如TLR4受体),引发全身性的炎症反应,导致白细胞介素-6(IL-6)等促炎细胞因子释放。这些炎症因子和LPS能够穿透或破坏血脑屏障,引起中枢神经系统的神经炎症。
已有研究表明,在神经炎症模型中,LPS会激活大脑中的小胶质细胞。被激活的小胶质细胞会释放大量炎症介质,同时上调组蛋白去乙酰化酶2(HDAC2)的表达。HDAC2的过度活跃会抑制BDNF基因的转录,从而导致BDNF蛋白合成减少。
简而言之,LPS就像一个信号,告诉大脑的免疫系统进入战备状态,而这种“战备状态”的代价之一,就是抑制了用于日常维护和修复大脑的“肥料”(BDNF)的生产。
——BDNF浓度下降与疲劳的关系
研究还发现:疲劳加剧的患者,血清中的脑源性神经营养因子(BDNF)浓度显著下降(P = 0.035)。BDNF被誉为“大脑的肥料”,是调节神经元能量代谢、促进神经可塑性和维持情绪稳定的核心物质。其水平的下降,直接与抑郁、认知障碍和疲劳感相关。这一发现也与以往“肠道菌群影响神经营养信号”的研究结论一致。
——肠道菌群失调对疲劳的影响
结合研究结果,为我们可以清晰理解肠道菌群通过肠道-脑轴影响癌因性疲乏的机制提供了线索。
本研究中观察到的血清脑源性神经营养因子BDNF水平下降,形成炎症与神经营养支持不足之间的恶性循环。很可能是短链脂肪酸减少与全身炎症增强共同作用的关键结果,而BNDF作为神经元能量代谢与可塑性的核心调控因子,其水平降低可能直接导致中枢性疲劳的发生(图4)。
图4:肠道菌群通过“肠-脑轴”影响癌因性疲劳的机制图
编辑
化疗导致菌群失调(产SCFA菌↓,革兰氏阴性菌↑) → 肠道屏障受损,LPS入血 → 激活TLR4等通路,引发全身炎症(IL-6↑) → 炎症信号和LPS穿过血脑屏障,抑制大脑BDNF表达(BDNF↓) → 最终导致中枢性疲劳(CRF↑)。
✍ 科普延伸:与“慢性疲劳综合征”异曲同工
其实,不仅是癌症化疗患者,在普通的肌痛性脑脊髓炎/慢性疲劳综合征(ME/CFS)患者中,科学家也观察到了极其相似的菌群特征。
一篇于2025年发表在《Journal of Translational Medicine》的综述指出,ME/CFS患者普遍存在肠道菌群失调,其核心特征同样是促炎类群(如变形菌门)增加,而产短链脂肪酸的有益菌(如粪杆菌、罗斯氏菌)减少。
这种跨疾病的共性,强有力地印证了本研究的结论:化疗作为一种强烈的外部冲击,诱导或加剧了与慢性疲劳疾病相似的肠道微生态失衡模式,最终通过“肠-脑轴”的神经炎症通路,在大脑中“投射”出了难以克服的疲劳感。
这项研究的重要价值在于,癌因性疲劳本是一种主观的、难以评估的症状,通过早期肠道菌群检测结合疲劳量表评估和相关血液标志物检测,可以为临床医生和患者提供了早期干预和防控的可能性。
总体而言,苏州大学的这项研究不仅为我们描绘了肠道菌群、神经免疫内分泌系统与化疗疲劳之间的复杂网络,更进一步证实并拓展了以往的研究结论:化疗前的肠道菌群特征,有望成为预测乳腺癌患者化疗期间CRF的生物标志物,例如,通过化疗前肠道菌群检测,评估其B/F比值、变形菌门丰度及产丁酸菌的比例,将可能成为由化疗引起的癌因性疲劳的重要评估预测指标。医生可以提前介入,给予更积极、更具针对性的支持性辅助干预。
通过监测血液标志物,来动态评估干预效果。血清BDNF水平的下降与疲劳加重密切相关。在临床实践中,血清BDNF可以作为一种便捷的血液生物标志物,用于动态追踪患者的疲劳进展,为干预措施的有效性提供有力证据。
我们可以从以下几个维度构建抗疲劳防线:
肠道菌群,作为连接化疗药物、宿主免疫和中枢神经系统的关键枢纽,未来有望成为肿瘤支持治疗的核心关注点。随着16S rDNA测序等高通量微生物组学技术的普及与成本降低,个体化肠道菌群检测或将成为肿瘤精准治疗不可或缺的一环。
注:本账号内容仅供学习和交流,不构成任何形式的医疗建议。

谷禾健康
我们谈到肠道菌群和植物化学物,总会说膳食纤维和多酚黄酮这些物质对身体好。但是你有没有想过,这些黄酮类化合物进入肠道,是谁在分解它们?是谁在利用它们?
今天我们要说的Flavonifractor(中文可以翻译为解黄酮菌属),就是一个专门能分解利用黄酮类化合物的肠道细菌。它这个名字,”Flavoni-“就是黄酮的意思,”-fractor”就是分解者,所以名字本身就告诉了你它的核心功能。
这个属是近年才被大家更多关注,它和很多疾病都有关系,包括痛风、糖尿病肾病、抑郁症等有牵连。今天我们就来全面认识它,帮助你拿到菌群检测报告后,能读懂结果,知道该怎么科学管理。
1
发现历史
Flavonifractor属其实很早已被分离,但近年才逐渐受到关注。随着对多酚黄酮与肠道菌群相互作用的研究深入,人们发现它在黄酮代谢中具有关键作用,因此相关研究不断增多。
其核心特点是能够裂解黄酮类化合物的C环,将大分子黄酮转化为更小的酚酸,这些小分子更易被机体吸收并发挥生物活性。因此,它在植物化学物代谢过程中具有重要意义。
2
分类定位
解黄酮菌属(Flavonifractor)在细菌分类学上的位置:
-门:厚壁菌门(Firmicutes)
-纲:梭菌纲(Clostridia)
-目:梭菌目(Clostridiales)
-科:毛螺菌科(Lachnospiraceae)
-属:解黄酮菌属(Flavonifractor)
这个属目前最主要的种,也是研究最多的,就是 Flavonifractor plautii。我们日常说肠道里的Flavonifractor,基本上就是指这个种。
3
核心特征
Flavonifractor有这些重要特点:
✔革兰氏染色:大部分该属细菌细胞壁为厚肽聚糖层,本质上是革兰阳性菌;但在临床标本和长期培养中常呈“革兰可变/假阴性”。
✔需氧性:只能在无氧环境生长,在有氧环境下会迅速失活,大肠下段环境适合它。
✔不产生芽孢:不会形成芽孢,对环境抵抗力一般。
✔细胞形态:直或轻度弯曲的杆菌,长度约2–10μm,单个或有时候成双或短链排列。
✔运动性:目前证据表明F.plautii不具鞭毛,为非运动性。
✔温度与pH:最适生长温度为37℃(与人体肠腔一致);生长pH范围约5.5–9.0,最适pH在6.0–7.5之间,符合结肠弱酸–中性环境。
✔黄酮代谢—产短链脂肪酸:其核心功能是能够代谢黄酮类化合物(名称由此而来),并具备产生短链脂肪酸(尤其是丁酸)及其他代谢物的能力。
4
在健康人肠道的丰度
Flavonifractor是高流行度、低丰度肠道共栖菌:在16S rRNA 或宏基因组数据集里,检出率很高(多数人有或可检出其属级信号),但在总菌群中通常只占0.1– 3%不等。
<来源:谷禾健康肠道菌群检测数据库>
-检出率可超过90%。
-相对丰度一般在0.1%—3%这个范围。(儿童粪便中的检出率和检出丰度会更高一些)
-饮食中多酚/黄酮类摄入高(如绿茶、蔬果丰富、某些亚洲饮食模式),丰度会高一点。
-长期低黄酮饮食,丰度会低一点。
-它属于毛螺菌科,毛螺菌科整体占肠道细菌10%-30%,Flavonifractor占其中一小部分。
5
核心:如何代谢黄酮类化合物
✔ 分解黄酮C环,这是它的绝活儿
黄酮类化合物是植物中的一大类多酚,结构中含有C环。多数细菌无法裂解该环,因而难以利用黄酮。
Flavonifractor的独特之处在于其能够催化黄酮C环的还原裂解,将大分子黄酮分解为更小的酚酸类产物。
这一过程至关重要,因为大分子黄酮不易被肠道吸收,而分解后生成的小分子酚酸更易进入血液,从而发挥抗氧化、抗炎等生物学作用。
因此,黄酮的吸收与功效在很大程度上依赖于Flavonifractor的作用。
✔ 主要代谢产物
Flavonifractor分解黄酮后,产生这些主要产物:
-对羟基苯丙酸;
-对羟基苯乙酸;
-其它小分子酚酸。
这些产物很多都有生物学活性,比如抗氧化、抗炎、抗菌。所以Flavonifractor本身不直接发挥作用,它通过活化植物黄酮发挥作用。
✔ 和其他细菌协作完成黄酮代谢
黄酮代谢不是Flavonifractor一个细菌就能完成的。它需要和其他细菌配合:
第一步:其他细菌先进行初步修饰,如去除糖苷键,释放苷元。
第二步:Flavonifractor随后还原裂解C环,生成小分子酚酸。
第三步:其他细菌进一步代谢这些酚酸,形成最终产物。
因此,它是黄酮代谢链中的关键环节;缺少它,代谢难以完成,黄酮也无法被充分活化。
✔ 它和哪些细菌一起生长?
研究发现,Flavonifractor丰度较高的人群通常也富含多酚利用菌,二者协同完成植物化学物的代谢过程。
此外,它还能与产丁酸菌共存,因为双方都偏好富含膳食纤维和植物化学物的环境。
6
尿酸代谢的特殊关系
近年研究发现,Flavonifractor plautii能分解嘌呤,产生尿酸。这个发现直接把它和痛风联系起来。
具体来说,它能利用嘌呤核苷,释放出游离尿酸。所以它丰度太高,会增加肠道尿酸产生,升高血尿酸水平,增加痛风风险。
这是近年一个非常重要的发现,直接把一个肠道细菌和一个常见病联系起来。
在正常丰度范围内存在的Flavonifractor,一般来说对人体健康是有益的,并能够发挥一些积极作用:
▸ 促进黄酮利用吸收
Flavonifractor可分解黄酮,生成可吸收的活性产物,从而发挥其抗氧化、抗炎作用,对黄酮功效至关重要。
膳食中的黄酮多为糖苷形式且分子较大,难以直接吸收;若Flavonifractor丰度不足,裂解不充分,黄酮难以完全活化,其效果也会降低。
▸ 参与代谢和维持肠道稳态
✔参与膳食纤维和植物化学物代谢:丰富的膳食纤维和多酚需要很多细菌协作分解,它是其中重要一员。
✔维持肠道生态多样性:它是正常菌群的一员,增加生态多样性,多样性是健康的基础。
✔能量回收:分解利用植物大分子,产生小分子能被宿主利用,回收一部分能量。
因此,正常丰度的Flavonifractor有助于维持肠道功能,并可能对部分疾病具有改善作用。
▸ 改善便秘
✔ Flavonifractor与便秘风险呈负相关
通过孟德尔随机化分析发现,Flavonifractor与便秘风险呈负相关关系,即其丰度较高时,可能有助于降低便秘发生的风险。
从机制上看,Flavonifractor作为肠道菌群的一部分,参与整体微生态平衡的维持。当其处于合适水平时,有助于优化肠道环境,从而间接促进肠道蠕动和排便功能的正常运行。
▸ 减轻肥胖
Flavonifractor是肠道健康的重要菌群,其含量与肥胖呈负相关。
口服Flavonifractor plautii可减轻肥胖脂肪组织的炎症反应,F.plautii可能参与抑制炎症环境中的TNF-α表达。
▸ 可能保护前列腺癌
PRACTICAL 和 FinnGen 联盟汇总的数据结果表明,Eubacterium fissicatena和Odoribacter与前列腺癌风险增加有关。相反,Adlercreutzia、Roseburia、Holdemania、Flavonifractor、Allisonella属则可能是预防前列腺癌的潜在保护因素。
▸ 减轻过敏与免疫反应
编辑
影响因子:5.085
研究发现,口服F.plautii可抑制小鼠的Th2免疫反应并降低IgE水平,同时促进调节性T细胞及特定树突状细胞的增加,这些变化共同提示其可能具有缓解过敏样反应的作用。
▸ 可能减轻急性肾损伤风险
编辑
影响因子: 3.37
一项研究及遗传因果推断结果提示,某一Flavonifractor亚型与急性肾损伤(AKI)风险的降低存在相关性,同时在临床队列研究中,该亚型还与更好的肾功能状态以及更低的死亡率呈相关关系。
▸ 缓解结肠炎
个别研究报道,口服 Flavonifractor plautii可通过抑制 IL-17 信号传导促进小鼠急性结肠炎的恢复。
虽然正常丰度的Flavonifractor对人体有益,但当其丰度过高或过低时,可能对健康产生不良影响。
下面分别来看其升高和降低与哪些疾病相关。
Part 1
丰度升高,和这些疾病相关
1
提升痛风概率
这是目前证据最强的关联。
研究发现痛风患者肠道中Flavonifractor plautii丰度显著升高。它能分解嘌呤核苷产生尿酸,肠道尿酸吸收增加,导致血尿酸升高。
研究还发现,降低它的丰度能减少血尿酸产生,帮助降尿酸。这个发现打开了痛风治疗新方向,就是通过调节肠道菌群降尿酸。
编辑
2025年另一项研究进一步验证,在无症状高尿酸血症患者,也看到Flavonifractor丰度升高。所以这个关联比较确定。
✔ 机制深入解读:Flavonifractor怎么影响尿酸
很多人会问,为什么肠道细菌会影响尿酸?
我们每天吃进去的食物,含有很多嘌呤核苷。这些核苷消化吸收不完全,到达结肠。Flavonifractor能分解这些嘌呤核苷,把嘌呤释放出来,嘌呤进一步分解产生尿酸。尿酸在结肠被吸收进入血液,增加整体血尿酸水平。
编辑
如果Flavonifractor丰度高,分解产生更多尿酸,血尿酸就容易升高。如果你本来就有痛风,这个影响就更明显。
这个机制是2023年科学家明确发现的,现在已经被多项研究重复证实。所以痛风患者调节Flavonifractor丰度,是有科学依据的辅助治疗方法。
2
结直肠癌患者中丰度升高
编辑
影响因子:6.633
多项研究发现,结直肠癌患者肠道中Flavonifractor丰度显著升高。
✔ 为什么它和结直肠癌相关?可能的机制:
•首先,F.plautii是一种能够降解黄酮类化合物的肠道细菌,而黄酮类物质通常具有抗炎、抗氧化及潜在抗肿瘤作用。其对黄酮的降解可能削弱这些保护性作用,从而为肿瘤发生创造有利环境。
•其次,该菌在结直肠癌患者肠道中呈现相关性变化,提示其参与肠道微生态失衡,而这种菌群失衡已被认为是结直肠癌的重要影响因素之一。
编辑
目前机制还在研究,但是关联已经被多项研究证实。
3
慢性肾病、尿结石
由于Flavonifractor会增加尿酸的水平,所以研究人员认为其丰度过高与慢性肾病及泌尿道结石存在关联,且研究证实了这一点。
✔ 慢性肾病个体中丰度更高
根特大学医院招募的 110 名非慢性肾病和慢性肾病患者,发现Flavonifractor属在慢性肾病个体中的水平高于非慢性肾病个体。
✔ 尿路结石风险
泌尿道结石可引起一系列并发症,如尿路阻塞、感染、不适以及对肾脏的潜在不可逆损害。
编辑
一项双向双样本孟德尔随机化研究显示,Flavonifractor属丰度增加(IVW OR = 0.69,95%CI 0.53-0.91,P = 8.57 × 10-3)与尿路结石形成风险之间存在因果关系。
4
双向情感障碍中丰度增加
编辑
影响因子:6.633
在新诊断双相情感障碍(BD)患者中,Flavonifractor的检出率显著高于健康对照(约65% vs 39%),提示其与疾病存在关联,且BD患者更可能携带该菌。
5
抑郁症中丰度升高
编辑
多项研究表明,在抑郁症人群中Flavonifractor丰度或检出率升高。常伴随抗炎有益菌(如Faecalibacterium)减少,提示菌群结构向不利状态转变。
✔ 可能机制:
•促进炎症反应:Flavonifractor增加与促炎菌群特征相关,可能通过增强低度全身炎症,参与抑郁症发生发展。
•削弱抗炎保护:其升高常伴随抗炎菌减少,打破肠道免疫平衡,使炎症信号更易通过“肠-脑轴”影响中枢神经系统。
•多酚/黄酮代谢:Flavonifractor可降解具有抗氧化和神经保护作用的黄酮类物质,可能削弱抗氧化能力,促进氧化应激,而氧化应激与抑郁密切相关。
•肠-脑轴调控:通过影响肠屏障功能、代谢产物及免疫信号,间接调节神经递质系统(如5-HT)和脑功能,从而影响情绪和认知。
6
糖尿病肾病中增加
编辑
一项研究发现,2型糖尿病患者Flavonifractor丰度显著高于健康对照。它丰度升高和胰岛素敏感性下降、糖调节异常增加相关。
✔ 糖尿病肾病患者中Flavonifractor也增加
编辑
影响因子:3.998
此外,糖尿病相关肾病患者中也发现Flavonifractor水平进一步改变。其可能机制包括:
•代谢通路调节:Flavonifractor可降解黄酮/多酚类物质,过度代谢可能削弱其抗氧化、抗炎作用,加重氧化应激与代谢紊乱,从而促进肾损伤。
•炎症与毒素相关通路:糖尿病肾病(DKD)人群中伴随细菌毒素相关功能增强,提示包括Flavonifractor在内的菌群变化可能促进低度炎症和内毒素负荷。
•肠-肾轴作用:菌群失衡可能影响肠屏障功能,增加有害代谢产物入血,进而损伤肾脏。
Part 2
丰度降低,和这些疾病相关
1
自闭症患者中丰度降低
编辑
影响因子: 3.727
在该研究中发现,Flavonifractor在自闭症谱系障碍(ASD)组中的平均相对频率为0.07%,而在对照组中为0.68%,表明丰度在ASD儿童中显著减少。
文献进一步指出,物种Flavonifractor plautii在ASD组中的丰度为0.05%,在对照组中为0.61%,同样显著降低(p < 0.05)。物种水平比较共识别出11个丰度差异显著的物种,Flavonifractor plautii是其中之一。
✔ 可能机制:
•代谢途径扰动:通过PICRUSt功能预测,ASD儿童的肠道微生物群显示多条代谢途径显著减少,包括赖氨酸降解和色氨酸代谢。Flavonifractor可能参与短链脂肪酸(SCFAs)或其他代谢物的生产,其减少可能导致SCFAs水平变化,进而影响肠道屏障功能和神经炎症。
•免疫和炎症关联:功能预测中,ASD儿童唯一富集的模块是“细胞抗原”,提示免疫应答异常。Flavonifractor的减少可能削弱微生物群对肠道黏膜的维护作用,加剧免疫失调,这与文献中提到的ASD儿童常伴随胃肠道症状(如肠漏症)一致。
编辑
•与其他菌属的相互作用:Flavonifractor的减少与多个有益菌(如Akkermansia muciniphila)的降低同步出现,可能共同导致微生物群生态失衡。
注:ASD儿童肠道微生物群的变化具有个体差异性,这可能意味着Flavonifractor的减少在部分儿童中更为显著,进而通过肠脑轴影响ASD严重程度。
2
心血管疾病患者中减少
一些研究发现,心血管疾病患者Flavonifractor丰度降低。因为心血管疾病患者常常黄酮摄入不足,所以它丰度低。
黄酮活化减少,抗氧化抗炎作用减弱,可能加重心血管疾病进展。
✔ Flavonifractor plautii 改善动脉硬化程度
人类粪便宏基因组测序显示,在正常对照组中,F.plautii丰度显著较高,并在微生物群落中处于中心地位,而在动脉僵硬度升高的受试者中,F. plautii缺失。此外,血压只部分介导了F.plautii对降低动脉僵硬度的影响。
正常对照组的微生物组表现出增强的糖酵解和多糖降解能力,而动脉僵硬度增加的受试者的微生物组则以脂肪酸和芳香族氨基酸的生物合成增加为特征。
整合代谢组学分析进一步表明,顺式乌头酸的增加是F. plautii对动脉硬化保护作用的主要效应物,通过抑制基质金属蛋白酶-2的激活,维持弹性纤维网络,缓解动脉功能障碍。
编辑
doi.org/10.1161/CIRCRESAHA.122.321975
我们一起来看看,在生活的哪些常见因素,可能会使Flavonifractor的水平出现升高或降低的变化情况。
1
饮食因素(最主要)
✔ 黄酮类化合物摄入增加其丰度
这是最主要的影响因素。你吃进去的黄酮越多,Flavonifractor底物越多,它丰度越高。所以长期吃蔬菜水果少的人,它丰度会降低。
富含黄酮的食物包括有色蔬果、红茶、绿茶、咖啡、黑巧克力和浆果等。此外,番石榴、榄仁树皮、葫芦巴籽、芥菜籽、肉桂、红辣椒粉、丁香、姜黄及豆类也都是类黄酮的良好来源。
✔ 高嘌呤饮食增加其丰度
Flavonifractor能分解嘌呤核苷产生尿酸。你吃进去的嘌呤越多,底物越多,它丰度越高。所以高嘌呤饮食会促进它生长。
✔ 膳食纤维摄入
膳食纤维丰富的饮食,一般黄酮也丰富,所以膳食纤维够能维持它正常丰度。膳食纤维太少,它丰度容易降低。
✔ 大量酒精摄入会降低其丰度
酒精会改变整体肠道菌群的结构,而其对Flavonifractor的具体影响取决于摄入剂量,一般来说,长期且大量饮酒通常会使其丰度出现下降的趋势。
2
生活方式
✔ 年龄大的丰度反而高
研究发现,年龄越大,Flavonifractor丰度越高。老年人丰度一般比年轻人高。
✔ 运动有助于维持正常平衡
规律运动改善整体菌群多样性,对Flavonifractor影响要看其他因素,运动一般帮助维持正常平衡。
3
疾病和药物因素
✔ 炎症性肠病中Flavonifractor减少
在活动期炎症性肠病状态下,由于整体肠道菌群发生失调,Flavonifractor的丰度通常会出现明显降低的情况。
✔ 抗生素影响其丰度
抗生素使用会改变菌群结构,对Flavonifractor影响要看抗生素种类,广谱抗生素一般会降低它的丰度。
现在当你拿到一份菌群检测报告,看到其中关于Flavonifractor的检测结果时,应该如何进行判断以及后续该如何处理,我们一步一步来详细说明。
1
判断丰度是否正常
健康人Flavonifractor正常参考范围:
•0.1% – 0.415%:正常范围,不用特殊干预;
•>0.415%:丰度过高(注:儿童中是>2.778%才为过高)
•<0.1%:丰度过低。
这个范围是基于人群研究总结,具体请以你的检测报告给出的参考范围为准。
2
结合你的健康状况,判断意义
在不同健康状况下,Flavonifractor丰度异常的意义各不相同,主要情况如下:
✔ 痛风或高尿酸血症:Flavonifractor丰度升高,需要进行干预,降低其水平有助于控制尿酸。
✔ 结直肠癌:Flavonifractor丰度升高,需要重点关注,通过调整饮食并定期复查进行管理。
✔ 糖尿病肾病:Flavonifractor丰度升高,需要干预,以帮助改善胰岛素抵抗。
✔ 抑郁症:Flavonifractor丰度升高,需要干预,通过调节神经递质水平改善情绪。
✔ 炎症性肠病:Flavonifractor丰度降低,需要从整体上调节肠道菌群,以恢复其正常水平。
✔ 健康人:Flavonifractor处于正常范围,无需干预,保持健康的生活方式即可。
3
针对性干预
如果你的Flavonifractor丰度升高需要降低,该如何应对?这种情况常见于痛风或高尿酸血症、结直肠癌及糖尿病肾病。
✔ 饮食调整(核心)
–限制嘌呤摄入:减少高嘌呤食物摄入,以降低底物供给。高嘌呤食物比如动物内脏、海鲜、红肉,适当减少。每天嘌呤摄入控制在150毫克以内,这个是痛风患者的标准。
-虽然黄酮对一般人有益,但若痛风且Flavonifractor偏高是否需要限制?不必。黄酮本身具有抗炎作用,对痛风有益,其影响远小于嘌呤,蔬菜水果仍可正常食用,无需担心。
-增加膳食纤维:保证膳食纤维摄入,维持菌群平衡。
-多喝水:帮助尿酸排泄,这对痛风也有好处。
✔ 生活方式
-规律运动:帮助维持菌群平衡,帮助控制体重,体重正常对痛风也有好处。
-戒烟限酒:酒精影响尿酸排泄,也改变菌群,所以要限酒。
✔ 医学干预
-如果您确诊痛风,一定要遵医嘱吃药降尿酸,饮食调节和菌群调节是辅助,不能代替药物。
-目前没有特异性抗生素降Flavonifractor,也不需要用抗生素,饮食调整就够了。
如果你的Flavonifractor丰度降低需要提升,应如何应对?这种情况常见于炎症性肠病和心血管疾病。
✔ 饮食调整(核心)
-增加富含黄酮的食物摄入:这是最直接的,给它提供底物,促进它生长。
-每天保证吃够蔬菜水果:特别是有色蔬菜水果,比如蓝莓、草莓、葡萄、紫甘蓝、胡萝卜这些,颜色越深黄酮越多。
-可以适量喝茶:红茶、绿茶,这些茶叶黄酮含量很高。
-适量吃黑巧克力(70%以上可可),黄酮含量也很高。
✔ 生活方式
-规律吃蔬菜水果:养成每天吃蔬菜水果的习惯,持续给它提供底物,慢慢丰度就上来了。
-减少抗生素不必要使用:抗生素会杀它,没事不要乱吃抗生素。
✔ 医学干预
-心血管疾病等:一定要遵医嘱规范治疗,饮食调节菌群是辅助。
-炎症性肠病:规范治疗基础疾病,炎症控制好了,菌群慢慢恢复,丰度也会恢复。
我们在这帮你整理出了一份清晰明了的清单,你可以直接参考并使用:
Flavonifractor过高,推荐这样吃
✔ 推荐多吃
主食——低嘌呤全谷物:大米、白面、燕麦(嘌呤低,可以提供膳食纤维);
蔬菜——浅色蔬菜为主,深色蔬菜适量(深色蔬菜黄酮嘌呤都高,适量吃没问题,不用完全不吃);
蛋白质——鸡蛋、牛奶、鸡肉、淡水鱼(嘌呤低,满足蛋白质需要);
水果——低果糖水果:苹果、梨、桃子(提供维生素和纤维,嘌呤不高)。
✔ 需要控制少吃
高嘌呤食物——动物内脏、海鲜、红肉、肉汤(嘌呤高,给Flavonifractor提供太多底物);
酒精——啤酒、白酒(升高尿酸,改变菌群);
高糖饮料——含糖可乐、果汁(不利于代谢,改变菌群)。
Flavonifractor过低,推荐这样吃
✔ 多吃
蓝莓、草莓、树莓、紫葡萄:黄酮含量非常高;
紫甘蓝、茄子、胡萝卜、菠菜:黄酮含量高;
苹果、梨、樱桃:黄酮含量中高;
茶——红茶、绿茶:黄酮含量很高;
黑巧克力(70%+可可):黄酮含量高;
咖啡:黄酮含量中等 ;
全谷物——燕麦、藜麦、全麦:黄酮含量中等,同时提供纤维。
✔ 少吃
过度加工食品:缺乏黄酮和纤维;
酒精:改变菌群,降低丰度。
它不是好细菌也不是坏细菌,它是正常肠道细菌。正常丰度对健康有益,能帮助活化黄酮。
丰度太高增加痛风和糖尿病风险,太低和抑郁炎症性肠病相关。所以关键是正常范围,不是它本身好坏。
不用。饮食减少嘌呤摄入,减少底物,它丰度慢慢自然降下来。不需要吃药杀它。饮食调整安全有效,是首选。
多数人能。只要持续每天摄入足够黄酮,给它提供底物,它慢慢就能长上来。这个过程大概需要一两个月。
不用怕。黄酮带来的好处远远大于底物增加这点影响。而且黄酮本身有抗炎作用,对痛风反而有好处。一般限制嘌呤就够了,蔬菜水果还是要吃。
不一定。它是风险因素,不是说高了一定得痛风。痛风发病是多因素,遗传、排泄、饮食都有关系。它高了只是增加风险,提示注意控制嘌呤,定期查血尿酸。
目前没有商品化的Flavonifractor益生菌。一般通过饮食调整就能把丰度调到正常,并且饮食调整更安全。
饮食调整后,一般两到三个月复查一次。调整正常了,以后一年复查一次就够了。
主要参考文献
Kvernberg J, Rehn M, Johansen IS, Peterslund P, Hansen PM. Flavonifractor plautii: A rare pathogen in sepsis and critical care—A systematic narrative review of literature. Am J Infect Control. 2026 Jan 29:S0196-6553(26)00050-7.
Zou R, Xu F, Wang Y, Duan M, Guo M, Zhang Q, Zhao H, Zheng H. Changes in the Gut Microbiota of Children with Autism Spectrum Disorder. Autism Res. 2020 Sep;13(9):1614-1625.
Coello K, Hansen TH, Sørensen N, Munkholm K, Kessing LV, Pedersen O, Vinberg M. Gut microbiota composition in patients with newly diagnosed bipolar disorder and their unaffected first-degree relatives. Brain Behav Immun. 2019 Jan;75:112-118.
Ogita T, Yamamoto Y, Mikami A, Shigemori S, Sato T, Shimosato T. Oral Administration of Flavonifractor plautii Strongly Suppresses Th2 Immune Responses in Mice. Front Immunol. 2020 Feb 28;11:379.
Wu M, Zhou X, Chen S, Wang Y, Lu B, Zhang A, Zhu Y, Huang M, Wang J, Liu J, Zhu F, Liu H, Lin R. The alternations of gut microbiota in diabetic kidney disease: insights from a triple comparative cohort. Front Cell Infect Microbiol. 2025 Jun 27;15:1606700.
Lu X, Ma J, Li R. Alterations of gut microbiota in biopsy-proven diabetic nephropathy and a long history of diabetes without kidney damage. Sci Rep. 2023 Jul 27;13(1):12150.
Gupta A, Dhakan DB, Maji A, Saxena R, P K VP, Mahajan S, Pulikkan J, Kurian J, Gomez AM, Scaria J, Amato KR, Sharma AK, Sharma VK. Association of Flavonifractor plautii, a Flavonoid-Degrading Bacterium, with the Gut Microbiome of Colorectal Cancer Patients in India. mSystems. 2019 Nov 12;4(6):e00438-19.
Pan Y, Su J, Liu S, Li Y, Xu G. Causal effects of gut microbiota on the risk of urinary tract stones: A bidirectional two-sample mendelian randomization study. Heliyon. 2024 Feb 14;10(4):e25704.