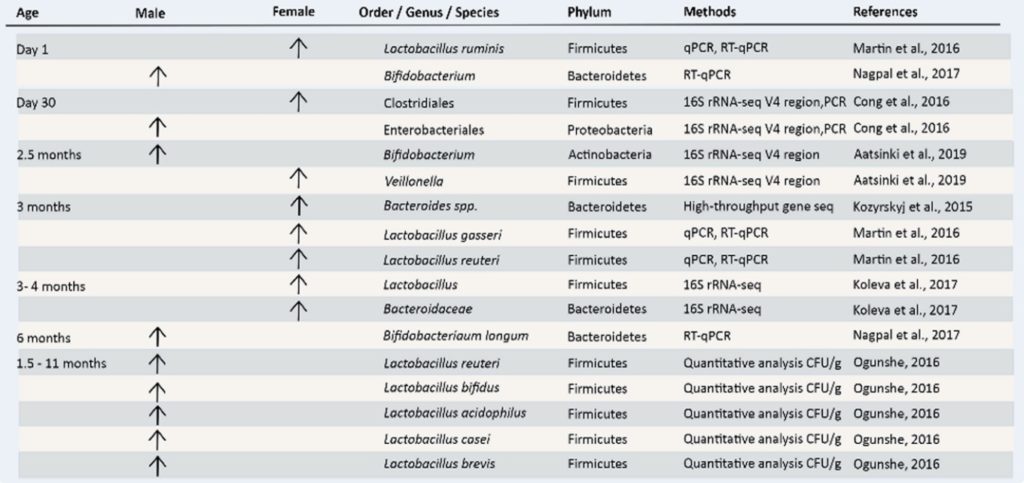
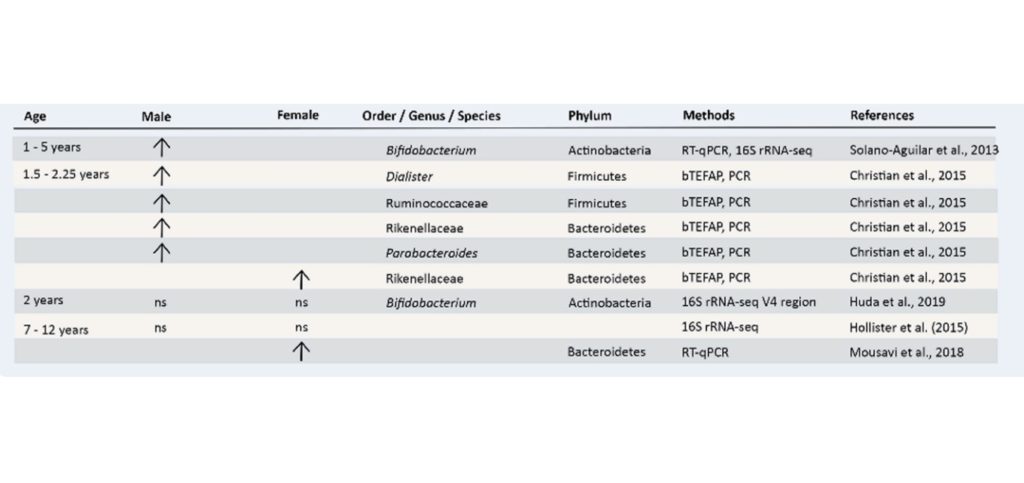
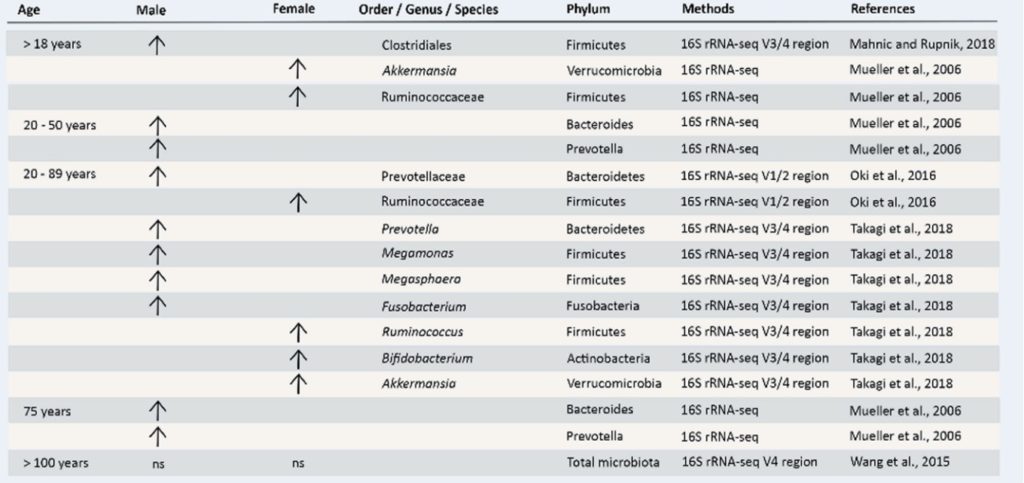
 国家高新企业 | ISO9001认证
国家高新企业 | ISO9001认证 二级病原微生物安全实验室
二级病原微生物安全实验室- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
国家高新企业 | ISO9001认证 二级病原微生物安全实验室

谷禾健康
花青素-肠道微生物群-健康轴
经常听到一些专家在介绍食品或保健品时说富含花青素,那么究竟什么是花青素?对我们的健康又有何益处?
花青素是在植物组织中发现的天然水溶性色素,水果、蔬菜、花卉中的呈色物质大部分与之有关,是类黄酮的一个亚类,迄今为止已鉴定出700多种结构不同的花青素。
花青素是一种重要的营养素,具有多种生物活性和保健作用,如抗氧化、抗炎、抗癌、降血压、降血糖、保护视力及心血管健康。
花青素在消化过程中还会影响肠道中的细菌代谢。花青素及其衍生物还通过调节肠道屏障功能和有益细菌的定植来促进肠道健康,从而更好地抵御病原体,改善营养代谢和整体免疫反应。
在本文中,谷禾简要介绍了不同类型的花青素,它们常见的饮食来源和通过胃肠道(包括胃、小肠和结肠)的吸收和代谢。
本文还总结了膳食花青素对健康的直接影响,以及肠道微生物群作为“媒介”时的影响。花青素-微生物的相互作用可能成为肥胖、2型糖尿病、心血管疾病、神经退行性疾病、炎症性肠病、癌症、慢性肾脏病和骨关节炎等疾病的治疗新途径。
目录
1.常见的花青素及膳食来源
2.膳食花青素在人体内的吸收和代谢
3.花青素对肠道微生物群的影响
4.花青素-微生物群对肥胖的改善作用
5.花青素-肠道菌群对糖尿病的影响
6.花青素改善心血管疾病
7.花青素降低神经退行性疾病风险
8.花青素对其他疾病的影响
9.日常补充膳食花青素的一些建议
花青素又称花色素,是自然界一类广泛存在于植物中的水溶性天然色素,是花色苷水解而得的有颜色的苷元。
水果、蔬菜、花卉中的主要呈色物质大部分与之有关。在植物细胞液泡不同的PH值条件下,花青素使花瓣呈现五彩缤纷的颜色。
▼
已知现有的花青素多达几百种,食物中重要的有6种,即天竺葵色素、矢车菊色素、飞燕草色素、芍药色素、牵牛花色素和锦葵色素。
花青素具有2-苯基苯并吡喃结构,具有完全不饱和的C环和3位的羟基。
植物性食物中主要花青素的结构
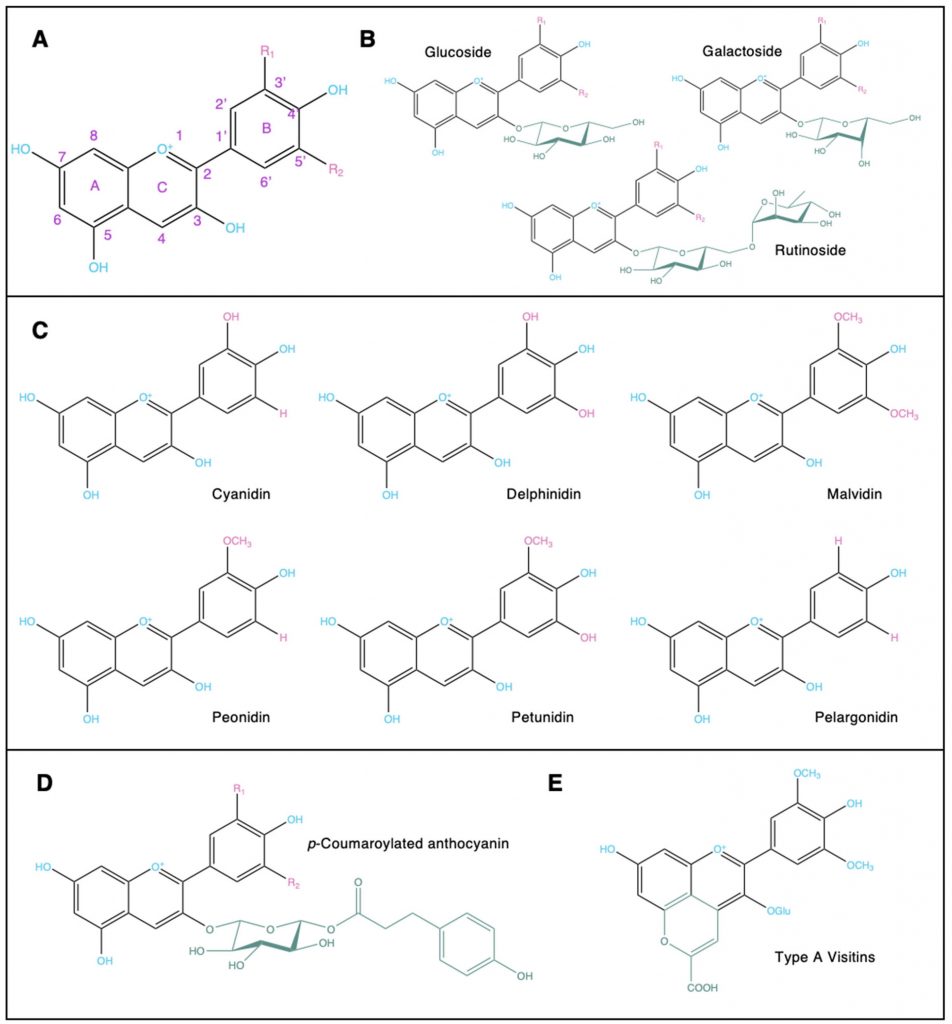

Liang A,et al.Crit Rev Food Sci Nutr.2023
(A)花青素中的2-苯基苯并吡喃骨架 (B)附着在糖基化花青素上的常见糖部分 (C)植物性食物中常见的花青素 (D)酰化花青素 (E)吡喃花青素
▼
植物性食品中花青素的浓度和组成高度依赖于植物品种、生长区域、植物部位、加工方法和测量方法。
花青素在各种红色、紫色和蓝色水果(如浆果、石榴、血橙和红梅)中含量丰富,这些水果代表了人类饮食中花青素的主要摄入。
常见水果中的花青素
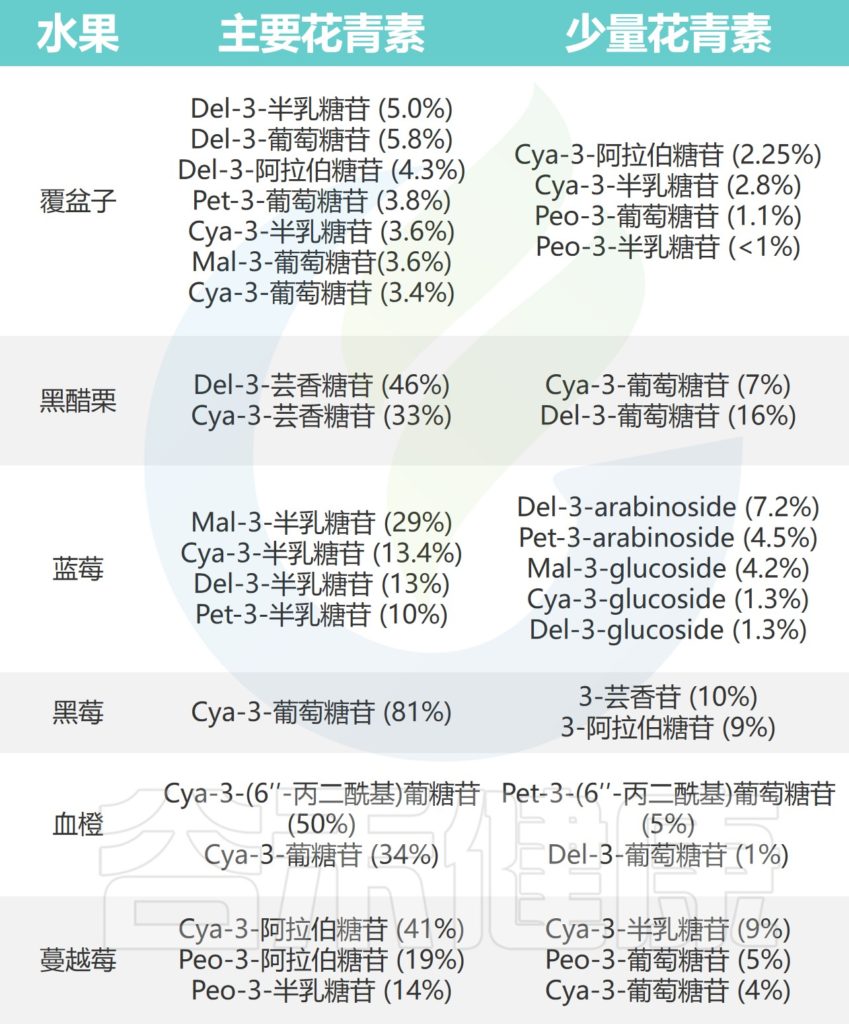

Liang A,et al.Crit Rev Food Sci Nutr.2023
•花青素在不同酸碱度下颜色不同
花青素在水果中根据pH条件,它会显示红色(pH<3)、紫色(pH=7-8)及蓝色(pH>11)。
例如飞燕草色素通常存在于花朵和葡萄中,它们提供蓝紫色,而天竺葵色素为许多浆果和水果提供橙色和红色,包括树莓、草莓和石榴。
•酰化花青素具有更强的耐热性和耐光性
酰化花青素是通过糖基和有机酸的乙酰化酯键得到的,进一步扩大了食品中花青素的多样性。常见的酰化酸包括脂肪酸(乙酸、丙二酸、苹果酸、草酸和琥珀酸)和环状酸(对香豆酸、咖啡酸、芥子酸、没食子酸和阿魏酸)。
在葡萄中鉴定出酰化花青素,在黑胡萝卜、红薯和红甘蓝中发现了肉桂酸酰化的花青素。在红萝卜中发现了多达12种酰化花青素。
酰化花青素在加工过程中表现出更强的耐热性和耐光性,因此被广泛用作食品工业中的着色剂。
•吡喃花青素具有更强的抗氧化能力
吡喃花青素是另一组具有额外吡喃环的花青素,主要在红酒的酒精发酵和陈酿过程中产生。少量吡喃花青素天然存在于血橙、草莓和黑加仑等水果中。
吡喃花青素的主要类别包括A型和B型视黄素、甲基吡喃花青素、羟基苯基吡喃花青素和黄烷基吡喃花青素。
相较于单体花青素,吡喃花青素,在更大范围的pH条件下保持其颜色,这是因为额外的吡喃环可以防止亲核攻击和无色甲醇碱的形成。
吡喃花青素在二氧化硫漂白和热降解过程中也具有更大的稳定性,并且相对于单体花青素具有更强的抗氧化能力。
由于这些原因,吡喃花青素作为颜料和生物活性剂广泛用于食品加工。
膳食花青素是指可以通过饮食摄入的一类花青素,广泛存在于深红色、紫色或蓝色蔬菜水果中,如葡萄、蓝莓、紫甘蓝、紫薯、黑枸杞等。
膳食花青素的代谢和治疗活性取决于其生物可及性(可从食物基质中获得的量)和生物利用度(在人体肠道中的吸收)。这受到许多因素的影响,包括化学结构、与食物基质的相互作用、其他膳食化合物的存在以及个体遗传学和生理学差异。
★ 不同结构的花青素消化率存在差异
例如,具有双糖部分的糖基化花青素相对于单葡萄糖苷和苷元更耐消化。酰化花青素的消化率低于非酰化化合物,主要是因为极性降低和酰基的阻碍。吡喃花青素和聚合物花青素在刺激消化过程中更稳定。
花青素及其衍生物在人体内的消化吸收发生在胃、小肠和大肠中。
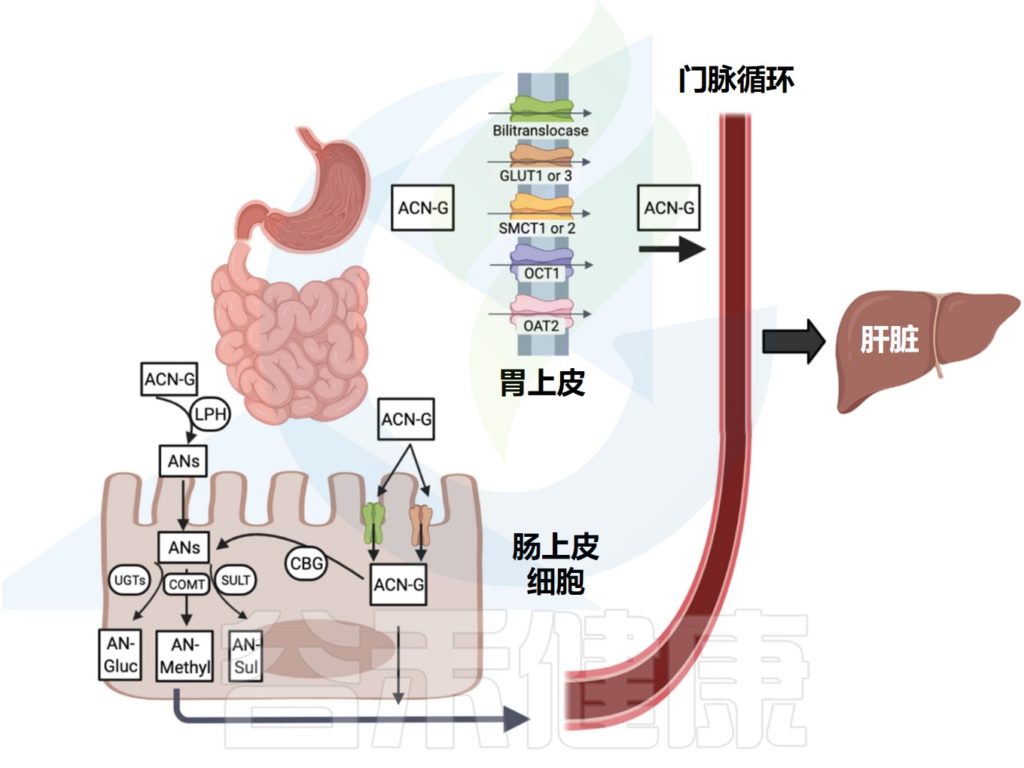
Liang A,et al.Crit Rev Food Sci Nutr.2023
▼
食用食物后第一个吸收花青素的部位是胃,在胃中,一小部分花青素穿过胃细胞屏障(大部分以完整的糖基化形式),并在血浆中检测到。
在胃内,花青素保持相对稳定,在大鼠胃肠道研究期间,从流出物中可回收总摄入花青素的75-79%,在体外研究期间可回收75-88%。
✦胃吸收效率受膳食成分影响
胃吸收效率也受到膳食成分的影响,在高葡萄糖 (>40mM)存在的情况下,花青素通过胃细胞屏障的摄取速率显著降低。
✦参与胃中花青素代谢的蛋白
胃胆转位酶是一种膜蛋白载体,可将完整的花青素转运穿过胃屏障。花青素的糖苷部分优先满足双转位酶提供的结构规格,因此无需去糖基化即可被吸收。
后来,葡萄糖转运蛋白GLUT1和GLUT3也参与了胃对花青素的摄取。花青素中的A环和C环以及葡萄糖部分在与人类GLUT1和GLUT3载体的跨膜结合中起重要作用。
其他参与花青素在胃中吸收的转运蛋白包括单羧酸转运蛋白(SMCT1和SMCT2)和有机阳离子/阴离子转运蛋白(OCT1和OAT2)。
胃以前被认为是花青素吸收而非代谢的部位,直到最近在胃上皮细胞中发现结合酶活性才证实了胃的代谢功能。
此外,红酒中花青素在体外消化(使用胃MKN-28和肠道Caco-2细胞)过程中代谢物malvidin-3-O-葡糖苷酸的存在提供了花青素在胃内代谢的证据。
▼
空肠(小肠中部)是花青素吸收的主要部位。花青素在小肠内的吸收通过膜转运蛋白促进的被动扩散或主动转运发生。
✦花青素的主动转运吸收
与胃吸收类似,完整的糖基化花青素被肠上皮细胞内的转位酶和葡萄糖转运蛋白(SGLT1和GLUT2)吸收。
SGLT1转运蛋白是一种在肠细胞顶端表面发现的钠-葡萄糖共转运蛋白1,而GLUT2转运蛋白是主要在上皮细胞基底外侧膜上发现的促进载体。这两种转运蛋白都与花青素的葡萄糖部分相互作用,并负责基于葡萄糖的花青素的肠道吸收。
除了己糖转运(糖、葡萄糖、果糖等简单碳水化合物的转运系统)外,最近发现有机阴离子转运多肽转运体(OATP)也参与了大鼠空肠上皮对完整β型酰化花青素的吸收,但其吸收率很低,仅为0.2%~2.2%。
✦花青素的被动扩散吸收
另一方面,当花青素被乳糖酶-根皮苷水解酶、β-葡萄糖苷酶、α-鼠李糖苷酶和β-葡萄糖醛酸酶等水解为其苷元形式时,花青素通过肠上皮的被动扩散吸收。
✦花青素的转运速率
最近使用Caco-2细胞(人类结直肠腺癌细胞)作为人类肠道屏障功能的模型,可以评估花青素的运输,并表明运输效率取决于附着在花青素骨架上的苷元或糖的类型,以及花青素是否为聚合物。
Caco-2细胞模型还估计,达到系统循环的花青素比例约为总膳食摄入量的1%。
✦肠道内花青素的代谢
在肠道内吸收的花青素经历第一阶段代谢,包括氧化、还原和水解,但它们发生的频率较低,在多酚代谢中也不太重要。
第二阶段的生物转化更加密集和快速,它们首先发生在小肠,然后发生在肝脏和肾脏。一旦被肠细胞吸收,花青素及其分解代谢物就会发生结合反应。
包括分别由UDP葡萄糖醛酸基转移酶(UGTs)、硫转移酶(SULT)和邻苯二酚-O甲基转移酶(COMT)催化的葡萄糖醛酸化、硫酸化和甲基化。
这些生物转化产生葡萄糖醛酸、硫酸盐和甲基化代谢产物,这些代谢产物由活性ATP结合盒蛋白转运,因为它们过于亲水,无法被动扩散到细胞中。
活性ATP结合盒蛋白——一个膜内在蛋白质超家族。将ATP水解释出的能量提供给各种分子进行穿膜转运。
花青素进入血液后,通过门静脉分布到肝组织,进行进一步的偶联反应,增加其在水中的溶解度,并最终促进其在尿液中的排泄。
▼
膳食中花青素的低生物利用度以及粪便和尿液样本中花青素的低回收率表明,花青素在结肠中会被降解。
例如,食用覆盆子后,总膳食花青素的40%从回肠液中回收。这些来自小肠的未被吸收的花青素通过细菌或化学降解进行生物转化,潜在地提高了它们的生物利用度,并允许通过结肠上皮壁吸收。
花青素的微生物降解需要多种细菌,如拟杆菌、Enterococcus casseliflavus、优/真杆菌属(Eubacterium)和梭状芽胞杆菌(Clostridium)等,它们产生各种酶,如β-葡萄糖苷酶、α-半乳糖苷酶和α-鼠李糖苷酶,用于微生物分解代谢。
✦原儿茶酸是最丰富的代谢物
由于这些相互作用,通过肠道微生物降解产生了各种代谢产物。代谢物原儿茶酸占花青素摄入量的73%,是花青素消耗后最丰富的人体代谢物。
桑椹的矢车菊素-3-O-葡萄糖苷和矢车菊苷-3-O-芸香糖苷的体外粪便发酵产生了原儿茶酸、对香豆酸和间苯三酚醛,而飞燕草素-3-O-芸香糖苷的发酵产生了没食子酸、丁香酸和间苯三酚醛。
✦其他花青素的代谢产物
桑椹的矢车菊素-3-O-葡萄糖苷和矢车菊苷-3-O-芸香糖苷的体外粪便发酵产生了原儿茶酸、对香豆酸和间苯三酚醛,而飞燕草素-3-O-芸香糖苷的发酵产生了没食子酸、丁香酸和间苯三酚醛。
在与结肠细菌孵育24小时后,Malvidin-3-O-葡萄糖苷会完全代谢为丁香酸。
如体内人体研究和体外研究所示,天竺葵素-3-O-葡糖苷被代谢为4-羟基苯甲酸。
六种主要花青素及其结肠代谢物


Liang A,et al.Crit Rev Food Sci Nutr.2023
★ 花青素的主要代谢物对人体健康有益
原儿茶酸
原儿茶酸是一种水溶性酚酸成分,并且是很多中药中的活性物质,其不仅具有抗血小板凝集、降低心肌耗氧量、提高心肌耐氧能力、减慢心率、抑菌、镇痛等药理活性,还具有抗氧化、抗肿瘤和神经保护作用。
没食子酸
没食子酸具有抗炎、抗突变、抗氧化、抗自由基等多种生物学活性;同时没食子酸具有抗肿瘤作用,可以抑制肥大细胞瘤的转移, 从而延长生存期;对肝脏具有保护作用。
丁香酸
丁香酸可以抑制炎症反应,减轻炎症症状;同时丁香酸具有抗菌作用,可以抑制多种细菌和真菌的生长;丁香酸还能够降血糖、降血脂。
✦肠道微生物在花青素的去酰化起重要作用
此外,人体肠道微生物组在花青素的去酰化过程中发挥重要作用,释放出具有更高生物利用度的酚酸。
花青素衍生的代谢物可能比母体花青素表现出更强的稳定性和生物活性,并可能调节人体肠道微生物组成。
肠道微生物群受年龄、体重、饮食模式、抗生素治疗和宿主遗传的影响,并且个体之间(以及个体内部)存在差异。这种变化可能会影响膳食花青素的生物利用度和代谢。
花青素及其结肠代谢物通过其生物活性直接改善健康,并通过重塑肠道微生物组的组成和操纵短链脂肪酸的产生间接改善健康。
在宿主的一生中,肠道微生物组的组成随着时间的推移而变化,以响应内部和外部刺激。因此,肠道微生物组成的动态平衡和多样性是正常肠道功能所必需的,包括维生素和短链脂肪酸的产生、脂质代谢、免疫防御和肠-脑轴相互作用等。
▼
花青素及其代谢物目前被认为是改变肠道微生物群的调节剂,主要是通过促进有益菌的增殖和抑制有害菌的生长,许多不同模型的研究都观察到这一点。
✦促进了有益菌的增殖
研究发现黑米中分离的矢车菊素-3-葡萄糖苷在体外发酵条件下对青春双歧杆菌(B. adolescentis)、婴儿双歧杆菌(B. infantis)、双歧双歧杆菌(B.bifidum)和嗜酸乳酸杆菌(L.acidophilus)的增殖具有促进作用。
培养48小时后还观察到生长培养基pH值的降低,这归因于由酚酸和短链脂肪酸组成的有机酸的产生。双歧杆菌和乳杆菌的数量增加。它们通过竞争底物和粘附位点以及降低结肠腔内pH值来对病原体产生抗菌作用。
研究发现,降低结肠内pH值可抑制肠杆菌科(Enterobacteriaceae)和梭状芽胞杆菌(Clostridia)等pH敏感有害菌群的过度增殖,并通过抑制前致癌物的形成来降低结直肠癌的风险。
✦抑制有害菌的生长
与上述发现一致,紫甘薯中提取的花青素增加了最知名的益生菌之一双歧杆菌(Bifidobacterium)和乳杆菌(Lactobacillus)的种群数量,但减少了溶组织梭状芽胞杆菌(C. histolyticum)和拟杆菌/普雷沃氏菌的种群数量。
溶组织梭状芽胞杆菌是与感染和炎症性肠病相关的最常见致病性物种之一。
▼
虽然许多研究表明花青素及其代谢产物促进有益菌群生长,抑制有害菌群生长,但对结肠细菌的促进作用存在差异。
对微生物群的不同影响可能是由于源自花青素的代谢物的差异。
花青素(单独和混合物)和没食子酸的孵育被证明可以增加双歧杆菌和乳酸杆菌的数量。而混合花青素和没食子酸显著诱导奇异菌属(Atopobium)的生长,并减少了潜在致病性溶组织梭菌。
没食子酸似乎是本研究中的关键代谢物,可抑制溶组织梭菌增殖。
在其他研究中,研究了没食子酸对梭菌、大肠杆菌和金黄色葡萄球菌等有害微生物的抗菌活性。
花青素对肠道菌群的调节作用



Liang A,et al.Crit Rev Food Sci Nutr.2023
▼
花青素的发酵有助于短链脂肪酸的产生,主要是乳酸、乙酸、丙酸和丁酸。短链脂肪酸通过调节细菌毒力基因表达和降低结肠中管腔pH值来抑制肠道病原体如沙门氏菌(Salmonella spp.)、大肠杆菌(E. coli)和李斯特菌(Listeria spp.)的入侵。
此外,短链脂肪酸在肠道生理事件中发挥着重要作用,如获取能量、调节pH、参与肠脑轴和保护肠道屏障功能。
花青素与肠道菌群作用的研究证据
花青素与肠道微生物的相互作用已经通过动物模型得到了广泛的研究。用黑甜樱桃对小鼠进行饮食干预发现,除了双歧杆菌和乳杆菌外,樱桃花青素的补充还促进了结肠中阿克曼氏菌(Akkermansia)的增殖。
阿克曼氏菌的丰富度被认为可以刺激粘液分泌,改善肠道屏障功能,从而降低炎症风险和免疫介导的疾病,如乳糜泻。
促进Coriobacteriaceae有助于健康
对小鼠进行为期六周的多种浆果饮食干预,结果发现放线菌(Actinobacteria)数量有所改善,这与补充蓝莓的研究结果一致。
放线菌门下的一些科,如双歧杆菌和Coriobacteriaceae,已被证明对健康有益。特别是,有人认为,在高脂肪饮食存在的情况下, Coriobacteriaceae在降低肥胖和肝脏相关疾病的风险方面发挥作用。
双歧杆菌和乳酸杆菌的丰度增加
与体内试验的结果一致,在参加为期六周的25克蓝莓能量饮料的健康男性中发现双歧杆菌和乳酸杆菌的丰度增加。
花青素、其代谢物和肠道微生物群间的相互作用
Liang A,et al.Crit Rev Food Sci Nutr.2023
小结
体外、动物和人体研究一致认为,花青素及其代谢物能够调节肠道微生物群。花青素会促进益生菌的生长,特别是双歧杆菌、乳杆菌和阿克曼氏菌,它们通过产生短链脂肪酸或通过竞争底物和定植位点来对抗有害和致病性菌株,从而有利于宿主健康。
然而,花青素对肠道微生物群的调节也可能依赖于其他相关的化学物质,如葡萄糖,因为与那些喂食高葡萄糖对照饮食的大鼠相比,在蓝莓补充后,在肠道中发现了较低水平的乳酸杆菌。
此外,不同的动物模型和志愿者群体在微生物组成方面存在较大差异,这是这些研究的另一个挑战。
因此,未来的临床研究重点应放在不同年龄和生理阶段的男性和女性不同亚类人群上,这可能会更深入地了解花青素在微生物群调节中的作用及其对特定人群的健康益处。
前面讲述了花青素对肠道菌群的调节作用,下面谷禾来具体讲讲花青素和肠道微生物群相互作用下对一些疾病的影响
肥胖和超重被定义为对健康构成威胁的脂肪组织的过度积累。脂肪组织中的代谢紊乱,如炎症、脂肪因子释放失调和脂肪生成增加,是肥胖和其他肥胖相关疾病的一些常见潜在特征。
有证据表明,与肥胖相关的疾病可能部分由肠道生态失调引起,包括微生物组成及其代谢物的变化。
最近,花青素与肠道微生物的相互作用被证明具有改善肥胖的作用。主要通过调节脂肪生成和脂肪分解,炎症反应的介导以及通过肠脑通讯控制食欲。
1
肠道微生物组影响脂质代谢,肠道菌群失调与血脂异常和肥胖等有关。
最近的一项体内研究显示,对高脂喂养的小鼠补充了200毫克/公斤体重的紫甘薯花青素(花青素和芍药苷酰基糖苷),改善了高脂饮食引起的体重增加和肝脏总脂肪积累。
√调节AMPK通路减少脂肪积累
他们认为,花青素抑制脂肪堆积的作用需要激活小鼠肝脏细胞AMPK通路。此外,在摄入花青素后,AMPK途径下游靶点乙酰辅酶a羧化酶、脂肪酸合成酶、甾醇调节元件结合蛋白也被下调,以减少小鼠肝细胞中的脂肪积累。
已知AMPK通路的上调在抑制脂肪生成中发挥重要作用,例如抑制脂蛋白脂肪酶 (LPL),这是一种可受肠道微生物群调节的酶。肠道菌群通过影响抑制脂蛋白脂肪酶,以相反的方式促进脂肪细胞中甘油三酯的储存。而活化的AMPK及其下游信号通路至少部分抵消了肠道微生物群促进的能量储存。
√操纵法尼醇X受体信号减轻代谢受损
此外,已知肠道菌群产生的次生胆汁酸通过胆汁酸受体法尼醇X受体(FXR)改变肝脏脂质代谢。胆汁酸-结肠微生物群相互作用的破坏可能导致肥胖易感性。
花青素被证明可以操纵肠道微生物群/法尼醇X受体信号,从而减轻高脂肪诱导的肝脏脂肪积累和脂质代谢受损。
黑米花青素(0.48g/kg)喂养12周使血清乳酸脱氢酶(LDH)、丙氨酸转氨酶(ALT)和天冬氨酸转氨酶(AST)含量正常化,这些指标是肝脏脂肪过度积累影响肝损伤的指标。
这些发现表明,膳食花青素可能通过改变肠道菌群来减轻高脂饮食引起的脂质代谢紊乱。
2
肠道微生物群参与调节炎症,在肥胖患者的脂肪组织中通常观察到系统性的低级别炎症并发症。
√经常摄入花青素的肥胖患者炎症标志物含量下降
在一项研究中,代谢综合征患者每天食用400克新鲜越桔,持续12周后,血清脂多糖、IL-6、IL-12和高敏C反应蛋白(hsCPR)浓度下降。
hsCPR是一种与肥胖相关的炎症标志物。
另一项动物研究显示,花青素的干预通过抑制21天大的小鼠白色脂肪组织中TLR4介导的NF-κB信号通路,降低了高脂肪诱导的促炎标志物IL-6和肿瘤坏死因子的表达。
此外,高脂肪喂养诱导下的粪便双歧杆菌减少,在妊娠期和哺乳期用花青素处理后恢复。
3
肠道微生物在调节涉及能量稳态的神经激素活动中发挥作用。由微生物群产生的花青素和短链脂肪酸会影响促食欲激素和厌食激素的释放。
√刺激肠促胰岛素激素分泌减少食物摄入量
飞燕草素3-芸香糖苷被证明可刺激释放胰高血糖素样肽-1,这是一种肠促胰岛素激素,可促进胰岛素分泌和饱腹感,从而减少食物摄入量。
√减轻肥胖引起的神经激素紊乱
肥胖会通过表观遗传修饰相关基因表达导致参与能量平衡的神经激素活动紊乱。食用花青素可能会恢复肥胖引起的这些不利影响。
在肥胖大鼠中,连续8周食用额外的樱桃和桑葚花青素可以减少体重增加,降低血清瘦素水平,增加脂联素。
与先前的研究一致,肥胖患者在食用富含花青素的水果6周后,发现瘦素受体表达增加,这表明补充花青素后瘦素敏感性有所改善。
此外,高脂饮食诱导的肥胖大鼠大脑皮层和海马体中的γ-氨基丁酸(GABA)水平下降。研究报道,与对照组相比,40天摄入黑大豆花青素减少了成年大鼠的体重增加。高剂量花青素可降低促氧神经肽Y (NPY)的表达,提高下丘脑GABA受体B (GABAB1R)的表达。
GABA是中枢神经系统中的一种抑制性神经递质,通过控制神经元的兴奋性、可塑性和额叶皮质的同步性,在摄食反应中起关键作用。
综上所述,花青素似乎通过多种方式与肠道微生物群相互作用,包括调节脂质代谢、炎症反应和参与能量平衡的神经激素活动,从而具有改善肥胖的作用。
注:关于健康和肥胖个体肠道中花青素代谢的差异,人们的理解有限。因此,未来的研究重点应放在花青素结肠代谢物及其对健康和肥胖受试者的健康影响上。
2型糖尿病是一种以高血糖为特征的代谢性疾病,由胰岛素抵抗逐渐发展而来。
许多研究已经报道了花青素及其代谢物的抗糖尿病潜力,这可能归因于以下单一或同时的作用,包括增强胰岛素敏感性,改善β细胞功能和延迟餐后血糖反应。
1
花青素可以通过自身或其微生物代谢物改善胰岛素敏感性。
脂肪组织能够分泌脂联素等脂肪细胞因子来调节葡萄糖稳态。
√花青素可以诱导脂肪细胞摄取葡萄糖
花青素及其微生物代谢物原儿茶酸已被证明可以通过上调葡萄糖转运蛋白4来增加人类脂肪细胞去除血浆葡萄糖的能力。
PPARγ的敲除消除了花青素的胰岛素增敏特性,这表明PPARγ可能在增加花青素诱导的脂肪细胞葡萄糖摄取中发挥作用。
PPARγ是一种参与葡萄糖处理和葡萄糖转运蛋白调节的核转录因子
√花青素还可以介导骨骼肌中的葡萄糖摄取
除了脂肪细胞外,骨骼肌中的葡萄糖摄取也可由花青素介导。
糖尿病小鼠口服紫玉米花青素提取物8周后,骨骼肌对葡萄糖的摄取增加,因为在肌肉细胞中发现葡萄糖转运蛋白4表达增加,并伴随AMPK通路的激活。增加AMPK活性是刺激葡萄糖转运蛋白4从细胞内囊泡转移到细胞膜的重要途径,从而促进葡萄糖流入细胞。
此外,在摄入原儿茶酸后大鼠肝脏中发现胰岛素受体底物1/磷脂酰肌醇3-激酶/蛋白激酶B-2 (IRS1/PI3K/AKT2) 信号通路的活性增加,表明肝脏胰岛素敏感性得到改善。
证据表明,花青素及其降解产物改善了脂肪细胞和肌肉细胞的胰岛素敏感性。
2
花青素还可以通过消除高血糖引起的氧化应激来保护胰腺β细胞的功能。
√改善高糖影响下的胰岛β细胞损伤
发现富含花青素的提取物可减轻高糖诱导的β细胞损伤,保护其分泌功能,并导致血清胰岛素水平升高。
一项体外研究(对与桑椹花青素孵育的高糖培养的β细胞)发现,花青素能够通过抑制β细胞中的凋亡相关蛋白,包括丝裂原活化蛋白激酶、细胞色素c和Bcl-2家族蛋白,消除高糖诱导的细胞内活性氧水平升高,并将β细胞活力恢复到84%。
除了完整的花青素外,丁香酸是花青素的主要微生物代谢产物之一,口服30天可改善四氧嘧啶诱导的糖尿病大鼠的胰岛素释放功能异常、血糖和糖蛋白水平升高。
丁香酸的抗糖尿病作用可能与保持β细胞的活力和功能有关。
3
在人体试验中也观察到了膳食花青素和微生物代谢物在调节餐前和餐后血糖中的作用,从而降低了患2型糖尿病的风险。
众所周知,血糖水平是动态的,全天都在变化。空腹和餐后血糖水平是诊断糖尿病的两个重要指标。空腹血糖值代表身体在一夜禁食后如何管理血浆葡萄糖,而餐后血糖则代表餐后的血糖反应。
√多摄入花青素表现出延迟的血糖反应
与未补充花青素的人相比,额外摄入150克混合浆果泥的参与者表现出延迟的血糖反应。
研究人员解释:餐后血糖降低至少部分是由于花青素对α-葡萄糖苷酶的抑制作用,α-葡萄糖苷酶是一种释放α-葡萄糖的酶。
该研究结果支持以下机制:花青素通过抑制胰腺α-淀粉酶和α-葡萄糖苷酶的活性,以及干扰葡萄糖转运蛋白(SGLT1和GLUT2),因为这些转运蛋白存在竞争关系,从而降低餐后血糖。
√降低了空腹血糖浓度
血清马尿酸是一种花青素的结肠代谢产物,在成年人食用越橘8周(200克/天)后显著增加。8周越桔 干预也降低了空腹血糖浓度。
然而,没有观察到血浆马尿酸与餐后血糖浓度之间存在任何相关性。
花青素可能通过抑制α-淀粉酶和α-葡萄糖苷酶活性以及与葡萄糖载体相互作用来调节餐后血糖,而花青素代谢产物马尿酸则由于微生物降解物的延迟出现而与改善空腹血糖控制有关。
小结
花青素及其代谢物的抗糖尿病作用已经在体外和体内研究中得到阐明。主要通过调节葡萄糖转运蛋白表达和易位来改善胰岛素抵抗,对胰岛β细胞及其分泌功能的保护作用,以及调节空腹和餐后血糖水平。
因此,由富含花青素的食物或补充剂组成的饮食干预可能是更好地控制患有2型糖尿病高风险个体和糖尿病患者血糖的替代途径。
心血管疾病是一类影响心脏和血管的疾病,最常见的形式包括动脉粥样硬化、冠心病、心律失常、心力衰竭和中风。
流行病学研究表明,高花青素摄入量与心血管疾病风险降低有关。花青素及其代谢物发挥保护作用,主要归因于抑制血小板过度活跃,缓解内皮功能障碍,改善脂质谱和逆转胆固醇转运。
1
花青素和原儿茶酸已被证明稳定血小板活性,从而防止血栓形成。
√摄入花青素抑制了血小板的过度聚集
一项研究发现,100μM和500μM没食子酸可以减少刺激物(二磷酸腺苷和血栓素A2激动剂U46619)诱导的血小板聚集、血小板-白细胞聚集和P-选择素表达。
P-选择素(一种参与细胞表面粘附的糖蛋白)的表达减少,表明血小板稳定性提高,血栓形成风险降低。
没食子酸抗血小板聚集的潜在机制是通过降低细胞内钙水平和下调蛋白激酶c-α(PKCα)/p38丝裂原活化蛋白激酶(p38MAPK)和Akt/糖原合成酶激酶3β (GSK3β)通路的活性。
另一项临床试验也支持了这种抗血栓作用,该试验发现,在健康志愿者中,200mL富含花青素和芸香苷的李子汁干预28天,可有效抑制人工刺激的血小板聚集,并降低P-选择素的表达。
此外,补充花青素28天后还观察到氧化应激生物标志物丙二醛水平降低,这表明血小板的抗机能亢进至少部分归因于花青素及其代谢物的抗氧化能力。
2
花青素及其代谢物具有血管内皮保护作用,内皮功能障碍被认为是动脉粥样硬化发展的早期指标。内皮功能受损可破坏血管收缩和血管舒张的止血作用,引发一系列生理事件,如促炎因子分泌、白细胞粘附等,从而引发动脉粥样硬化。
√减缓动脉粥样硬化
研究了花青素代谢物原儿茶酸对载脂蛋白e缺乏小鼠的内皮保护作用。原儿茶酸(0.03g/kg)喂养20周小鼠,动脉粥样硬化形成延迟,血管细胞粘附分子1 (VCAM-1)和细胞间粘附分子1(ICAM-1)表达显著降低。
这些粘附分子的减少可以减少单核细胞对血管内皮的附着,从而减缓动脉粥样硬化的早期发展。
√改善血管舒张情况
蓝莓花青素的血管保护特性已在临床试验中得到证实。健康志愿者在摄入蓝莓花青素(766-1791毫克)1至2小时和6小时后,内皮依赖性血管舒张得到改善。
3
许多研究也报道了富含花青素的饮食对血脂的有益影响。
√直接补充花青素缓解高脂血症
在高脂饮食诱导的肥胖小鼠中,每天补充150 mg/kg黑加仑花青素12周后,高脂血症得到缓解,相应的血清总胆固醇、甘油三酯和低密度脂蛋白水平降低。
另一项研究报告,50mg/kg原儿茶酸干预10周后,降低了先前由脂肪和果糖丰富的饮食引起的高脂血症雄性大鼠的血清甘油三酯、总胆固醇和低密度脂蛋白水平。
三项新指标,即血浆动脉粥样硬化指数(AIP)、动脉粥样硬化指数(AI)和冠状动脉危险指数(CRI),在50mg/kg原儿茶酸治疗后分别显著降低33%、52%和36%。
√通过调节菌群降低血脂
此外,花青素干预后观察到嗜黏蛋白阿克曼菌(Akkermansia muciniphila)和狄氏副拟杆菌(Parabacteroides distasonis)的富集,其与循环甘油三酯和低密度脂蛋白水平呈负相关。已知阿克曼菌可以积极调节肠道屏障完整性,防止脂多糖易位诱导炎症。
低密度脂蛋白(LDL)及其氧化修饰(oxLDL)通过刺激泡沫细胞的形成与动脉粥样硬化的发病机制有关。
最近发现狄氏副拟杆菌(Parabacteroides distasonis)通过产生琥珀酸和次生胆汁酸,如石胆酸和熊去氧胆酸,具有降血脂作用。
综上所述,花青素及其代谢物可能通过改善血脂来预防动脉粥样硬化和冠状动脉疾病。
小结
综上所述,花青素及其代谢物通过维持血小板稳定、保护血管内皮功能、改善血脂等发挥心血管保护作用。
然而,关于花青素心血管保护作用的人类和动物研究之间存在差距。因为动物中使用的浓度可能无法在人体中达到生理水平。
此外,临床研究的实验持续时间通常在4至12周之间,这可能无法捕捉到长期补充的影响。因此,未来可能需要进行长达一年的长期饮食干预研究。
帕金森病、阿尔茨海默病和肌萎缩侧索硬化症等神经退行性疾病是发生在神经系统中的疾病,导致日常活动协调困难、认知障碍和记忆丧失。
这些疾病的病因可能多种多样,但它们有一系列共同的潜在因素。这些因素包括但不限于长时间的氧化应激、中枢神经系统炎症、神经元钙信号失调和兴奋性毒性(即神经毒性)。
有证据表明,一些花青素代谢物,如原儿茶酸、香草酸、没食子酸和其他简单的酚酸,可以通过血脑屏障进入中枢神经系统,这表明这些物质可能直接在发生神经系统疾病的中枢神经系统中发挥作用。
1
花青素肠道代谢物与防止神经毒性和维持Ca2+体内平衡有关。
√没食子酸可以减轻谷氨酸诱导的神经毒性
谷氨酸是一种兴奋性神经递质,有助于大脑正常功能,而谷氨酸水平异常升高可引起谷氨酸兴奋性毒性。谷氨酸兴奋性毒性被认为是神经退行性疾病的主要原因之一。
没食子酸是能够穿透血脑屏障的代谢物之一。报道,25和50μg/ml 没食子酸预处理2小时可导致大鼠皮层神经元细胞中N-乙酰天冬氨酸(NAA)水平和微管相关蛋白2(MAP-2)表达增加,表明没食子酸处理减轻了谷氨酸诱导的大鼠神经元神经毒性。
在缺乏没食子酸的细胞中观察到的NAA和MAP-2的下调被认为是神经元损伤和丧失的直接指标。
此外,在没食子酸处理的细胞中也观察到钙结合蛋白(一种能够缓冲Ca2+内流的钙结合蛋白)的上调,表明没食子酸在保护神经元免受谷氨酸引发的钙失衡方面的作用。
√天竺葵素代谢物减轻神经兴奋性毒性
观察到4-羟基苯甲酸(HBA)是一种天竺葵素的代谢物,可在体外保护大鼠小脑颗粒神经元免受谷氨酸兴奋性毒性。当仅用谷氨酸处理的大鼠小脑颗粒神经元导致近50%的细胞损失时,4-羟基苯甲酸处理保留了细胞并且只有13%的细胞死亡得到证实,揭示了4-羟基苯甲酸在谷氨酸兴奋性毒性缓解中的作用。
注:原儿茶酸和4-羟基苯甲酸似乎发挥互补作用,表明它们的共同治疗可能比单一代谢物更有效。然而,本研究并未提供原儿茶酸和4-羟基苯甲酸的联合治疗,因此其最佳效果的混合比例值得进一步研究。
2
花青素及其代谢物的神经保护作用与它们在防止蛋白质聚集和促进自噬中的潜在作用有关。
蛋白质聚集被认为刺激神经元变性,与许多神经退行性疾病有关。例如,由聚集的淀粉样蛋白(Aβ)形成的斑块和由tau蛋白形成的神经原纤维缠结是阿尔茨海默病的标志。导致路易体形成的α-突触核蛋白(αS)的异常组装与帕金森病有关。
√原儿茶酸防止蛋白质异常聚集
48小时原儿茶酸处理(10至100 μM)对Aβ和αS原纤维聚集的抑制率分别为30–79%和70–80%。原儿茶酸治疗6天(100μM)也使预先形成的Aβ原纤维和αS原纤维不稳定65%和80%。
这些结果表明,原儿茶酸不仅能抑制Aβ和αS纤颤,还能逆转预先形成的纤颤,它们共同调节蛋白质相互作用,防止蛋白质异常聚集导致的细胞死亡
√香草酸的干预有助于治疗阿尔兹海默病
香草酸是黑芝麻色素释放的主要代谢物,它显示出对β-淀粉样蛋白的抑制活性。在用6μM香草酸处理24小时后发现一半(50%)的抑制,在50μM香草酸孵育时达到90%的抑制。
此外,较高浓度的香草酸(60μM)可使β-分泌酶活性降低60%。β-分泌酶是一种在神经元中催化淀粉样前体蛋白合成淀粉样肽的第一步酶,促进蛋白质聚集。抑制β-分泌酶活性被认为是治疗阿尔茨海默病的另一种方法。
3
花青素微生物降解产物的有益作用已通过调节神经元细胞凋亡信号通路被发现。诱导细胞死亡是神经退行性疾病的最终病理活动。
√花青素代谢物调控细胞凋亡信号
原儿茶酸在最近的动物研究中显示出抗凋亡能力。原儿茶酸通过上调大脑皮层中参与抗氧化活性的基因,如Sod2、Cat、Gpx1和Gsr,来阻止镉诱导的细胞凋亡。
原儿茶酸处理可以减轻镉引起的促炎生物标志物肿瘤坏死因子和IL-1β的表达增加,这表明原儿茶酸通过介导促凋亡蛋白和抗凋亡蛋白,增强抗氧化防御系统和减少炎症细胞因子的产生来保护皮质组织免受镉损伤。
总之,这些发现表明花青素代谢物不仅可以减轻导致神经元死亡的各种诱导剂,还可以调节凋亡信号,从而在多个层面上降低神经退行性病变的风险。
√花青素介导的肠道菌群变化减少了神经退行性疾病的风险
除了花青素及其代谢物有助于神经保护作用外,花青素介导的肠道微生物群变化间接减少了神经退行性疾病的发生。
研究表明,由于脂多糖诱导的神经炎症反应、血脑屏障渗漏增加和β-淀粉样蛋白错误折叠的发生率,肠道生态失调可能增加神经退行性疾病的易感性。
在高脂喂养大鼠中补充黑莓花青素(25mg/kg)17天可以抵消高脂饮食引起的部分生态失调,例如孢杆菌属(sporobacter)数量增加和瘤胃球菌(Rumminococcus)数量减少。
花青素相关的肠道菌群调节通过犬尿氨酸途径改变色氨酸代谢,从而与抗神经炎症活性相关。这种改变被证明可以增加犬尿氨酸的产生,犬尿酸是一种抗兴奋毒性分子,可以减轻神经退行性病变。
尽管花青素介导的色氨酸代谢对神经保护的证据有限,但肠道微生物群对色氨酸代谢的操纵很可能会影响肠-脑轴。
建议
花青素及其微生物代谢产物可能是治疗神经退行性疾病的有效分子。然而,由于大多数研究结果都是基于体外或动物模型,因此在未来的研究中需要进行大量的临床前和临床研究。
由于衰老是神经退行性变的主要危险因素,因此有必要以老年人为重点补充花青素,以探索其在认知改善中的作用和肠道微生物群。
▼
•花青素抑制了炎性细胞因子的产生
花青素可以通过调节微生物群来预防肠道疾病。在葡聚糖硫酸钠(DSS)诱导的结肠炎小鼠中,用枸杞中的花青素提取物(200 mg/kg/天)喂养持续至少一周,恢复了紧密连接蛋白的表达,抑制炎症细胞因子(如TNF-α,IL-6)的产生。
同样,食用富含花青素的马铃薯持续9周喂养结肠炎小鼠,缓解了结肠长度的缩短,降低了髓过氧化物酶(炎性粘膜标志物)水平和炎性细胞因子。
•微生物群是花青素调节炎症性肠病的介质
然而,这种作用在抗生素治疗的小鼠身上消失了,强烈表明微生物群作为花青素对炎症性肠病介质的作用。
▼
摄入花青素可通过多种潜在机制降低患非酒精性脂肪性肝病的风险,例如其固有的抗炎作用、脂质代谢和葡萄糖代谢的管理以及改变微生物群特性。
•改变了脂肪肝小鼠体内的微生物群
其他研究也证明了花青素改变微生物群的作用,但花青素来源的多样性可能导致不同的微生物种群发生改变。在高脂饮食诱导的非酒精性脂肪肝变性小鼠中,摄入桑树水提取物(0.2-0.6%)可减少促炎的Turicibacteres,并提高拟杆菌(Bacteroidales)与梭菌(Clostridiales)的比例。
总的来说,花青素的摄入可能会改变肠道微生物群的组成,从而改善肠道屏障功能和能量消耗,降低肥胖、炎症、胰岛素抵抗,并最终降低非酒精性脂肪性肝病及其代谢并发症的风险。
▼
先前的动物研究已经证明了花青素通过微生物群调节途径抑制肿瘤生长和癌症进展的潜力。
•花青素治疗后肿瘤计数显著降低
在结直肠癌小鼠模型中,用覆盆子花青素治疗可显著削弱结肠癌的发生,每只动物的肿瘤计数减少约90%,肿瘤发病率降低(从100%降至72%左右)。
肠道生态失调可能与结直肠癌的增加有关,而花青素似乎可以恢复这种平衡,并通过促进乳酸菌的生长、抑制肠球菌和脱硫弧菌来调节微生物群。
此外,口服越桔花青素可能通过微生物群加速免疫检查点抑制剂(即程序性死亡配体1,可以限制T免疫细胞反应激活的蛋白质)的肿瘤抑制特性。
研究人员提出,补充花青素可改善肠道物种多样性,诱导梭状芽孢杆菌、约氏乳杆菌和毛螺菌科的丰度增加。
▼
花青素有可能降低慢性肾脏疾病的风险。慢性肾脏疾病的特点是肠道生态失调和由此产生的尿毒症毒素。
在一项研究中,花青素显示出通过AMPK磷酸化减轻培养细胞中糖尿病肾病的能力,并在腺嘌呤诱导的慢性肾病小鼠模型中减轻症状。
▼
花青素和富含花青素的食品的抗炎作用也与减缓骨关节炎和骨修复的进展有关,主要是通过抑制MAPK信号通路和NF-kB信号通路。
认识了花青素对人体健康的重要性,那么我们在日常生活中应该如何去补充呢,谷禾在这里列出了一些建议。
水果:存在于葡萄、蓝莓、桑葚、血橙、草莓、樱桃、红心火龙果、山楂等水果中,与水果色泽有一定的关系,在外观上有所体现,水果是补充花青素的较好选择;
蔬菜:如紫甘蓝、紫薯、紫玉米、紫土豆、黑枸杞、茄子、紫苏、甜菜、萝卜、紫洋葱等蔬菜中,也含有较多的花青素;
谷物:大麦、高粱、黑米,以及豆类如绿豆、红豆、黑豆、黄豆、赤小豆等,其中也含有较多的花青素。
花青素易受热、光、氧等因素的影响而降解,因此建议选择新鲜的食物食用,以保证花青素的含量。
花青素在高温下易被破坏,因此建议采用轻微加热的烹饪方式,如蒸、煮、炖等,以尽可能保留花青素的含量。
花青素与其他营养素的搭配可以增强其吸收和利用效果。例如,与维生素C搭配可以增强花青素的抗氧化作用。
如果日常饮食中花青素的摄入量不足,可以考虑适当补充花青素保健品。但是,需要注意选择正规品牌和适量摄入。
虽然花青素对人体健康具有多种保护作用,但过量摄入也可能对健康造成负面影响。
因此,建议适量摄入花青素,不要过度依赖花青素保健品等补充剂。
总之,通过日常饮食摄入富含花青素的食物,可以有效补充花青素,发挥其保健作用。同时,需要注意花青素的摄入量和搭配方式,以充分发挥其保健作用。
总的来说,花青素的健康益处已被广泛研究,它们被认为可以有效减轻或预防一些疾病。
通过与肠道微生物相互作用了解花青素及其健康益处是很重要的。肠道菌群在花青素降解过程中发挥重要作用,并介导多种生理活动。
花青素也会改变肠道微生物组成,维持微生态平衡,操纵短链脂肪酸的产生、脂肪代谢、肠道上皮功能、与中枢神经系统的联系和免疫反应。
因此,花青素的潜在益处可能间接归因于其对肠道微生物群的调节作用及其随之而来的生理影响。由于这些原因,研究花青素和肠道微生物群的相互作用在慢性疾病预防和治疗中是很重要的。
主要参考文献
Liang A, Leonard W, Beasley JT, Fang Z, Zhang P, Ranadheera CS. Anthocyanins-gut microbiota-health axis: A review. Crit Rev Food Sci Nutr. 2023 Mar 17:1-26.
Akbari, G. 2020. Molecular mechanisms underlying gallic acid effects against cardiovascular diseases: An update review. Avicenna Journal of Phytomedicine 10 (1):11–23.
Agustí, A., M. P. García-Pardo, I. López-Almela, I. Campillo, M. Maes, M. Romaní-Pérez, and Y. Sanz. 2018. Interplay between the gut-brain axis, obesity and cognitive function. Frontiers in Neuroscience 12:155.
Cortés‐Martín, A., C. E. Iglesias‐Aguirre, A. Meoro, M. V. Selma, and J. C. Espín. 2021. Pharmacological therapy determines the gut microbiota modulation by a pomegranate extract nutraceutical in metabolic syndrome: A randomized clinical trial. Molecular Nutrition & Food Research 65 (6):2001048. 2001048.
Drago, L. 2019. Probiotics and colon cancer. Microorganisms 7 (3):66.
Foley, M. H., S. O’Flaherty, R. Barrangou, and C. M. Theriot. 2019. Bile salt hydrolases: Gatekeepers of bile acid metabolism and host-microbiome crosstalk in the gastrointestinal tract. PLOS Pathogens 15 (3):e1007581.
Hampel, H., R. Vassar, B. De Strooper, J. Hardy, M. Willem, N. Singh, … A. De Vos. 2020. The β-secretase BACE1 in Alzheimer’s disease. Biological Psychiatry 89 (8):745–756.

谷禾健康
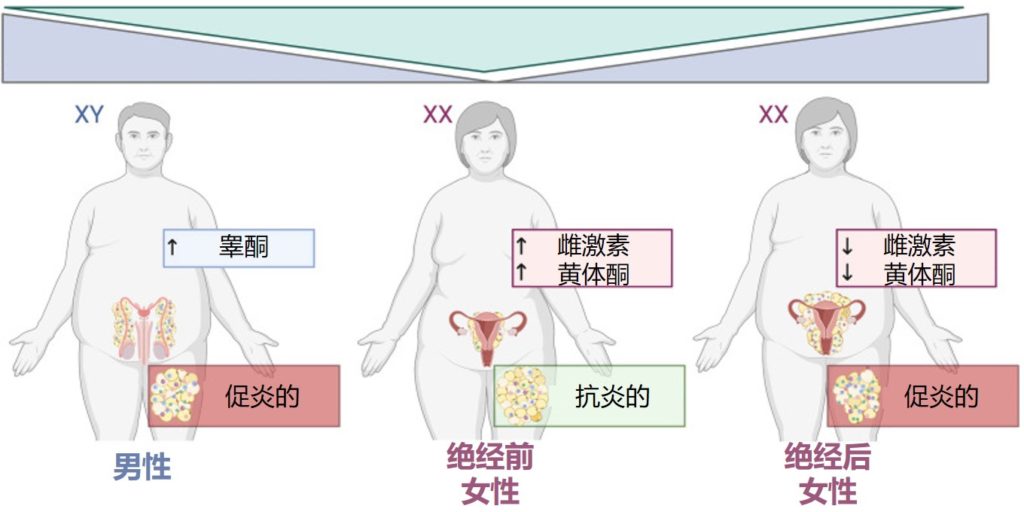
我们的身体中有很多不同的器官,组织,腺体等会产生许多信号分子来精确控制和影响身体的反应和活动,这些信号分子包括激素、神经递质、生长因子、细胞因子等。它们可以促进或抑制细胞的生长和分化,调节细胞间的相互作用和通讯,影响身体的代谢和能量平衡,参与免疫系统的细胞和分子相互作用,调节身体的内环境和外环境的适应性等。
其中,激素是一类重要的生物活性物质,也可以说是信号分子,一般由特定的内分泌细胞,腺体或器官所产生,包括性激素、脂质激素、蛋白质激素、胰岛素、甲状腺激素、肾上腺素和去甲肾上腺素等。
这其中性激素是一类重要的激素,主要由生殖腺(如卵巢、睾丸)和肾上腺产生,包括雌激素、孕激素和雄激素等。它们在生殖系统中发挥重要作用,对女性月经周期、妊娠、分娩、哺乳等过程起调节作用,对男性性发育、精液生成、性欲等方面也具有重要影响。此外,性激素还与骨密度、心血管健康、认知能力等方面有关。
临床上性激素六项是生殖内分泌门诊常用的检查。性激素六项检查主要用于评估男女生殖内分泌功能,对月经失调、性功能障碍、不孕不育、生殖系统肿瘤、性染色体病等的鉴别和诊断及内分泌治疗的效果检测都具有重要的临床意义和参考价值。
近年来,越来越多的科学证据表明,肠道菌群与性激素(宿主的内源激素或外源补充激素)存在直接或间接的相互作用。性激素对于塑造肠道微生物群组成具有一定影响,与此同时,肠道微生物群本身也会影响性激素水平。
目前的研究主要基于横断面研究,基于纵向的检测肠道菌群发育以及激素水平变化的研究队列还较少,目前无法区分是性激素变化在前还是菌群变化在前,但是肠道菌群的失调和性激素的异常水平都与多种疾病的发生和发展有关,如肥胖症、炎症性肠病、乳腺增生、甲状腺疾病,妇科疾病、自身免疫疾病等。因此,保持肠道菌群和激素水平的平衡,对于维持人体的健康十分重要。
本文旨在阐述激素与肠道微生物群的相互作用,主要着重讲述性激素影响下的人体一些疾病和菌群的差异。
这些发现可能会在未来用于开发治疗激素紊乱相关疾病或与性别和激素活动相关的自身免疫性疾病、甲状腺疾病、代谢疾病甚至是压力等情绪状态的新疗法。
目录/contents
Part1:肠道微生物与激素的关联
Part2:肠道微生物通过激素对宿主的影响
Part3:性别差异下的肠道微生物群
Part4:肠道菌群与性激素的相互作用
Part5:性激素影响下的相关疾病
Part6:结语
肠道微生物群以多种方式影响动物和人类的行为,甚至可能影响人类的情绪状态和疾病状态。最近揭示了菌群的一个关键作用:调节激素分泌。
微生物和激素之间的相互作用会影响宿主的新陈代谢、免疫力和行为。这种相互作用是双向的,因为微生物群已被证明既受宿主激素影响又影响宿主激素。
肠道微生物群通过激素影响宿主情绪、免疫及代谢等
微生物群对宿主激素水平的影响可能是直接的,即微生物群产生激素,也可能是间接的,即微生物可能调节肾上腺皮质(控制焦虑和压力反应)的功能,或调节炎症和免疫反应。
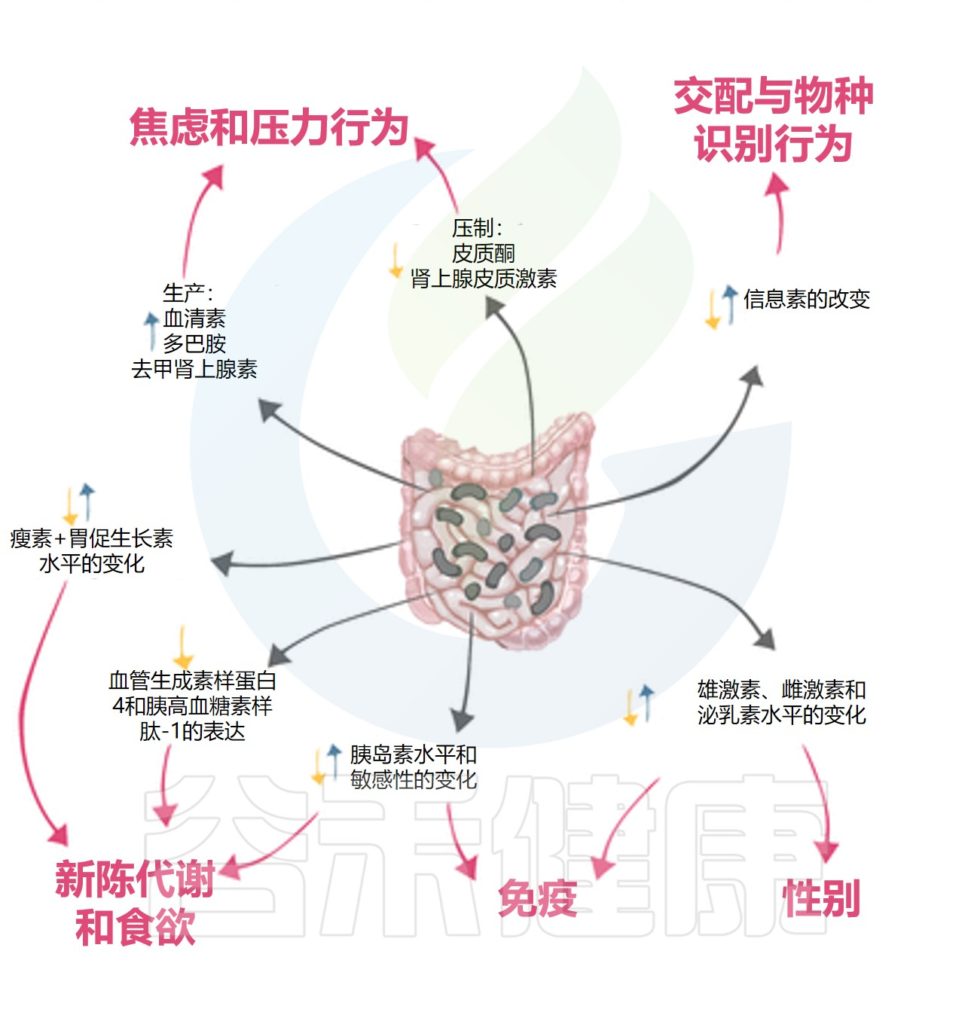
Neuman H,et al.FEMS Microbiol Rev.2015
激素水平调节微生物群的生长、毒性
多种宿主因素(如饮食、运动、情绪、健康状况、压力和性别)会导致激素水平发生变化,进而对微生物群产生多种影响(包括生长、毒力和抵抗力)。
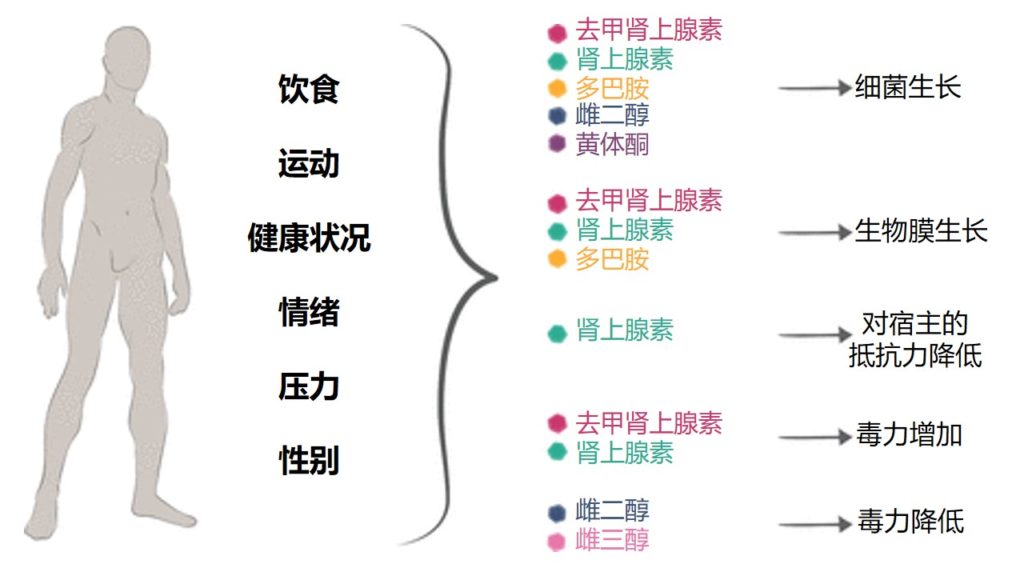
Neuman H,et al.FEMS Microbiol Rev.2015
文献中已记载激素和微生物群之间的相互作用




Neuman H,et al.FEMS Microbiol Rev.2015
▸ 菌群通过影响神经激素前体调节肠道运动、情绪、食欲等
神经内分泌细胞响应神经元输入而分泌神经激素。尽管它们被分泌到血液中以发挥全身作用,但它们也可以充当神经递质。微生物群对行为的调节(例如小鼠的焦虑)被认为是通过神经激素前体(例如血清素、多巴胺)发生的。
最近,肠道细菌被证明可以产生和响应神经激素,例如血清素、多巴胺和去甲肾上腺素。
这些机制对于研究病原体的研究人员来说很有趣,因为它们可能会影响病原体对宿主防御反应的敏感性。例如,为了响应宿主肾上腺素,沙门氏菌会下调其对宿主抗菌肽的抵抗力并诱导关键的金属转运系统,从而影响细胞中的氧化应激平衡。
// 血清素
血清素,也称为 5-羟色胺 (5-HT),是大脑中的主要神经递质之一。然而,超过90%的哺乳动物宿主血清素都存在于肠道中。
肠道血清素的分泌受饮食影响,调节肠道运动、情绪、食欲、睡眠和认知功能。这种双重作用表明血清素可能将肠道(包括其微生物群)与宿主行为联系起来。
血清素与肠易激综合征和克罗恩病有关
大脑血清素可以通过血清素转运蛋白穿过血脑屏障进入血液,这表明肠-脑轴中存在另一个联系。血清素与肠易激综合征和克罗恩病等胃肠道疾病有关,这些疾病也与微生物群的差异有关。
// 多巴胺
多巴胺由芽孢杆菌(Bacillus)和沙雷氏菌(Serratia)等细菌产生 。
无菌小鼠中的游离管腔多巴胺水平显著低于常规小鼠,并且在接种表达β-葡萄糖醛酸酶的细菌后再次升高。
多巴胺不足与帕金森病有关
这些结果表明,在以多巴胺形成不足为特征的帕金森病等疾病中,肠道细菌与多巴胺水平之间可能存在相关性。
//γ-氨基丁酸
γ-氨基丁酸 (GABA) 是哺乳动物中枢神经系统中的主要抑制性神经递质,它也由微生物群产生,并可能影响宿主行为。这很有趣,因为中枢γ-氨基丁酸受体表达的改变与焦虑和抑郁的发病机制有关。
γ-氨基丁酸影响焦虑和抑郁
已经研究了由乳杆菌产生的γ-氨基丁酸,以试图大规模发酵安全的γ-氨基丁酸。因此,向小鼠施用鼠李糖乳杆菌会改变不同中枢神经系统区域中γ-氨基丁酸受体的表达,从而减少与焦虑和抑郁相关的行为。
▸ 菌群通过调节应激激素降低宿主焦虑行为
微生物群可以通过改变应激激素水平来帮助我们保持冷静和平衡。无菌小鼠具有升高的应激激素皮质酮和促肾上腺皮质激素 (ACTH) 血浆水平以响应轻度压力,增加与焦虑和压力相关的行为。
促肾上腺皮质激素通过进一步产生皮质类固醇在下丘脑-垂体-肾上腺轴中发挥重要作用。因此,两种特定物种,瑞士乳杆菌(Lactobacillus helveticus)和长双歧杆菌,降低了大鼠和健康人类的应激激素皮质醇水平和焦虑样行为。
此外,与对照组相比,长期接受益生菌鼠李糖乳杆菌治疗的小鼠在强迫游泳测试中皮质酮水平较低,抑郁行为也较少。
▸ 肠道菌群影响甲状腺激素的转化影响代谢
肠道菌群产生的代谢物对甲状腺和内分泌功能也是必不可少的。它影响硒、铁、锌和碘的吸收,这些都是甲状腺激素产生所需的重要矿物质。它制造短链脂肪酸(SCFAs)与甲状腺激素协同工作,以改善肠道屏障功能和加强细胞间紧密连接。
肠道菌群失衡是影响甲状腺激素转化的主要障碍。事实上,20%的T4(甲状腺素)在肠道中转化为活性的T3(三碘甲状腺原氨酸)形式,肠道菌群不健康,肠道内存在过多的病原体,肠道中甲状腺激素的转化也会减少。
此外当肠道屏障功能减弱时,毒素和未完全消化的食物残渣等颗粒会逃逸,导致肠胃不适和全身炎症。炎症会在组织水平损害甲状腺激素的转化。受体受损,身体也不能将T4激素转化为活性的T3形式。
▸ 细菌通过信息素影响宿主行为
信息素是在性识别、吸引力和交配行为以及攻击行为和支配行为中发挥重要作用的激素。
信息素也称为外激素,一种分泌到一个人体外并影响他人行为的化学物质。
共生细菌和信息素之间的这种联系也发生在哺乳动物身上。两种鬣狗的气味腺中有不同的细菌群落,这与气味分泌物中不同的挥发性脂肪酸特征有关。作者推测,共生细菌产生的代谢产物提供了物种特有的气味。
这进一步支持了细菌可以产生影响宿主行为的气味特征的观点。细菌也可以在配偶选择中发挥作用:雌性小鼠不会被感染沙门氏菌的雄性小鼠的尿液所吸引。
化学信号和嗅觉刺激也在人类行为中发挥作用,未来的研究将帮助我们了解细菌产生的气味是否会影响我们自身的相互作用,甚至可能影响进化。
▸ 肠道菌群代谢性激素影响健康
肠道菌群也影响雌激素,它们通过特定的酶对雌激素进行循环利用并产生雌激素代谢物。当肠道菌群失衡或不健康时,这些过程就会被破坏,从而导致健康问题。
例如,Prevotella intermedius吸收雌二醇和黄体酮,从而促进其生长。雌激素受体ER-β表达的变化也会影响肠道微生物群的组成。
雌二醇在卵巢中合成,是雌激素中含量最高的,活性最强的激素,经肝脏代谢后,通过胆道排泄到胃肠道,部分雌二醇被胃肠道重新吸收进入血液。肠道菌群可以通过改变雌二醇的活性和非活性状态,影响血清中类固醇激素的水平。
武汉大学人民医院转化医学研究所的李艳教授团队在cell metabolism上发表研究论文,发现绝经期前抑郁症患者肠道细菌Klebsiella aerogenes(产气克雷伯氏菌)的3β-羟基类固醇脱氢酶(3β-HSD)通过降解雌二醇诱导绝经期前女性出现抑郁症。
肠道微生物群作为人体最大的菌群库,动物和人类研究发现,肠道微生物群存在性别相关差异。
影响微生物群的重要因素(例如环境或激素)取决于不同的生命周期,并且在男性和女性中会有所不同。
Kim YS,et al.World J Mens Health.2020
需要注意的是,性激素对肠道微生物具有显著的影响,青春期、怀孕、更年期、多囊卵巢综合征和性腺切除术等各种因素会导致性激素水平(睾酮和雌二醇)发生变化,进而导致肠道菌群组成发生变化。
肠道微生物群与性激素之间的相互作用
Santos-Marcos JA,et al.Biol Sex Differ.2023
▷婴儿时期肠道菌群组成与性别相关的差异
婴儿期的特点是参与人乳低聚糖代谢的细菌种类占主导地位(据估计,婴儿细菌微生物群的25-30%来自母乳)并且肠道微生物群多样性的差异减少。
特别是,这个年龄段的特点是被属于双歧杆菌(Bifidobacterium)和链球菌(Streptococcus)的几种物种定殖,例如长双歧杆菌和嗜热链球菌。
6个月至2岁之间的年龄段的特点是将固体食物引入婴儿的饮食中,并伴随着肠道微生物群结构和功能多样性的快速增加。这是婴儿生长的关键时期,肠道微生物群开始接近成人特征。
最近对人类婴儿进行的几项研究发现了他们出生后第一年不同时间点肠道微生物群落的潜在性别依赖差异。
男婴α多样性较低
例如,对出生后前30天新生儿粪便样本的分析显示,与女性相比,男婴的α多样性较低,梭状芽孢杆菌的丰度也较高,而肠杆菌的丰度较低。
男婴双歧杆菌更丰富
在新生儿肠道中最早和最丰富的细菌定植者中,已知双歧杆菌可为宿主肠道提供健康益处。改变在从出生后一天到六个月大,在男性和女性阴道分娩的婴儿中观察到双歧杆菌丰度。发现与女孩相比,男孩的双歧杆菌水平更高。
▷儿童时期肠道菌群组成与性别相关的差异
在出生后的前12个月,婴儿的肠道因饮食中引入固体食物而发生变化,随之而来的是微生物群落的变化。
特别是,在引入固体食物后,婴儿大肠中厌氧菌种群的初始定植在数量和组成上开始与成人相似。几项研究表明,在没有饮食变化或抗生素治疗等外部压力因素的情况下,与儿童相比,成年期的肠道微生物群更加稳定和有弹性。
这表明尽管出现了部分趋同,但仍存在差异,因此分析这一生命阶段与性别差异的关系很有价值。
男孩双歧杆菌与乳酸杆菌之间的正相关性比女孩更强
对来自哥伦比亚两个不同地点的277名1至5岁儿童(154名女孩和123名男孩)的评估(研究临床腹泻的存在)表明,粪便微生物群受到多种宿主因素的影响,包括年龄、健康状况、地点和性别。
有趣的是,与女孩相比,健康男孩的双歧杆菌和乳酸杆菌之间存在显著更强的正相关性,而在受腹泻影响的儿童中未观察到显着相关性。
▷青春期与性别相关的肠道菌群差异最明显!
青春期是发展的关键时期,性激素,如雌激素和睾酮,在这个年龄段人群的肠道微生物群中发挥重要作用。
在性激素的影响下异性间微生物群差距变大
一项针对青春期肠道微生物群组成的人类研究表明,与同性双胞胎相比,青少年(13-17岁)异性双胞胎的粪便微生物群差异更大。
最近的一项研究得出结论,与青春期受试者(5-15岁)相比,非青春期的α和β多样性没有差异。随着青春期的开始,梭状芽孢杆菌(Clostridium)的水平降低,而变形杆菌(Proteus)的丰度增加。
在青春期受试者中,发现Adlercreutzia、Ruminococcus、Dorea、Clostridium和Parabacteroides与睾酮水平有关。
考虑到人类这一生命阶段的可用数据有限——必须使用啮齿动物模型的研究作为替代。在小鼠身上进行的实验表明,肠道微生物群的性别差异出现在与性激素水平相关的青春期开始时。
动物实验中青春期的菌群性别差异
特别是,仅在10至13周龄的非肥胖糖尿病青春期后小鼠中检测到α多样性的差异,与雄性相比,雌性的水平显著更高。
此外,雄性小鼠表现出更高丰度的卟啉单胞菌科(porphyromonas)、韦荣球菌科、消化球菌科、乳杆菌科和肠杆菌科与女性相比。而当雄性小鼠被阉割后,这些差异就消失了。
▷成年后肠道菌群组成与性别相关的差异
肠道微生物组在生命的最初几年大部分时间都不稳定,但在成年期变得更加稳定,之后细菌丰富度和组成可能伴随着特定疾病会发生变化。
在整个生命周期中,随着免疫和神经系统的成熟,肠道微生物群的伴随发育是性别二态性的,导致成年男性和女性的不同微生物群落以及免疫和神经炎症通路。
注:年龄、环境(饮食和身体活动)和地理位置在内的多种因素似乎在这些性别依赖性肠道菌群差异中发挥了作用。
分析来自结肠癌研究对照组的82名受试者得出结论,性别与肠道微生物组的整体组成显著相关。然而,这项研究包括的参与者数量相对较少,年龄范围相当广泛(30-83 岁),这可能混淆了研究的结果。
年轻女性肠道微生物α多样性比同年龄男性更高
在包括美国、英国在内的四个地理区域以及来自哥伦比亚和中国的两个队列中评估了年龄在20至69岁之间的三个大型队列中年龄、性别和肠道微生物群α多样性之间的关系。
有趣的是,与同年龄段的男性相比,年轻的成年女性(20-45 岁)表现出更高的α多样性。此外,性别和α多样性之间的关联在年轻人中比在中年人中更为明显,而当参与者的平均年龄为60岁时,男女之间的α多样性没有差异。这可以解释为更年期女性的雌激素水平下降。而更年期之后男性与女性的激素水平差异变小,肠道微生物群的相似程度更接近。
总之,对来自不同国家的男性和女性进行的人口研究发现人体肠道微生物群组成存在性别相关差异。
这意味着,性激素的动态变化会影响肠道共生,而青春期时激素的变化最大,此时男性与女性的肠道微生物群之间的差异非常明显。
注:由于该研究所覆盖的样本量还较小,年龄之间的跨度比较大,仅作为研究参考,还不能用于诊断标准。
整个生命周期中男性和女性肠道微生物群组成的差异
Valeri F,et al.Front Neuroendocrinol.2021
不仅肠道微生物群受性激素的影响,而且肠道微生物群本身也会影响激素水平。
▷ 微生物代谢雌激素进而影响多个器官
生殖内分泌功能涉及由复杂反馈机制控制的多种激素。卵巢、肾上腺和脂肪组织会产生雌激素。体内产生的或作为食物摄入的雌激素可以被肠道微生物代谢。
产生的代谢物再次影响宿主。性激素通过类固醇受体(包括雌激素受体β)直接调节细菌的新陈代谢。
具有雌激素代谢能力的肠道微生物群基因库
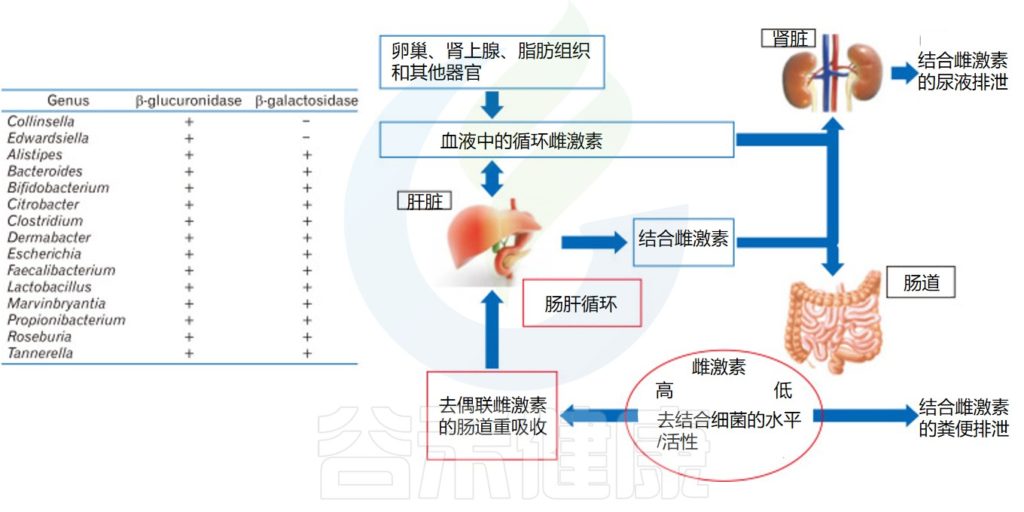
Yoon K,et al.J Neurogastroenterol Motil.2021
同时,具有β-葡萄糖醛酸酶活性的肠道微生物群解离胆汁中排出的结合循环雌激素。去结合使雌激素对系统的重吸收过程成为可能。去结合的雌激素通过雌激素受体循环并影响多个器官,不仅影响生殖器官,还影响骨骼系统、心血管系统和中枢神经系统。
雌激素循环途径
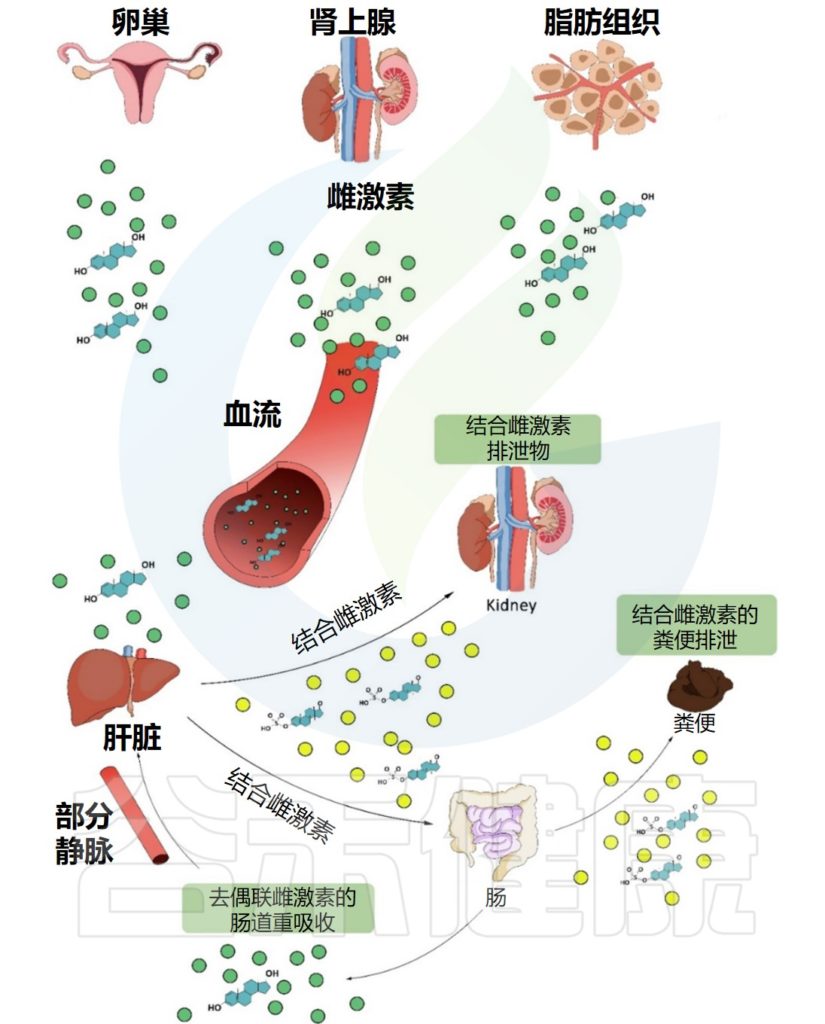
Valeri F,et al.Front Neuroendocrinol.2021
雌激素在几个组织中局部产生(主要是卵巢,但也有肾上腺和脂肪组织)。它们在血液中循环,到达多个靶器官,如肝脏和肾脏。一旦到达肝脏,它们就会通过葡萄糖醛酸化或磺化反应进行结合。
此外,结合雌激素被转化为水溶性分子,随后可以通过胆汁、尿液和粪便排出体外。某些部分的结合雌激素可以在肠道中被肠道微生物分解,并通过门静脉被重新吸收到肝脏中。
雌激素对发病机制具有影响
通常,雌激素与核受体结合,引起构象变化。通过肠道屏障通透性和免疫系统调节,中枢和外周改变与肠易激综合征 (IBS) 相关联。流行病学研究表明女性在肠易激综合征中比例更大,暗示性激素对其发病机制的影响。
雌激素受体作用的分子途径
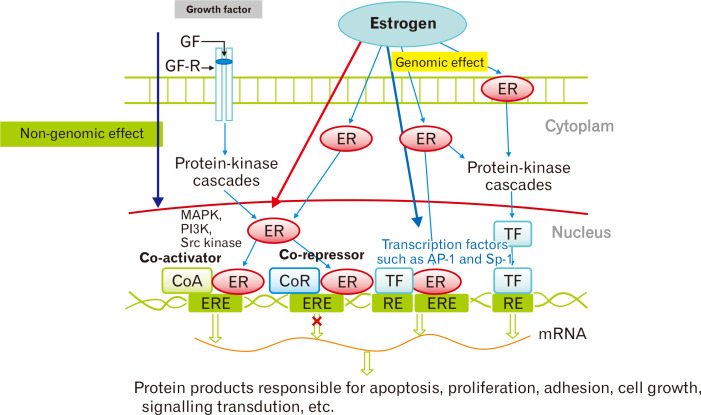
Yoon K,et al.J Neurogastroenterol Motil.2021
▷ 微生物群影响雄激素的代谢
据报道,在雄激素代谢中,肠道微生物群是一个重要的调节因子。在小鼠的小肠内容物中,葡萄糖醛酸化睾酮和双氢睾酮含量很高。远端肠道具有高水平的游离双氢睾酮。
在年轻成年男性的粪便中观察到明显高水平的未结合双氢睾酮。在无菌小鼠中,葡萄糖醛酸化睾酮和双氢睾酮较高;然而,在远端肠道中发现游离水平非常低。这意味着肠道微生物群会影响双氢睾酮和睾酮的肠道代谢。
▷ 性激素高的人拥有更多样化的肠道微生物
血清睾酮升高的男性和雌二醇水平升高的女性分别拥有更多样化的肠道微生物群,许多细菌属与睾酮(Acinetobacter, Dorea, Ruminococcus和Megamonas)和雌二醇(Slackia和Butyricimonas)水平相关。
高雄激素的女性微生物群明显改变
在这方面,患有多囊卵巢综合征(高雄激素)的女性在妊娠早期到晚期的变化中,表现出明显的微生物群改变,变形菌(Proteobacteria)和放线菌(Actinobacteria)总体增加,丰富度降低。
肠道微生物群组成的性别差异在青春期增加,随着青春期的进展,女孩的肠道微生物群变得与成年人更相似。这些结果可能还表明肠道微生物群可能会影响青春期的时间,可能是通过调节宿主性激素水平。
雌激素水平与肠道微生物多样性密切相关
在男性和绝经后女性中,尿液中的雌激素水平与肠道微生物群丰富度和α多样性密切相关,而绝经前女性雌激素水平在月经周期中收集时变化很大,并未显示出这种关联。
▷ 微生物群可以预测粪菌移植后睾酮水平
最近报道,绝经后妇女的肠道菌群与男性比绝经前妇女更相似,绝经后妇女与同龄男性之间实际上没有显著差异。
这项研究还表明,性腺类固醇与微生物群差异之间存在关联,绝经前妇女的类固醇生物合成和降解途径丰富,并且与血浆睾酮水平显著相关。
此外,微生物群可以预测人类粪便转移后人类和(经抗生素处理的)雄性小鼠的循环睾酮水平。
综上所述,这些结果表明,肠道微生物群组成的差异与性别以及不同激素状态有关。
既然知道了肠道微生物群与性激素之间存在相互作用,那么它们具体是通过什么机制影响的呢?
性激素和肠道微生物组之间相互作用机制
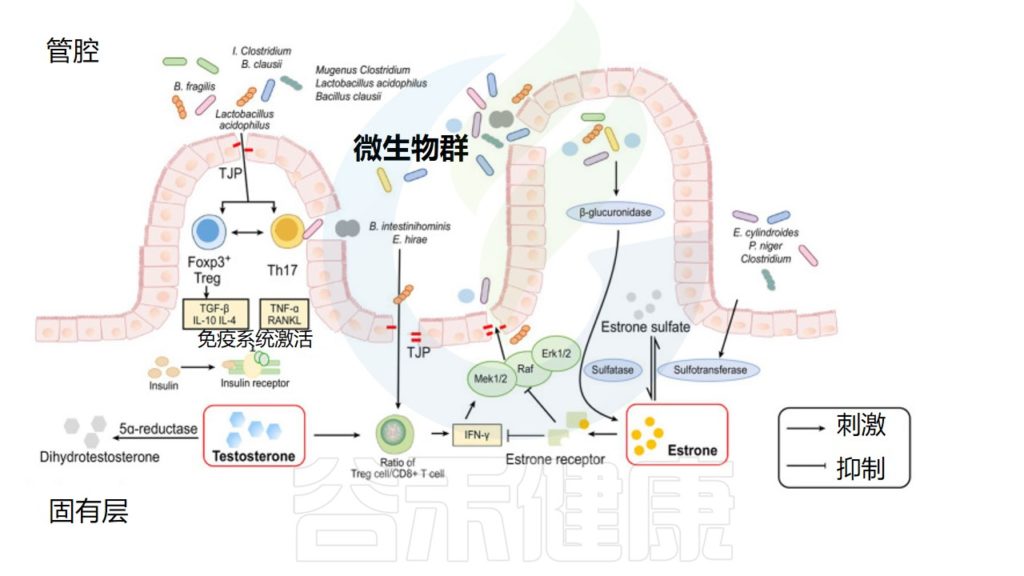
He S,et al.Front Microbiol.2021
1
微生物群通过改变胆汁酸影响性激素水平
最近有人提出,肠道微生物群的部分性别差异可能取决于胆汁酸,因为男性的胆汁酸库比女性大。在肝脏中由胆固醇合成后,它们被肠道微生物群代谢为次级胆汁酸,从而改变微生物群的结构并导致各种病理。
因此,肠道微生物群通过依赖于法尼醇X受体(FXR)的机制调节回肠中成纤维细胞生长因子15(FGF15)和肝脏中胆固醇7α-羟化酶(CYP7A1)的表达,从而调节胆汁酸的二次代谢,并抑制胆汁酸在肝脏中的合成。
FGF15抑制肝脏中CYP7A1的表达,CYP7A1是一种催化和调节胆汁酸合成第一步的酶。
胆汁酸同时也能影响微生物群的结构和丰度
已经观察到胆汁酸的减少导致细菌增殖,并且法尼醇X受体抑制细菌过度生长。
几项研究证实了胆汁酸、性激素和肠道菌群组成之间的关系。通过这种方式,给大鼠服用胆汁酸引起的微生物群变化类似于高脂肪饮食引起的变化,以牺牲拟杆菌属为代价增加厚壁菌门的水平。
此外,粪便微生物群(来自瘦供体)的移植产生了与瘦供体相似的肠道微生物群和胆汁酸库的变化,而小鼠的性腺切除术也改变了胆汁酸模式。
由于睾酮是由胆汁酸衍生的,并且如上所述,胆汁酸水平会被微生物群改变,因此微生物群可能间接影响睾酮水平的说法是成立的。
2
微生物群通过酶促激活性激素
共生微生物群落可以通过其酶的活性影响性激素水平。通过这种方式,术语“strobolome”被创造出来,定义为肠道微生物群中能够从其无活性葡萄糖醛酸苷中激活雌激素的一组基因,这主要归功于β-葡萄糖醛酸苷酶,它将雌激素解离为活性形式。这些活性雌激素进入血液并作用于雌激素受体α(ERα) 和雌激素受体β(ERβ)。
同样,最近的一项研究得出结论,肠道微生物群参与双氢睾酮 (DHT) 和睾酮的代谢和肠道去葡萄糖化,从而导致最高水平的雄激素。
肠道微生物群在性别差异中的另一种可能作用机制可以在羟基类固醇脱氢酶 (HSD) 中找到,这些酶参与类固醇激素的代谢并控制类固醇与其核受体的结合,使它们充当激活剂或抑制剂。
✦ 肠道微生物还通过植物雌激素影响代谢
除了三种主要形式的雌激素(胆固醇衍生的类固醇激素)、雌二醇(E2,在绝经前的非孕妇中占主导地位)、雌酮(E1,在绝经后占主导地位)和雌三醇(E3,在怀孕期间占主导地位),还有一种称为植物雌激素的植物化合物,其结构和功能与雌激素相似。
植物雌激素是天然存在于植物中的化合物,植物雌激素包括异黄酮,如染料木黄酮和黄豆苷原,它们主要存在于大豆中,并在被肠道微生物群代谢后被激活。从这个意义上说,肠道微生物群允许从黄豆苷元中获得O-Desmethylangolensin (ODMA) 和雌马酚,这两种物质都具有雌激素活性。
与雌激素类似,植物雌激素通过影响细胞信号传导引起生理效应,因为它们可以通过激活或抑制雌激素受体α或雌激素受体β来诱导或抑制雌激素作用,并且还可能引发表观遗传效应和细胞内信号级联反应。
植物雌激素可以调节内分泌系统改善病症
与此相关的是,几项人类研究表明,植物雌激素可以通过调节内分泌系统来改善各种病症,包括更年期症状,并且可以逆转代谢性内毒素血症的症状。
在这方面,植物雌激素代谢物雌马酚通过促进雌激素的尿液排泄和改变女性的血液水平,与降低女性激素相关疾病的风险有关,而不产生O-Desmethylangolensin与肥胖有关。
植物雌激素在饮食中被消耗,因为它们出现在水果、蔬菜、豆类和一些谷物中。事实上,饮食成分对肠道微生物群生态系统具有急性影响。通过促进更多样化和更稳定的微生物系统的发展,以植物为基础的饮食似乎对人类健康更有益。
!
药物和益生菌治疗的性别差异
已有一些关于药物治疗和微生物群性别差异的报道。
▷男女服用同一种抗炎药后微生物变化有差异
一项人体研究调查了23名健康男性和女性在非甾体抗炎药诱导的肠道屏障功能和微生物组方面的差异。使用 16S 核糖体 RNA 测序确定了十二指肠和粪便微生物群组成。健康女性的肠道通透性较低,十二指肠和粪便微生物多样性高于健康男性。
两性服用吲哚美辛后肠道通透性增加。然而,只有雌性在摄入吲哚美辛后粪便微生物多样性下降,包括普氏菌丰度增加。十二指肠微生物群组成没有表现出性别特异性变化。
▷益生菌给药存在性别差异
如今,益生菌因多种原因而流行,并且有人认为其效果可能因性别而异。
一项研究报告说,将5种乳杆菌菌株的益生菌混合物给予易患狼疮的小鼠可改善肾功能,并在雌性和阉割的雄性小鼠中显示出抗炎作用,但在性腺完整的雄性小鼠中则没有,这表明益生菌给药存在性别差异。
另一项研究报告了金枪鱼油和海藻油混合物对肠道微生物群的抗衰老作用存在性别差异。雄性小鼠在特定混合油比例下表现出比雌性小鼠更好的抗衰老效果。
肠道菌群失调引发的疾病在两性之间表现不同。研究最多的疾病主要是代谢性疾病。研究发现,性激素等多种因素影响这些疾病,它们密切相关,相互作用。
▸ 全球肥胖发病率越来越高
肥胖症的体重指数(BMI)为30kg/m2或更高,在全球各国成人和儿童中的流行率都在增加。这种病理学是复杂的遗传、社会经济和文化相互作用的结果,会导致严重的健康、经济和社会问题。
科学研究表明,一些代谢紊乱的发生与身体脂肪的分布有关,而且这种分布表现出性别二态性。脂肪倾向于堆积在男性的躯干和腹部周围,而女性的脂肪一般在臀部和大腿周围。
•腹部肥胖
腹部肥胖,尤其是内脏肥胖,会导致血糖和甘油三酯升高、高密度脂蛋白胆固醇降低和低密度脂蛋白 (LDL) 颗粒的增加,以及炎症标志物的增加 。
•臀部肥胖
相反,臀部脂肪与保护性脂质和葡萄糖水平以及降低代谢风险有关,似乎通过长期脂肪酸储存和有益的脂肪因子水平(与瘦素和脂联素水平呈正相关,与炎性细胞因子水平负相关)发挥其保护作用。
肥胖中的微生物多样性和性激素
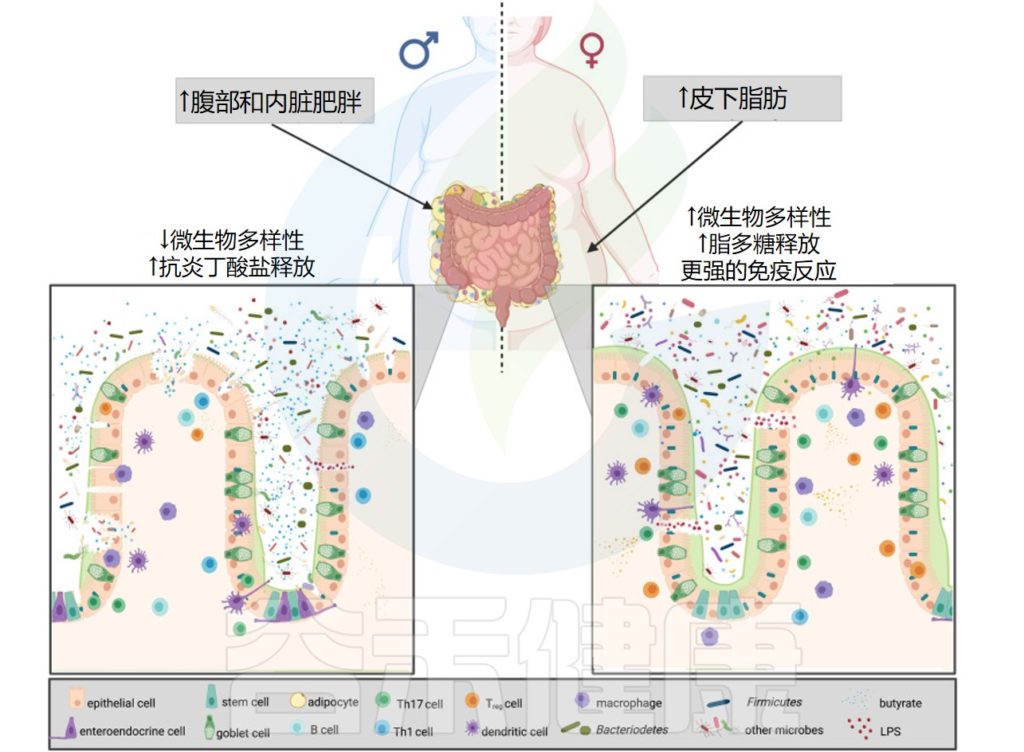
Brettle H,et al.Front Immunol.2022
在肥胖症中,拟杆菌门决定了疾病的严重程度。肥胖男性的物种丰富度较低,并且发现睾酮与厚壁菌门增加有关,因此会释放更多的抗炎丁酸盐。
另一方面,肥胖女性尽管微生物多样性更高,但雌二醇和拟杆菌增加,导致脂多糖释放更多,从而引发更强的免疫反应。
脂肪组织分布、性激素和肥胖的代谢紊乱
Brettle H,et al.Front Immunol.2022
与绝经前女性相比,男性和绝经后女性心血管风险增加,腹部/内脏肥胖和胰岛素皮下脂肪分布减少。男性和绝经后女性体内的脂肪组织比绝经前女性的脂肪组织更具促炎性。
√性激素调节身体脂肪分布
大量证据支持性类固醇调节身体脂肪分布的观点。在这方面,青春期激素的变化与不同性别之间的体重增加有关,这是由于男孩的瘦体重增加和女孩的脂肪量增加,以及男性和女性的脂肪分布。
此外,几项研究表明,一些基因参与了在体脂分布中观察到的性别二态性,以及性类固醇激素在这些基因调节中的潜在作用。
睾酮影响不同部位的脂肪堆积
在男性中,睾酮抑制腹内区域甘油三酯的摄取,并似乎促进它们在皮下区域的积累,同时导致儿茶酚胺刺激的皮下脂肪分解减少,而不是内脏脂肪分解。
这些过程似乎受到雄激素受体 (AR) 基因的影响,因为在雄激素受体敲除小鼠模型中,雄激素受体的缺失通过减少脂肪分解导致肥胖增加,尤其是晚期肥胖。
此外,caveolin-1蛋白(CAV1)似乎在脂肪堆积中起着重要作用,并且它受雌激素(雌二醇)和雄激素(双氢睾酮)的不同调节。
√性激素影响脂肪细胞功能
在细胞水平上,已经观察到性激素(雄激素和雌激素)对白色脂肪组织中脂肪细胞功能影响的差异,涉及脂肪细胞分化、脂肪分解、脂肪生成、胰岛素敏感性和脂肪因子产生/分泌等关键方面。
在这种情况下,睾酮和双氢睾酮调节小鼠间充质干细胞的分化,以雄激素依赖性方式分别促进和抑制它们向肌细胞和脂肪细胞的分化。
双氢睾酮增加脂肪分解
同样,在对人体细胞进行的体外研究中,双氢睾酮以雄激素受体依赖性方式抑制人间充质干细胞和人前脂肪细胞的脂肪形成分化,增加脂肪分解并减少脂质积累。
睾酮阻止脂肪细胞肥大和脂肪细胞生成
在阉割小鼠(雄性性腺功能减退症模型)中,脂肪量通过脂肪细胞肥大和脂肪生成增加,而当这些小鼠接受激素替代疗法时,睾酮阻止内脏和皮下脂肪量的扩张。
此外,致肥胖脂肪生成也通过抑制雄激素受体活性而升高。这项研究还显示了脂肪分布的不同调节,睾酮衍生的雌二醇和双氢睾酮分别阻止了内脏和皮下脂肪的增加。
√睾酮浓度较高利于减少内脏脂肪
睾酮浓度与向心性肥胖呈负相关,并且已发现睾酮治疗可减少具有雄激素缺乏症状和低血清睾酮水平的男性的内脏脂肪。
睾酮含量下降后脂肪更多的积累
此外,随着年龄的增长,睾丸激素的下降伴随着肥胖的增加,腹部脂肪的优先积累和内脏脂肪组织的更多积累。
还有报道称,内脏脂肪组织与生物可利用和游离睾酮呈负相关,皮下脂肪组织与性激素结合球蛋白 (SHBG) 呈负相关。
√雌二醇水平较低的女性易患向心性肥胖
在女性中,向心性肥胖与睾酮水平升高和雌二醇降低相关。更年期的荷尔蒙变化导致脂肪重新分布,与总脂肪和年龄无关,然而,一些研究表明,绝经后上半身脂肪的分布可能是由于衰老而不是绝经本身。
向心性肥胖——指患者体内脂肪沉积是以心脏、腹部为中心发展的一种肥胖类型。
最近发现,绝经前妇女的身体或躯干脂肪量与较低的总雌二醇和较高的游离雌二醇浓度有关,绝经前和绝经后妇女的总睾酮和计算游离睾酮浓度较高,性激素结合球蛋白和胰岛素样生长因子-I(IGF-I)浓度较低。
▼
代谢综合征是一种以腹部肥胖、胰岛素抵抗、高血压和高血脂为特征的病理状况,已遍布全球并导致2型糖尿病、冠心病和中风等疾病患病率上升。
性激素对肥胖、代谢综合征和2型糖尿病的影响

Santos-Marcos JA,et al.Biol Sex Differ.2023
√代谢综合征患者中男女的菌群差异
在代谢综合征患者中也观察到特定菌属的丰度差异。男性中韦荣氏球菌属(Veillonella)、Methanobrevibacter、Acidaminococcus、梭状芽孢杆菌(Clostridium)、罗氏菌属(Roseburia)和Faecalibacterium的丰度较高,而女性中嗜胆菌属(Bilophila)、瘤胃球菌属(Ruminococcus)和拟杆菌属(Bacteroides)的丰度更高。
√睾酮水平会影响代谢综合征
在睾酮对代谢综合征的影响中观察到的性别二态性似乎是雄激素受体依赖性的,并且已经提出了几种机制来解释睾酮水平与代谢综合征之间的关联。
男性中睾酮水平高不易患代谢综合征
在男性中,有证据表明睾酮与内脏肥胖、胰岛素抵抗和代谢综合症的发展呈负相关。沿着这些思路,在男性和啮齿动物中都报道了睾酮的雄激素受体介导的抗肥胖作用。
过量的睾酮可能导致女性胰岛素分泌受损
在女性中,据报道睾酮水平升高与胰岛素抵抗和葡萄糖耐受不良相关,因为它会降低全身葡萄糖的摄取。
关于睾酮对胰腺的作用,一项小鼠研究表明,雄激素受体调节男性胰岛β细胞生理,因此该受体的缺乏会降低葡萄糖刺激的胰岛素分泌并导致葡萄糖耐受不良。相反,有人提出,过量的睾酮可能通过雄激素受体依赖性机制导致女性胰岛β细胞功能障碍,胰岛素分泌受损。
√雌激素缺乏的女性代谢综合征患病率较高
雌激素水平似乎也影响代谢综合症的患病率。因此,大鼠卵巢切除术引起的雌激素耗竭导致大多数代谢综合征成分(脂质、葡萄糖、高密度脂蛋白和低密度脂蛋白)恶化,而在50岁以下的女性中,即经历更年期,其患病率增加。
卵巢切除的女性血糖和血压升高
此外,据报道,在接受过子宫切除术(通常伴随双侧卵巢切除术以预防后续卵巢癌)的女性中,血糖水平和高血压升高。
绝经导致性激素结合球蛋白水平下降,至少部分是由于雌激素减少,而睾酮水平在绝经期间没有改变 。从这个意义上说,绝经可以被认为是代谢综合征及其独立于年龄的所有独立组成部分的预测因素(风险因素)。
此外,已经描述了性激素结合球蛋白和代谢综合征之间的负相关,尤其是在绝经后妇女中。
▼
糖尿病包括一组疾病,根据其发展机制进行区分,这些疾病会降低调节血流中葡萄糖水平的能力,并导致长期高血糖症。
由于自身免疫和代谢过程,糖尿病有两种主要形式,即胰岛素依赖型糖尿病(1型糖尿病,T1D)和非胰岛素依赖型糖尿病(2型糖尿病,T2D)。
2型糖尿病的特征是胰岛B细胞产生的胰岛素不足和肝脏葡萄糖代谢受损,以及胰岛素抵抗,导致组织对胰岛素的反应性降低。
√糖尿病前期的症状表现出性别差异
空腹血糖受损 (IFG) 和葡萄糖耐量受损 (IGT) 作为2型糖尿病的前期阶段发生,表现出性别二态性,葡萄糖耐量受损在女性中更常见,而空腹血糖受损在男性中更常见。
有人提出,性激素可能是造成这种二态性的原因。事实上,更年期的雌激素治疗会降低空腹血糖并恶化葡萄糖耐量。
√男性的2型糖尿病发病率高于女性
此外,已证实男性的2型糖尿病发病率高于女性,这进一步支持了性激素参与这种病理学的发展。更年期2型糖尿病的风险增加,而更年期的激素治疗可能会延迟2型糖尿病的发作。
√睾酮水平影响男性2型糖尿病患病率
患有2型糖尿病的男性总睾酮和游离睾酮水平较低。与此相关的是,低水平的睾酮和性激素结合球蛋白与男性胰岛素抵抗和随后的2型糖尿病的发展有关。
低睾酮水平男性死亡率增加
此外,高水平性激素结合球蛋白和低水平睾酮的结合与2型糖尿病男性死亡率增加有关。其他研究表明,在患有2型糖尿病的男性中,低睾酮水平本身与死亡率增加相关,而睾酮替代疗法可能会提高这些男性的生存率。
同样,据报道,经过2年的睾酮治疗后,患有2型糖尿病的男性比例有所降低。此外,已发现前列腺癌的雄激素剥夺疗法会增加患糖尿病的风险。
患有2型糖尿病的男性睾酮水平往往较低
与上述一致,患有2型糖尿病的男性往往睾酮水平较低,而且他们中的大多数患有性腺功能减退症。事实上,许多研究已经证实,患有性腺功能减退症和低睾酮水平的肥胖2型糖尿病患者在接受睾酮替代疗法 (TRT) 后表现出改善的胰岛素抵抗和血糖控制。
√雌二醇水平高的男性患2型糖尿病的风险增加
关于女性荷尔蒙,雌二醇水平高的男性患2型糖尿病的风险增加,这种高雌二醇浓度与低性激素结合球蛋白浓度一起对男性患2型糖尿病的风险产生不利影响。
√女性睾酮水平较高可能易患2型糖尿病
与男性相反,女性的高睾酮水平与胰岛素抵抗和2型糖尿病相关。然而一项研究表明,中国女性性激素结合球蛋白值升高与患2型糖尿病的可能性降低相关,但雌二醇和睾酮水平与该族群中的2型糖尿病无关。
注:这些关于睾酮与2型糖尿病发病率之间关系的相互矛盾的结果可能是由于睾酮的测量,一些作者使用总睾酮而其他人使用游离睾酮,并且根据最近的一项研究,不同研究的分析方法可能不同。
此外,当性激素结合球蛋白浓度低于30nmol/L时,游离雄激素指数(FAI)不是游离睾酮的可靠指标,这将导致性激素结合球蛋白水平较低的女性可能出现研究错误。
因此,在女性中,总睾酮与2型糖尿病之间没有关联,尽管游离睾酮水平较高与2型糖尿病风险增加有关。
√雌二醇水平异常会增加2型糖尿病发病率
关于雌二醇,据报道患有2型糖尿病的绝经后女性的雌二醇水平高于健康女性。然而,基于早期初潮或绝经以及子宫切除术和卵巢切除术实践的大量证据表明,非生理性雌二醇水平(高于或低于正常值)可能是导致2型糖尿病发病率增加的原因。
月经初潮提前会增加2型糖尿病的风险
在这方面,月经初潮提前似乎会增加患2型糖尿病的风险。然而,一些研究表明,初潮早导致的2型糖尿病风险的部分原因可能是肥胖增加,因为早期月经初潮已被证明也与成年期体重指数的增加有关。
提前绝经和卵巢功能不全导致2型糖尿病风险增加
另一方面,提前绝经或卵巢功能不全会导致患2型糖尿病的风险增加。在接受双侧卵巢切除术的绝经后妇女中也观察到了类似的结果。
切除卵巢的女性2型糖尿病风险增加
最后,伴有双侧输卵管卵巢切除术(BSO)的子宫切除术显示2型糖尿病的风险高于子宫切除术本身。然而,其他研究表明子宫切除术与2型糖尿病风险增加有关,而双侧输卵管卵巢切除术本身或与子宫切除术一起并没有增加2型糖尿病的风险。
研究表明表明接受子宫切除术或卵巢切除术的女性患2型糖尿病的风险增加,但并未区分这两种情况是单独发生还是同时发生。
另一项研究表明,相对于完整女性,双侧卵巢切除的子宫切除女性的总睾酮和生物可利用睾酮水平较低,而保留卵巢的子宫切除女性的睾酮水平居中。
与此相关,绝经后妇女(子宫完整和子宫切除)的雌激素和孕激素激素疗法降低了糖尿病的发病率。
▼
肠易激综合征是一组持续或间歇发作,以腹痛、腹胀、排便习惯或大便性状改变为临床表现,胃肠道结构和生化异常的肠道功能紊乱性疾病。
√女性患肠易激综合征的比例更高
肠易激综合征 (IBS) 是一种具有性别差异的代表性疾病,通常在女性中的发生频率是男性的两倍。特别是,到三级中心就诊的患有严重症状的肠易激综合征患者的女性比例更高。
感染性结肠炎后肠易激综合征的风险增加,约10%的肠易激综合征患者在感染性结肠炎后开始出现症状。有趣的是,感染后肠易激综合征的发病率在女性中较高,这表明性别间肠道微生物群的差异可能在肠易激综合征的发病机制中发挥重要作用。
√肠易激综合征的症状存在性别差异
女性更有可能出现腹痛和便秘
肠易激综合征症状存在性别差异:具体而言,IBS-C在女性中占主导地位,而IBS-D在男性中占主导地位。
便秘型肠易激综合征(IBS-C)是肠易激综合征的一种亚型:至少25%的排便为硬粪或干球粪,且松散(糊状)粪或水样粪<25%。
腹泻型肠易激综合征(IBS-D):至少25%的排便为松散(糊状)粪或水样粪,且硬粪或干球粪<25%。
在对22项关于肠易激综合征症状性别差异的研究进行的系统回顾和荟萃分析中,发现女性比男性更可能出现腹痛和便秘相关症状。便秘型肠易激综合征在患有肠易激综合征的女性中的合并患病率为40%,而在患有肠易激综合征的男性中为21%。
男性出现腹泻症状更多
相比之下,患有肠易激综合征的男性比女性更有可能报告腹泻相关症状。腹泻型肠易激综合征(IBS-D)在患有肠易激综合征的女性中的合并患病率为 31%,而在男性中为50%。
√女性月经期间肠易激综合征症状加重
女性在月经期间报告更频繁和更严重的肠易激综合征症状,例如稀便、腹胀和腹痛加重,这些症状的增加可能与月经周期的变化有关。
在冰岛进行的一项基于人群的邮政研究表明,患有痛经的女性比没有痛经的女性更容易出现肠易激综合征症状。此外,更年期似乎与肠易激综合征症状的恶化有关。也就是说,女性在绝经后报告的腹痛比绝经前更严重。
此外,纤维肌痛、慢性疲劳综合征、慢性盆腔痛和偏头痛等经常与肠易激综合征重叠的慢性疼痛疾病的患病率在肠易激综合征女性中更高,这表明她们的症状与激素之间存在关联。
▼
甲状腺是重要的内分泌腺。甲状腺的主要功能是分泌含碘的甲状腺激素三碘甲状腺原氨酸(T3)和甲状腺素(T4)以及肽类激素降钙素,通过影响新陈代谢和组织发育来影响心血管和生殖疾病。
肠道微生物群在甲状腺疾病中起着至关重要的作用,包括桥本氏甲状腺炎 (HT) 和格雷夫斯氏病 (GD)。桥本氏甲状腺炎和格雷夫斯氏病分别是甲状腺功能减退症和甲状腺功能亢进症的主要原因。
√女性甲状腺疾病发病率高于男性
甲状腺疾病性别差异特别的显著,它的高发人群多为中年的女性,男性的患病率相对来说比较低,而之所以会这样是因为女性的内分泌活跃程度很高,没有一定的稳定性。
在平时压力大和情绪发生变化的情况下,会导致自身的免疫调节出现异常,致使甲状腺激素分泌过多,从而就会容易造成甲状腺疾病的发生。
√胆汁酸的组成可以反映甲状腺功能
胆汁酸是肠道微生物群的重要代谢产物,在甲状腺疾病中起着至关重要的作用。甲亢和甲减患者的血清胆汁酸谱不同,胆汁酸的组成可以反映甲状腺功能。甲状腺功能减退患者中最主要的胆汁酸是次级胆汁酸脱氧胆酸,而鹅去氧胆酸是甲状腺功能亢进患者中最主要的胆汁酸。
胆汁酸的组成和水平参与甲状腺激素的分泌,促甲状腺激素水平与亚临床甲减患者血清总胆汁酸呈负相关。
√短链脂肪酸与甲状腺激素相互作用影响女性情绪健康
短链脂肪酸是肠道菌群代谢的主要产物之一,短链脂肪酸与甲状腺激素三碘甲状腺原氨酸相互作用,并通过调节肠细胞基因转录来影响激素的分泌。例如,粪便短链脂肪酸酯可通过影响高脂血症和甲状腺疾病来影响女性的情绪健康。
▼
卵巢癌症的病因尚不清楚,可能与环境、生殖、行为和遗传因素有关。在这些因素中,雌激素水平的失调和雌激素的活性被认为是重要因素。
肠道微生物群可能通过影响雌激素水平参与卵巢的发育。发现17β-雌二醇治疗会改变卵巢癌小鼠模型的病理生理学,导致肿瘤提前发作、缩短总生存时间,并具有典型的乳头组织学特征。
√雌激素促进细胞粘附和迁移从而影响卵巢癌的病症
利用不同的卵巢癌症细胞系进行了一项体外研究,以确认雌激素受体在卵巢癌症病因中的需求。
他们的结果表明,雌激素治疗后,雌激素受体阳性细胞系的粘附和迁移能力增强,而雌激素受体阴性细胞系的粘附和移动能力没有显著变化。
这些结果表明,雌激素的作用与雌激素受体的表达直接相关,雌激素促进细胞粘附和迁移,从而影响卵巢癌症的转移和定植。
√微生物通过影响活性雌激素含量影响卵巢癌
如上所述,具有 β-葡萄糖醛酸酶活性的细菌可以介导雌激素的去结合,从而影响循环中活性雌激素的数量。结合雌激素水平和活性在卵巢癌病因学中所起的作用,我们可以推断肠道微生物组可能通过影响活性雌激素的量来促进雌激素驱动的疾病(如卵巢癌)的发展。
▼
绝经后骨质疏松症 (PMOP) 是一种由雌激素缺乏引起的骨质疏松症,会导致绝经后妇女骨折频率增加。
√雌激素缺乏易引起骨质疏松
目前的研究表明,肠道微生物群与骨重塑以及骨代谢疾病之间存在潜在的密切关系。
研究人员发现,在无菌小鼠中,性类固醇缺乏不能诱导破骨细胞因子表达增加、骨吸收激活,这表明肠道微生物组在性类固醇缺乏导致的骨质疏松中至关重要。
他们进一步证明,用益生菌鼠李糖乳杆菌或市售益生菌补充剂每周两次治疗性类固醇缺乏小鼠可以避免骨质疏松。这种情况的发生可能是由于肠道通透性的降低、肠道和骨髓炎症的抑制。
相比之下,补充非益生菌大肠杆菌菌株或突变鼠李糖乳杆菌并没有显示出对骨丢失的保护作用。
上述结果表明,肠道微生物群失调可能导致肠道通透性增加,并触发重要炎症通路的激活,从而导致性类固醇缺乏小鼠的骨质流失。
这些结果将雌激素缺乏引起的骨质疏松症与肠道微生物多样性、肠道通透性和炎症联系起来。
▼
√高雄激素可能导致多囊卵巢综合征
多囊卵巢综合征是育龄妇女常见的内分泌疾病,其病因可能与高雄激素血症、胰岛素抵抗和神经内分泌功能障碍有关。
一些研究关注多囊卵巢综合征患者的肠道微生物组,这些研究发现肠道微生物组与多囊卵巢综合征之间存在某种关联。然而,与多囊卵巢综合征相关的微生物的确切机制尚未确定。
√多囊卵巢综合征患者肠道微生物多样性较低
一些研究人员发现,患有多囊卵巢综合征的患者肠道微生物组的多样性明显低于健康对照组。在多囊卵巢综合征患者的肠道微生物组中,普通拟杆菌(Bacteroides vulgatus)显著升高,脱氧胆酸和熊去氧胆酸水平降低。
通过比较多囊卵巢综合征女性的粪便微生物组、肠上皮通透性和炎症状态,一些研究提出了一个假设,即肠上皮高通透性导致的毒血症与炎症、胰岛素抵抗和高雄激素血症有关。不过还需要更深入的研究来阐明内毒素血症的具体机制。
√调节肠道微生物有助于改善多囊卵巢综合征
菊粉和二甲双胍对多囊卵巢综合征的缓解作用与抗炎和肠道微生物群的调节有关,这可能有助于多囊卵巢综合症的潜在临床治疗。
环丙孕酮Diane-35(雌激素和孕激素)和益生菌可以帮助重建肠道微生物群的多样性,减少肠道菌群紊乱可以改善多囊卵巢综合征样大鼠的生殖功能。
还需要进一步的研究来确定改善肠道微生物组是否可以作为多囊卵巢综合征的有效治疗方法。
小结
根据性激素和肠道微生物组之间的相互作用,性激素相关疾病的可能治疗策略的显示在下图:
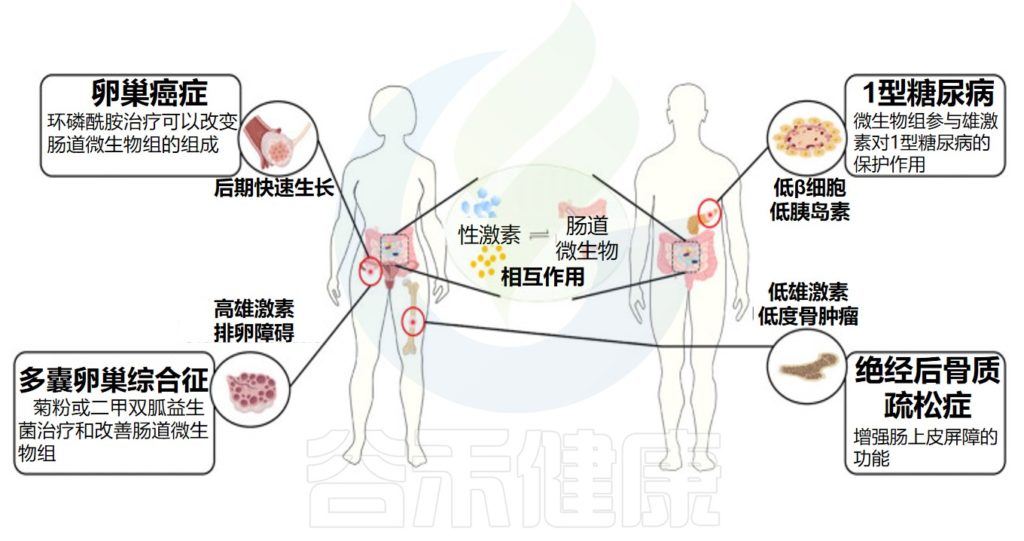
He S,et al.Front Microbiol.2021
在对性激素相关疾病治疗的过程中发现了肠道微生物群的变化,未来通过对肠道微生物的检测或许有助于判断性激素相关疾病的治疗效果。
性激素,特别是雌激素和睾酮,与肠道微生物群的结构和组成有关,性激素与肠道微生物群的相互作用对疾病的发展和性别差异具有重要影响。
肠道微生物群的特定变化可能有助于缓解激素紊乱相关的疾病,因此肠道微生物群被认为是一个重要治疗靶点。
在制定治疗策略时,确定导致疾病发病率中性别二态性的机制具有特殊的重要性,因为肠道微生物群的组成取决于与性激素的相互作用以及其他因素。治疗与这些疾病相关的肠道微生物群失调的疗法可能具有性别特异性效应。
总之,肠道微生物群与激素相互作用在疾病中的影响已经成为了一个备受关注的研究领域。肠道微生物群在激素变化及其带来的健康风险的重要作用及其检测的意义也被越来越多的人所认识。未来,随着相关研究的不断深入,肠道微生物群与激素相互作用的机制和应用价值也将会进一步得到探索和发展。
主要参考文献
Santos-Marcos JA, Mora-Ortiz M, Tena-Sempere M, Lopez-Miranda J, Camargo A. Interaction between gut microbiota and sex hormones and their relation to sexual dimorphism in metabolic diseases. Biol Sex Differ. 2023 Feb 7;14(1):4.
Li D, Sun T, Tong Y, Le J, Yao Q, Tao J, Liu H, Jiao W, Mei Y, Chen J, Liu Z, Wang G, Li Y. Gut-microbiome-expressed 3β-hydroxysteroid dehydrogenase degrades estradiol and is linked to depression in premenopausal females. Cell Metab. 2023 Apr 4;35(4):685-694.e5.
Brettle H, Tran V, Drummond GR, Franks AE, Petrovski S, Vinh A, Jelinic M. Sex hormones, intestinal inflammation, and the gut microbiome: Major influencers of the sexual dimorphisms in obesity. Front Immunol. 2022 Sep 27;13:971048.
Kautzky-Willer A, Harreiter J, Pacini G. Sex and Gender Differences in Risk, Pathophysiology and Complications of Type 2 Diabetes Mellitus. Endocr Rev. 2016 Jun;37(3):278-316.
Saklayen MG. The global epidemic of the metabolic syndrome. Curr Hypertens Rep. 2018;20(2):12.
Paschou SA, Papanas N. Type 2 diabetes mellitus and menopausal hormone therapy: an update. Diabetes Ther. 2019;10(6):2313–2320.
Valeri F, Endres K. How biological sex of the host shapes its gut microbiota. Front Neuroendocrinol. 2021 Apr;61:100912.
Kim YS, Unno T, Kim BY, Park MS. Sex Differences in Gut Microbiota. World J Mens Health. 2020 Jan;38(1):48-60.
Yoon K, Kim N. Roles of Sex Hormones and Gender in the Gut Microbiota. J Neurogastroenterol Motil. 2021 Jul 30;27(3):314-325.
He S, Li H, Yu Z, Zhang F, Liang S, Liu H, Chen H, Lü M. The Gut Microbiome and Sex Hormone-Related Diseases. Front Microbiol. 2021 Sep 28;12:711137.

谷禾健康

音乐对身心的影响
近年来,环境和动物生理及心理的相关研究越来越多。环境因素的丰富性和多样性是改善动物生理和心理状态的重要研究参数。
环境因素指的是正常环境,在这种环境中,动物通过获得环境激励以做出有益的增强,使它们能够正常表达其行为和心理活动,从而改善健康状况和生长性能。
★ 音乐对人体健康具有益处

音乐这种声波刺激,影响着动物的生理和心理。以前的研究表明,伴随音乐喂养动物会影响它们的生长性能和动物生产。音乐的节奏鼓励肌肉活动并激发机体的活力。当音乐的节奏接近机体的心律时,会刺激机体分泌调节荷尔蒙。
美国开展的一项音乐疗法研究表明,它可以缓解情绪、改善消化、平衡心理状态并促进疾病康复。
★ 音乐可以辅助调节肠道菌群
中国在先秦时期就将五行音乐疗法(FEMT)引入医学领域。中国的五行音乐疗法有着悠久的历史和完整的体系。科学家观察到五行音乐疗法可以缓解焦虑症状、改善空间认知、调节肠道菌群和辅助药物治疗。
五行音乐疗法——中医理论描述了五种音乐音调、五种器官和五种元素之间的相互联系,五音分属五行,通肝、心、肺、脾、肾五脏。用角、徵、宫、商、羽五种不同的音调的音乐来治疗相对应的疾病。
虽然目前音乐疗法治疗疾病存在一定的局限性,但音乐疗法已成为临床常用的辅助治疗手段。
然而,关于喂食过程中听音乐对肠道微生物影响的报道较少。本文通过最新的研究评估喂养时伴随音乐对小鼠肠道微生物和生长性能的影响,为音乐治疗提供理论支持。
•实验选用生长状况良好、健康的小鼠
在整个研究过程中,没有动物患抑郁症、疾病或死亡,尸检和电子显微镜成像均未显示宏观和微观病理。
整个实验期间的小鼠体重
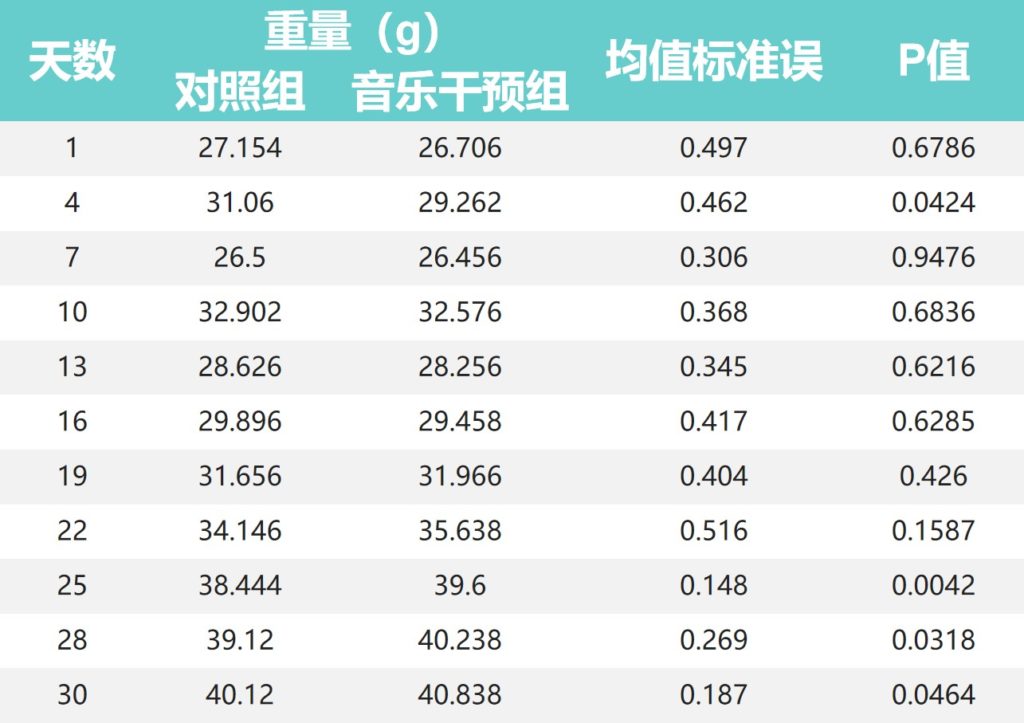
•对照组小鼠体重在前16天高于音乐干预组
对照组小鼠体重在30天喂养期的最初16天高于音乐干预组,第4天两组差异显著(P<0.05)。
•19天后音乐干预组体重开始高于对照组
然而,在第19天后,两组之间的小鼠体重发生了翻天覆地的变化,音乐干预组的体重在第19-30天高于对照组。
其中,第25天(38.444:39.600)、第28天(39.120:40.238)和第30天(40.120:40.838)组间差异有统计学意义(P<0.01;P<0.05;P < 0.05)。
•音乐干预组小鼠情绪和状态更活跃
值得注意的是,在喂食期间,我们发现音乐干预后的小鼠在情绪状况和活动状态方面比对照小鼠更活跃。
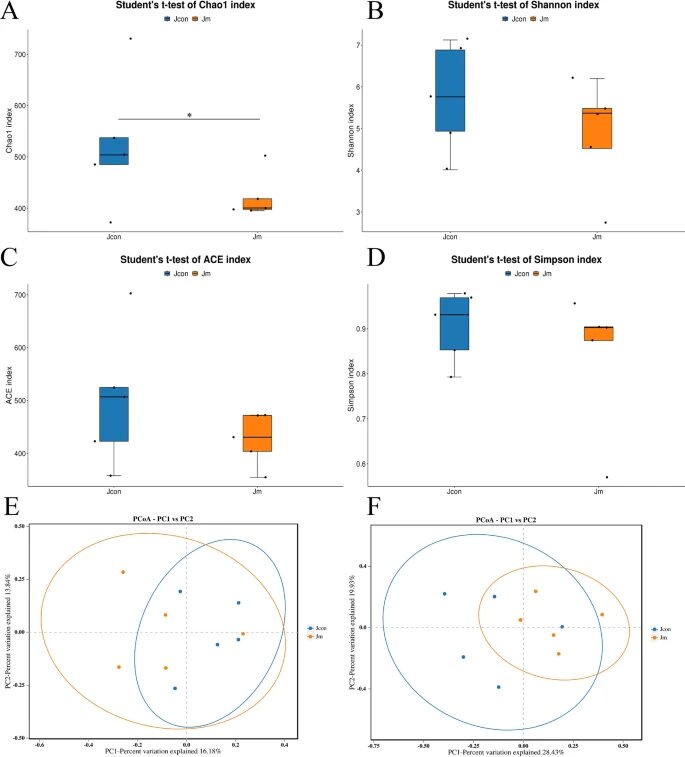
匹配了获得的合格序列,并进行了α-多样性分析,以评估对照组和音乐干预组之间肠道菌群多样性和丰度的差异。
✦伴随音乐的喂养降低了肠道菌群丰度
结果表明,对照组的微生物多样性和丰度高于音乐干预组,两组间chao1指数差异显著(P<0.05)。其他三个指数(Shannon指数、ACE指数和Simpson指数)没有显示出统计差异。
注:chao1指数——又称Chao1丰富度指数或Chao1多样性指数,是一种用于衡量群落物种多样性的指标。
综上所述,可以看出伴随音乐的喂养过程降低了肠道菌群的丰度。
✦β多样性较低
利用QIIME软件计算β多样性分析,比较样本间物种多样性的相似性。
PCoA分析基于未加权的二进制 jaccard(P>0.05)和加权的 bray curtis(P>0.05)显示两组之间没有统计学差异,表明两组之间具有高相似性和低物种多样性。
α和β多样性分析
▼
通过使用 QIIME2 软件对来自不同物种的微生物类群进行分类,评估了门和属水平上优势样本的相对比例。
✦两组中都以厚壁菌和变形菌为主
结果表明,对照组中门水平的优势菌为厚壁菌门(Firmicutes)占42.19%,变形菌门(Proteobacteria)占19.57%,蓝藻(Cyanobacteria)占13.05%和拟杆菌门(Bacteroidetes)占9.85%,占总肠道菌群的84.66%。
在音乐干预组中,门水平的优势细菌为厚壁菌门(Firmicutes)占61.42%,变形菌门(Proteobacteria)占16.00%、蓝藻占7.07%和拟杆菌门(Bacteroidetes)占4.46%,占微生物总数的88.94%。两组的微生物组成(属水平)均显示乳杆菌(Lactobacillus)为优势群落。
此外,uncultured_bacterium_f_Enterobacteriaceae(3.97%比5.83%)和uncultured_bacterium_f_Muribaculaceae(3.47%比1.62%)是两组中的优势菌株。
小结
从以上结果可以得出结论,在小鼠喂食过程中进行音乐干预可以改变肠道微生物的组成。
热图通过色带和接近程度表明多个样品的肠道微生物种群的异同。
结果表明,无论细菌门水平还是在属水平,两组样品之间的微生物丰度相似性较低,但组内样品之间的差异较小,微生物丰度相似。
小鼠肠道微生物组成分布和热图
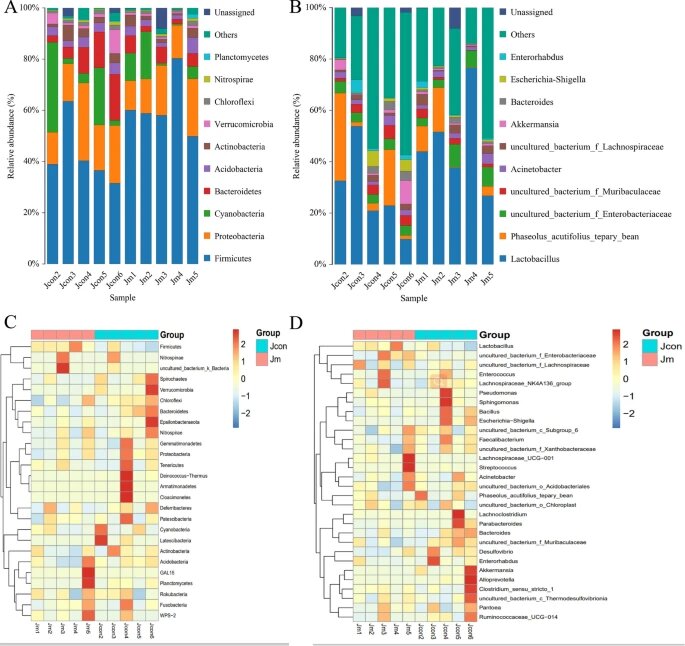
注:(A) 门级的优势细菌种类和丰度;(B) 属级的优势细菌种类和丰度;(C) 门级样品间微生物相似性热图;(D) 属级样品间微生物相似性热图。
▼
✦两组之间厚壁菌丰度存在显著差异
使用 Metastats 分析,研究了两组微生物的门和属水平组成的统计学差异。结果显示厚壁菌门的相对丰度存在显著差异(对照组0.422 ± 0.056与音乐干预组0.614 ± 0051,P < 0.05)。
Metastats分析——组间群落显著性差异分析,用于寻找组间差异物种
✦对照组丰度高于音乐干预组的菌群
在属水平上发现了11个具有统计学意义的分类群,而对照组在以下菌种丰度高于音乐干预组
uncultured_bacterium_o_Microtrichales
(P<0.0001);uncultured_bacterium_f_Micromonosporaceae
(P<0.05);
Pseudolabrys (P<0.05);
Methylobacterium (P<0.05);
uncultured_bacterium_f_Muribaculaceae
(P<0.05);
Ruminococcaceae_UCG-005 (P<0.05) 。
✦音乐干预组丰度高于对照组的菌群
与此相反,以下菌种丰度音乐干预组高于对照组:
uncultured_bacterium_f_Atopobiaceae(P<0.01);Ileibacterium(P<0.01);Lachnospiraceae_FCS020_group(P<0.05);Serratia(P<0.05);
Dietzia (P<0.05)。
门和属水平肠道细菌丰度的统计差异
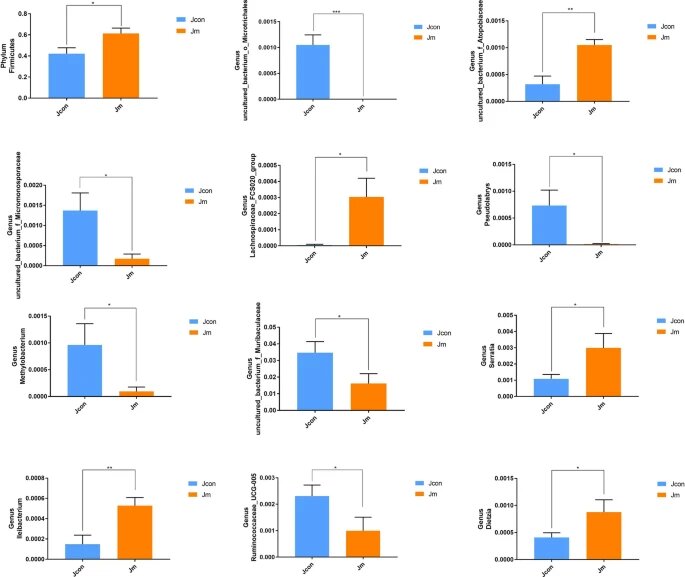
注:所有数据均表示平均值±SD(*P<0.05;**P<0.01;***P<0.001)
▼
考虑到 Metastats 分析对两组间细菌相对丰度差异的局限性,进行了 LEfSe 分析以寻找对照组和音乐干预组之间具有统计学差异的生物标志物。
LEfSe分析是一种将非参数的Kruskal-Wallis以及Wilcoxon秩和检验,与线性判别分析效应量相结合的分析手段, 能够在不同组间寻找具有统计学差异的Biomarker,其要求组内样本数≥3。
结果表明,除了上述显著差异的细菌外,还回收了15个具有统计学差异的生物标志物,对照组中最主要的群体是疣微菌门(Verrucomicrobia)、δ-变形菌(Deltaproteobacteria)和酸微菌(Acidimicrobiia),而Lleibacterium和大洋芽孢杆菌(Oceanobacillus)在音乐干预组中显著表达。
LEfSe分析和LDA评分揭示了细菌丰度的显著差异
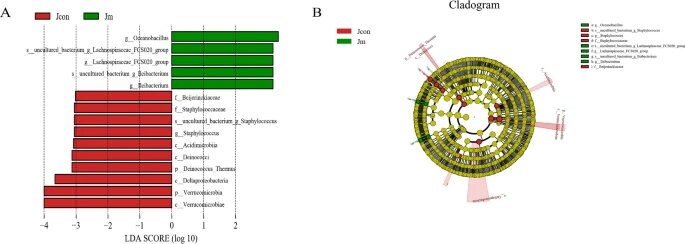
注:(A) LDA分数>3.0被认为具有统计学意义。(B) 分支图描绘了与对照组和音乐干预组相关的肠道微生物群的系统发育分布。
▼
在生态学领域,相关性通常用于构建可用于分析物种群落数据(共现模式)或组合多个数据集进行分析的网络模型。以python为基础,使用前50个属来创建基于网络的关联,该关联表示78个节点、237个边和6个社区。
✦乳杆菌是两组中最丰富的菌
我们发现,在基于网络的相关性分析中,乳杆菌(Lactobacillus)是两组中最丰富的细菌。
此外,音乐干预后小鼠(Jm组)体内乳杆菌丰度高于对照组(47.34%比28.03%),而Phaseolus_acutifolius_tepary_bean丰度低于对照组(6.24% 比 12.36%)。
✦乳杆菌与其他菌属的关联性
乳杆菌与Parabacteroides(0.6364)、Faecalibacterium(0.7455)、幽门螺杆菌(0.7720)、RB41(0.6485)呈负相关。
Phaseolus_acutifolius_tepary_bean与Candidatus_Koribacter(0.7173)和Breznakia(0.7195)呈正相关,与泛菌属(0.66060)、肠球菌属(0.6809)和Ruminococcacee_UCG-014(0.6606)呈负相关。
物种间相关性分析
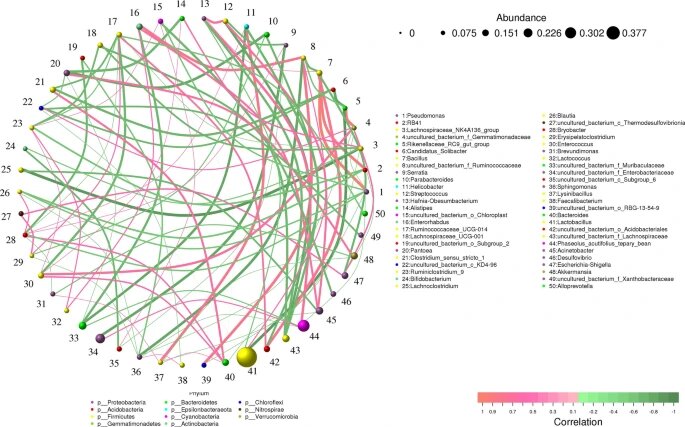
线条描绘了两个物种之间的相关性,线条粗细描绘了相关性的强度。根据线条的颜色,橙色表示正相关,绿色表示负相关。
已发现音乐对身体有积极的影响,引起协调的共振,促进器官节奏的和谐,以及一系列内分泌转变。
▼
✦提高动物的生长性能
先前的研究表明,音乐和检查声音的混合会显著提高小猪生长性能(同样的饲料量下长了更多的肉)。然而,有些音乐(例如重金属,频率<20赫兹,95–105分贝)具有相反的效果。
注:这项研究是在 El Condor Pasa 音乐上进行的,该音乐风格自然且没有过度分层。
在为期30天的喂养实验(有音乐干预)中,发现小鼠的体重在第25天时明显高于对照组。此外还显示,在音乐干预后,动物消耗了更多的食物,体重明显增加。
✦促进对营养的吸收
一些研究发现,音乐可以促进免疫系统和肠道微生物对营养的吸收,同时还可以减轻噪音的负面影响。总的来说,表明音乐可以改善体重,促进小鼠的生长和身体发育。
▼
✦增强肠道菌群的活性
先前的研究表明,音乐疗法可以减少抑郁症,它还会影响消化系统的协调共振、生长激素的分泌并增强肠道菌群的活性和多样性。
肠道微生物群是动物肠道中的微生物群落,在微生物学、医学和遗传学等肠道微生物群相关领域中观察到越来越多的研究趋势。
小鼠胃肠道α-多样性和PCoA分析表明,与对照组相比,音乐干预组肠道微生物发生了变化。有趣的是,整个多样性分析揭示了音乐组中组内样本的高度聚集和组内相似性的显著增强。
✦有助于肠道菌群的相对稳定
我们发现喂食期间的音乐干预从统计学上增加了厚壁菌和乳杆菌的丰富度,同时成倍减少了蓝藻、拟杆菌、肠道微生物中的Phaseolus_acutifolius_tepary_bean和uncultured_bacterium_f_Muribaculaceae。
厚壁菌门是数量最多的细菌,大多数为革兰氏阳性菌,呈球形或杆状,厚壁菌门的许多成员是有益菌,如乳杆菌、芽孢杆菌、双歧杆菌、丁酸梭菌等。
表明肠道微生物多样性的减少与特定细菌的减少或个别菌株的过度生长有关。这表明喂养期间的音乐干预可以减少离散的微生物群并有助于相对稳定性。
✦肠道微生物的稳态有助于防止其他疾病
肠道微生物群的稳态被发现是外部病原体入侵和定植的重要障碍,肠道微生物群的改变可能与多种疾病有关。
除了胃肠道疾病和代谢疾病,肠道菌群还与多种全身性疾病有关,例如神经系统疾病、呼吸系统疾病、心血管疾病和肿瘤疾病。
研究表明,乳杆菌等微生物可以产生乙酸盐、乳酸和抗菌物质,可以防止病原体干扰健康。 此外,它们还有助于维持肠道微生态平衡,预防和抑制肿瘤的发生,增强动物免疫力,促进消化,合成氨基酸和维生素。
总体而言,本研究描述了小鼠在进食期间听音乐情况下肠道微生物群的变化。
结果表明,音乐干预后肠道微生物群发生了显著变化,其特征是肠道细菌多样性下降和小鼠肠道微生物群组成发生变化。此外,在伴随音乐喂养小鼠期间,有益菌的数量增加,而致病菌或条件致病菌减少。
这些结果有助于理解音乐与肠道微生物群之间的关系,以及肠道微生物群会根据不同的喂养环境发生变化。本研究也为音乐疗法改善动物生长环境提高动物生理及心理状态提供了理论依据。

关于肠道菌群干预的相关措施有很多,比如说饮食、睡眠、运动、益生菌、粪菌移植、天然产物、抗生素等,这些在我们之前写过的相关主题的文章都有涉及;而关于音乐对肠道菌群的干预的研究较少,本文对此做一个详细介绍,为音乐疗法的有效性提供了理论依据。
音乐是我们日常生活中轻而易举就能接触到的,可以说是一种非常便捷的治疗方式,因为它不需要使用任何药物或器械,无需去医院等特殊的场所。通过本文的研究可以窥见音乐疗法对于干预人类肠道菌群的潜力。
对于研究人员而言,未来可以更深入了解音乐疗法的作用机制,通过研究不同类型的音乐、不同的音乐治疗对象,探索更多有效的个性化的音乐调节肠道菌群的方案,从而更好地应用于临床实践中,让大众有机会感受音乐疗法带来的不可思议的力量。
参考文献
Niu J, Xu H, Zeng G, Wang P, Raciheon B, Nawaz S, Zeng Z, Zhao J. Music-based interventions in the feeding environment on the gut microbiota of mice. Sci Rep. 2023 Apr 18;13(1):6313.
谷禾健康
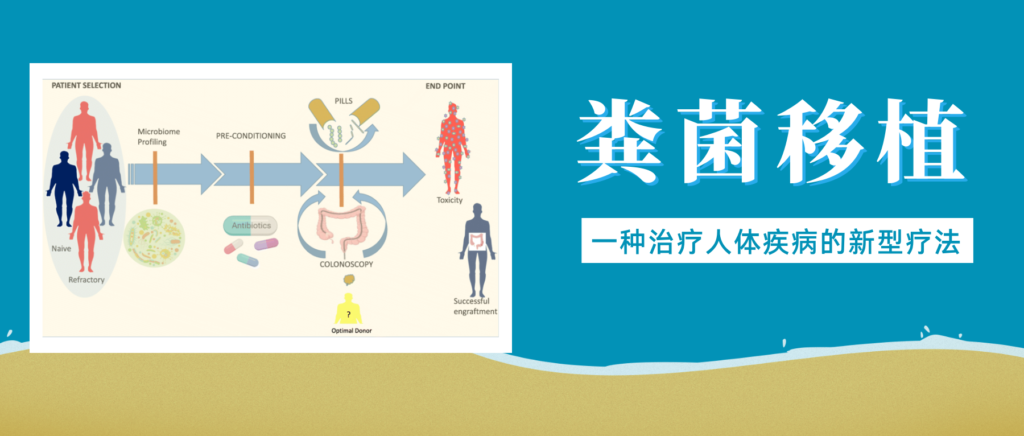
粪菌移植是一项近年来备受关注的医疗技术,它涉及将健康捐赠者的粪便物质转移至患有疾病或障碍患者的胃肠道。

简单来说就是选择健康合适的人粪便,通过科学方法提取出有用的微生物,去除有害与无用的部分,然后制成制剂,给病人口服或者直接注入肠道,目的是恢复健康的肠道微生物群并改善整体健康状况。
粪菌移植的历史悠久
粪菌移植在传统中医中已有数百年的应用,粪菌移植的第一个记录可以追溯到四世纪的中国,当时人类粪便被称为黄汤,用于治疗严重腹泻的患者。
15到16世纪的中国明朝,有关于将新鲜或发酵的粪便悬浮液用于治疗腹泻、便秘和腹痛等胃肠道疾病的记载。
但直到20世纪50年代晚期,它才首次用于现代医学,以治疗由抗生素耐药菌引起的严重腹泻。
在20世纪80年代和90年代,粪菌移植主要用于治疗反复发作的艰难梭菌感染。然而,随着肠道微生物群的发现及其在健康和疾病中的作用,粪菌移植开始被探索作为治疗其他疾病的潜在方法,包括炎症性肠病、肠易激综合征甚至神经系统疾病等。

粪菌移植在未来拥有巨大前景
近年来,粪菌移植受到医学界和公众越来越多的关注,许多临床试验和研究正在进行,以评估其疗效和安全性。
粪菌移植的疗效已经在多种肠道疾病中得到证实,如艰难梭菌感染、肠炎、炎症性肠病等。粪菌移植的出现,为肠道微生物失调带来了新的治疗思路和方法,也为人类健康提供了新的希望。
随着我们对肠道微生物群研究的不断发展,粪菌移植很可能成为治疗其他疾病越来越重要的工具。谷禾在本文中将为大家介绍粪菌移植。
粪便微生物群移植 (粪菌移植) 是一种医疗程序,涉及将粪便从健康供体转移到受体的胃肠道中。已发现粪菌移植可有效治疗各种胃肠道疾病,包括艰难梭菌感染、炎症性肠病和肠易激综合征。
来自大量临床试验的数据表明,粪菌移植也可能对许多其他疾病具有治疗潜力,从胃肠道到肝脏疾病,神经系统疾病、自身免疫性疾病、皮肤疾病、心血管疾病、脑部疾病、肥胖和代谢综合征。
根据这些相关研究,谷禾带大家了解粪菌移植在治疗人体疾病中的作用。
人类疾病中的粪菌移植
Biazzo M,et al.J Clin Med.2022
▼
粪菌移植主要用于治疗反复发作的艰难梭菌感染(CDI),这是一种细菌感染,会引起严重的腹泻和结肠炎症。
√粪菌移植对治疗艰难梭菌感染特别有效
研究表明,粪菌移植在治疗复发性艰难梭菌感染方面特别有效,已发现粪菌移植在治疗艰难梭菌感染方面的成功率超过90%,而且几乎没有副作用。
▼
粪菌移植已被证明是治疗多种胃肠道疾病的有效方法,包括炎症性肠病(IBD)、肠易激综合征(IBS)。
炎症性肠病是一种复杂的炎症和慢性疾病,其特征是免疫失调,最终导致消化道免疫介导的损伤,包括溃疡性结肠炎(UC)和克罗恩病(CD)。肠道微生物群失调被认为是炎症性肠病发展中的关键调节事件,因此粪菌移植代表了一种可能的治疗策略。
√缓解溃疡性结肠炎患者症状
最近一项随机对照试验的荟萃分析发现,粪菌移植可有效诱导溃疡性结肠炎患者症状缓解,缓解率为36%,而对照组仅为9%。对核糖体 16S RNA 的分析表明,粪菌移植后微生物多样性增加并持续存在。
对活动期溃疡性结肠炎的儿科患者(4至17岁)进行了随机临床试验。粪菌移植组中92%的患者在第6周实现了小儿溃疡性结肠炎活动指数的改善(与安慰剂组的50%相比)。
√短期内改善了克罗恩病患者症状
评估了25名患有腹腔内炎症性肿块的克罗恩病患者使用多种新鲜粪菌移植(初始粪菌移植随后每三个月重复粪菌移植)的疗效和安全性。
超过一半的患者在第一次粪菌移植后三个月表现出临床反应的缓解,这一比例在12个月和18个月时有所下降。
注:这表明尽管粪菌移植在短期内缓解了临床症状,但未能产生持久的临床效果。
√对肠易激综合征有积极影响
六项不同的临床试验发现粪菌移植对肠易激综合征症状有积极影响:
(1) 在一项只有10名患者入组的小型临床试验中,在粪菌移植后4周,6 名患者有所改善;有趣的是,作者发现报告改善的患者接受了来自双歧杆菌含量高于无效供体的粪菌移植,这表明富含双歧杆菌的粪便供体可能是粪菌移植成功的预测因素。
(2)观察到65%的患者在通过结肠镜检查接受粪菌移植治疗三个月后症状缓解以及粪菌移植之后微生物特征向供体特征的转变,包括增加的α和β多样性。
(3)报告了腹泻型肠易激综合征患者的症状和生活质量的改善,这些患者通过胃镜接受新鲜粪便,并且(在连续分析中)增加了短链脂肪酸。
(4)报告显示,在接受来自健康供体(同种异体移植)或来自他们自己(自体移植)的粪便材料的患者中,肠易激综合征症状出现短暂改善;接受同种异体移植的患者抑郁评分有所下降。
(5)使用仅从一名健康、特征良好的供体获得的粪便样本通过胃镜粪菌移植治疗肠易激综合征患者,发现了疲劳和生活质量的改善以及细菌微生物群概况的变化、粪便短链脂肪酸的变化。
(6)在最近的一项临床试验中,招募了难治性肠易激综合征患者,主要是腹胀,并通过鼻空肠给药接受一次粪菌移植治疗;粪菌移植一年后,56%的患者报告肠易激综合征症状和生活质量有所改善。
▼
复发性肝性脑病是肝硬化的并发症(与饮酒无关),由肝功能衰竭和肠-肝-脑轴受损引起,可导致意识障碍和昏迷 。
√粪菌移植改善肝性脑病患者认知和微生态失调
粪菌移植最近被研究为一种可能的新治疗工具。第一项试验描述,通过灌肠递送的单一粪菌移植在短期内减少了肝性脑病患者的住院时间,改善了他们的认知和微生态失调。
粪菌移植治疗后一年内,与对照组相比,降低了住院率并改善了认知功能。
√粪菌移植展现出肝病的治疗潜力
广泛的研究支持粪菌移植作为一种控制肝病的疗法。粪菌移植改善了高脂肪饮食引起的肝损伤和脂质代谢,同时增加了小鼠肠道微生物群的多样性。来自对酒精性肝病有抵抗力的供体小鼠的粪菌移植可以预防酒精性肝损伤。
此外,粪菌移植已经用于慢性肝病患者。最近一项针对严重酒精性肝炎患者的初步研究表明,粪菌移植与存活率提高和腹水消退有关。
▼
多项研究探索了粪菌移植在治疗神经系统疾病方面的潜在益处,例如帕金森病、多发性硬化症和自闭症谱系障碍。
√改善帕金森病患者症状
发表在《神经病学》杂志上的一项研究发现,粪菌移植改善了帕金森病患者的症状,如便秘和睡眠障碍。
在帕金森病小鼠模型中,来自正常小鼠的粪菌移植可以减少黑质的病理特征并减轻身体损伤。
√改善多发性硬化症患者症状
发表在《神经炎症杂志》上的另一项研究发现,粪菌移植改善了多发性硬化症患者的症状,例如疲劳和抑郁。
√影响抑郁症、阿尔兹海默症状态
此外,来自重度抑郁症患者的粪菌移植在正常小鼠中诱导了抑郁症的行为/生理特征。
最后,在阿尔茨海默病小鼠模型中,源自正常对照小鼠的粪菌移植改善了认知功能。
▼
除神经系统疾病外,粪菌移植在治疗其他非胃肠道疾病(如肥胖、糖尿病和代谢综合征)方面也显示出前景。
√改善代谢综合征患者胰岛素敏感性
几项临床试验研究了粪菌移植对代谢综合征患者的的影响,显示出改善:报告说,患有代谢综合征的男性参与者接受了来自健康人群的肠道微生物群输注六周后胰岛素敏感性增加。
研究人员概括了关于胰岛素抵抗增加的发现,并补充说它取决于粪菌移植后肠道微生物群的变化。
最后,与移植正常供体粪便的受试者相比,使用代谢综合征供体粪便移植肥胖和胰岛素抵抗的男性受试者时,胰岛素敏感性降低,从而显示代谢综合征中胰岛素敏感性和微生物群之间的因果关系。
发表在《胃肠病学》杂志上的一项研究也发现,粪菌移植改善了代谢综合征患者的胰岛素敏感性。
注:代谢综合征是糖尿病的前兆。
除了这些已经确定的应用,粪菌移植还有一些新兴的用途。例如,粪菌移植被探索作为治疗肥胖和2型糖尿病的潜在方法。
▼
在过去的几年中,肠道微生物群在致癌过程中的作用已得到越来越多的认可。肠道微生物失调和个别细菌可以通过激活致瘤途径、诱发炎症和破坏宿主DNA来诱发癌症或影响癌症进程。
细菌影响癌症的具体机制
几种细菌拥有或产生促进β-连环蛋白与E-钙粘蛋白分离的蛋白质,激活参与癌发生的β-连环蛋白信号通路。
肠道生态失调导致细菌衍生的短链脂肪酸的产生减少。肠道生态失调通过Toll 样受体 (TLR) 的微生物相关分子模式发挥促炎作用,增加细胞产生促炎因子,从而增加致癌作用。
除了诱发炎症外,许多细菌还能够通过释放特定代谢物来破坏DNA,从而促进癌症进展。
β-连环蛋白(β-catenin)是一种细胞骨架蛋白,主要位于细胞膜,在乳腺上皮中,β-catenin游离量较少,通过介导与E-cadherin的相互作用,将细胞外黏附因子与细胞质内细胞骨架相互连接,以保持质膜的完整性,并参与细胞的黏附、迁徙与转移等过程。
β-catenin异常表达时脱离细胞间连接进入细胞质或细胞核,将失去介导细胞间链接、黏附的功能,并引起某些癌基因的表达,导致细胞癌变、转移。
β-catenin还是介导Wnt信号传导的关键分子,其在细胞核中决定着Wnt信号通路的开放或关闭,通过调节基因表达过程,在乳腺发育以及肿瘤发生的调控中起到了重要的作用。
细胞膜中β-catenin的丢失以及细胞质、细胞核中β-catenin的高表达都可以经过不同的调节途径独立发生,诱导乳腺癌的发生,并影响乳腺癌的预后。
E-钙粘蛋白(E-cadherin)是钙黏蛋白家族成员,在上皮细胞中连接细胞骨架与细胞外环境,参与细胞信号传导。在正常乳腺组织中,E-cadherin在肌上皮中呈颗粒状膜阳性,在腺上皮细胞中呈细胞膜强阳性,是区分DCIS和小叶原位癌的标志物;E-cadherin几乎在所有的DCIS细胞上呈线状膜染色,而小叶原位癌的细胞膜大多不表达E-cadherin。
令人惊讶的是,特定的微生物群物种具有调节癌症治疗的功效,显著影响癌症患者的临床预后。
√粪菌移植有助于治疗黑色素瘤
黑色素瘤是一种皮肤癌,涉及产生黑色素的黑色素细胞。最近,新工具利用免疫检查点抑制剂来增强患者对肿瘤的免疫反应(这种治疗方法称为“癌症免疫疗法”),并且有趣地发现肠道微生物组调节这种反应。
注:谷禾前面的文章中也有讲过癌症免疫疗法。
在两个具有不同肠道微生物组成的小鼠中,黑色素瘤的生长及其对抗程序性死亡配体1(PD-L1)免疫疗法的反应显著不同。
如果将来自对抗PD-1治疗有积极反应的黑色素瘤患者的粪便样本移植到无菌小鼠体内,则PD-1阻断的抗肿瘤作用会得到改善。
粪菌移植似乎有望通过转移有利的肠道微生物群来增强黑色素瘤患者的抗肿瘤免疫力。
通过粪便微生物群移植治疗癌症
Chen D,et al.Int J Cancer.2019
▼
根据最近的研究,粪菌移植可能是治疗各种皮肤病症(包括湿疹和牛皮癣)的潜在治疗选择。人体肠道微生物群在免疫调节中起着至关重要的作用,肠道微生物群的改变与各种皮肤病有关。
一些案例报告了湿疹和牛皮癣患者经粪菌移植后成功治疗。
√改善了牛皮癣患者的严重程度
在研究皮肤病学杂志上发表的一项研究中,研究人员发现粪菌移植改善了四分之三的牛皮癣患者的严重程度。
√湿疹症状有了显著改善
此外,发表在美国皮肤病学会杂志上的一项研究报告说,粪菌移植使三分之二的患者的湿疹症状有了显著改善。
总之,粪菌移植显示出作为治疗湿疹等皮肤病的巨大潜力。同样重要的是要注意粪菌移植具有潜在的风险,包括感染和不良反应。
需要更多的研究来确定粪菌移植治疗皮肤病的最佳频率和剂量。因此,在考虑将其作为皮肤病的治疗选择之前,必须彻底评估粪菌移植的潜在益处和风险。
▼
关于微生物群和粪菌移植干预在心血管疾病中的作用,只有有限的证据可用,而且大多是在动物模型中获得的。
√菌群影响房颤的易感性
最近提出了老年人肠道菌群失调在房颤发病机制中的因果作用。研究表明,将患有心房颤动的老年大鼠粪菌移植移植到年轻大鼠体内会导致更高水平的脂多糖和更高的疾病易感性。
√菌群移植改善了心肌损伤
仍然在动物模型中,实验性自身免疫性心肌炎(EAM)小鼠模型中对照小鼠的粪菌移植增加了微生物的丰富度,包括厚壁菌门/拟杆菌门的比例增加,并且由于炎症减轻而改善了心肌损伤。
尽管科学文献中很少有证据表明粪菌移植策略在心血管疾病中有治疗价值,但在动物模型和人类中获得的这些数据肯定会为进一步研究提供坚实的基础。
本章小结
未来,粪菌移植也可能用于治疗自身免疫性疾病,如类风湿性关节炎和红斑狼疮,以及精神健康状况,如抑郁症和焦虑症。
值得注意的是,这些新兴应用仍处于研究的早期阶段,需要进一步的研究来确定它们的疗效和安全性。
总的来说,目前粪菌移植主要用于治疗反复发作的艰难梭菌感染和一部分胃肠道疾病,但它在治疗其他疾病和障碍方面具有巨大的潜力,正在进行研究探索其在各个医学领域的潜在应用。
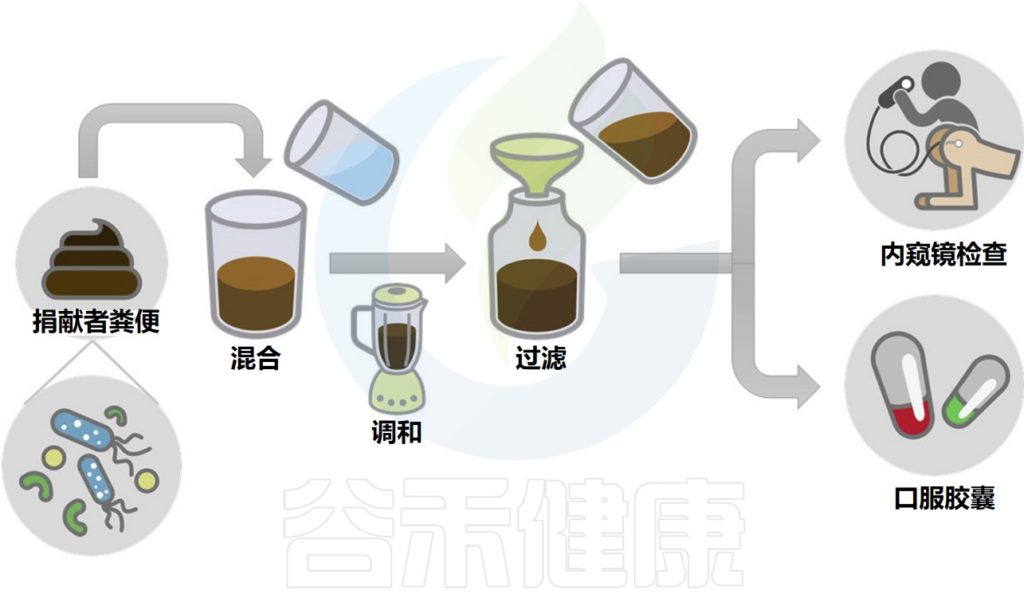
▼
移植所需的粪菌来自经过严格筛查的供体,供体捐赠样本后,用一套全自动化的机器进行分离,经过注水、搅拌、过滤、离心、沉淀等步骤,得到纯化的细菌。
粪便菌群移植过程示意图
Wang JW,et al.J Formos Med Assoc.2019
其中单单过滤的步骤就要重复数次,每一道过滤用的滤网孔径都比前一道滤网的孔径更小,最后一道滤网的孔径只有0.07毫米,基本上可以把所有的杂质去除,只剩下细菌这些微生物。
分离出来的粪菌再用生理盐水制成混悬液供移植使用。
▼
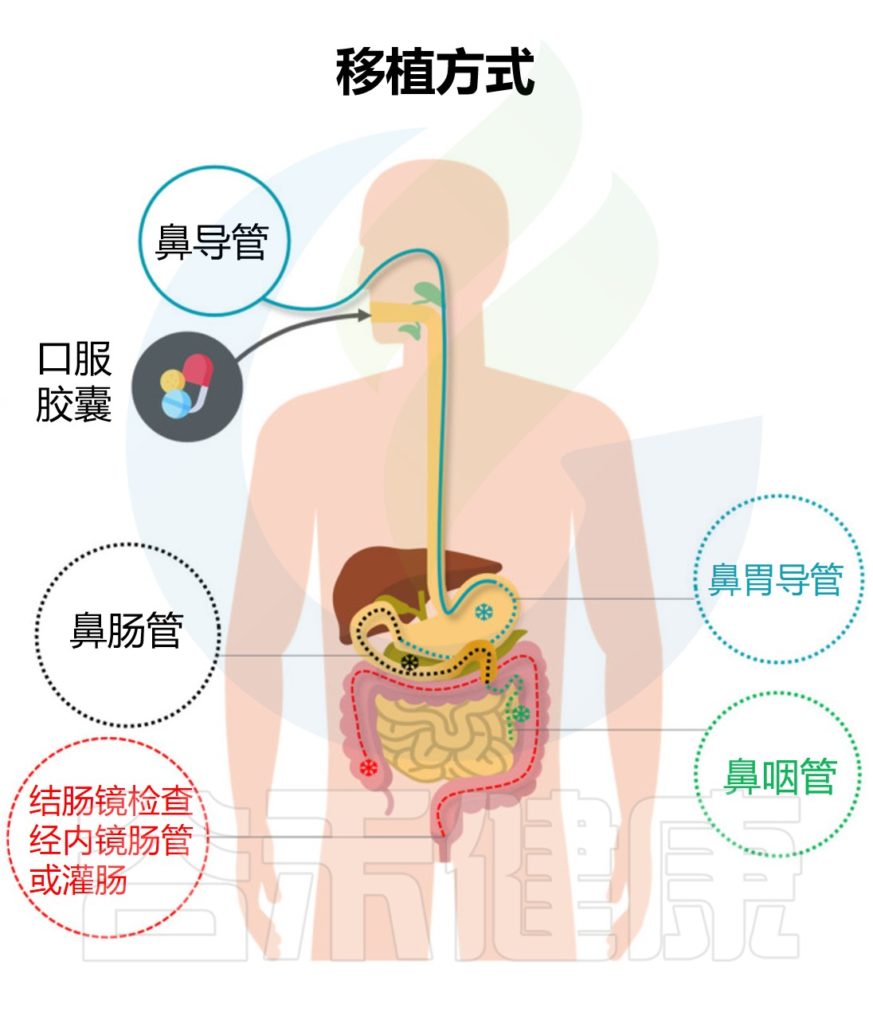
“移植”过程不是简单粗暴的将一个人的粪便直接注入患者的肠道,主要是将健康供体的粪便离心纯化,获得功能菌,然后通过上消化道、中消化道和下消化道转移至受体肠道。从而重建稳定的肠道微生态环境,缓解相关症状,治疗疾病。
★ 粪菌移植的途径与治疗目的有相关性
例如,想要治疗溃疡性结肠炎可以采取经结肠镜灌肠。要治疗慢性乙型肝炎,则是经上消化道,可以采取:经麻醉胃镜、普通胃镜、经鼻饲管,或者用粪菌胶囊,经典方式是通过麻醉胃镜。
粪便微生物群移植的方法
Biazzo M,et al.J Clin Med.2022
不同的粪菌移植方式具有各自的优缺点,下面谷禾具体来讲述经不同部位消化道进行粪菌移植的特点。
▸ 上消化道
操作简单,总体风险较低
上消化道途径中,口服给药和鼻胃管应用广泛,操作简单,总体风险较低。但在此过程中,可能会因菌液反流或吸出而导致窒息,如果机体本身存在梗阻,则移植的菌液可能无法准确到达肠道。
其中,口服途径主要指包封粪菌材料,通过将提取的粪菌与冷冻保护剂(主要是甘油)混合制备,然后进行多重包装以保护粪菌移植材料免受胃肠道环境的影响。同时,操作人员可以根据预期的释放位置进一步选择胶囊壳的材料。目前,市售的胶囊壳通常针对胃或结肠的释放,并被配制以确保细菌的存活率和定植。
降低潜在疾病传播的概率
此外,在制作胶囊的过程中,也可以去除无症状供体中可能存在的真菌、寄生虫、病毒和部分炎症介质,以降低潜在的疾病传播概率。
总的来说,通过上消化道途径的粪菌移植具有无需昂贵器械、重复使用对患者损伤小、患者耐受性好等优点。该途径适用于不能耐受鼻肠管和胃镜移植的患者,以及需要口服定制菌的患者。
▸ 中消化道
减少移植时的细菌位移
中消化道途径主要是指鼻肠移植和经内镜肠道植管术,能够实现全肠道给药。其中鼻肠管通过导丝拉伸拉直,在胃肠蠕动正常的情况下自动通过幽门,进一步减少粪菌移植时的细菌移位。
在一项随机对照试验中报道,通过鼻肠管的粪菌移植优于单独使用万古霉素治疗艰难梭菌感染,但粪菌移植组患者也出现了腹泻、痉挛和便秘等不良事件。
什么是经内镜肠道植管术?
经内镜肠道植管术(TET)具体指的是在内镜下辅助植入固定管并固定在肠道深处,同时外端沿肠道与外部相通。TET主要包括结肠途径介导的TET和中胃肠道途径介导TET。
结肠通路介导的TET需要应用结肠镜检查,并且使用这种途径进行重复粪菌移植是昂贵的。相比之下,中胃肠道途径介导的TET更方便,更容易维持。
由于中胃肠道介导的TET不需要在内镜手术后通过X射线或其他医疗仪器进一步确认肠道位置,对于无法进行结肠镜检查肠道准备的患者,或需要重复粪菌移植的患者,中胃肠道中介导的经内镜肠道植管术也是主要选择。
▸下消化道
下消化道通路主要指灌肠、结肠镜检查、结肠通路介导的经内镜肠道植管术等操作。
灌肠侵入性较小且相对简单
其中,灌肠是一种侵入性较小且相对简单的操作。灌肠法患者耐受性好,不需要昂贵的器械,有效降低了手术风险。但这种方法需要将输注的粪悬液保留较长时间,患者保持仰卧位以减少粪便排泄,同时反复灌肠易于被患者接受。
注意:粪菌移植的细菌是否可以通过灌肠的方式保留在肠段的命题尚不明确,而且这种方式也伴随着无法到达结肠和脾曲的风险,需要多次灌注才能获得疗效和更长的手术时间,以弥补肠道菌群保留率低的缺陷。同时,由于细菌液的滞留,这种方法可能不适用于肛门括约肌松弛或尿失禁的患者。
此外,在结肠镜检查和结肠通路介导的经内镜肠道植管术方面,它具有多种优势,主要包括:
(1)能够完整显示结肠和活检可疑组织的情况,有利于疾病的诊断、识别和分期;
(2) 允许操作人员直接评估肠道炎症,并在适当的部位注入足量的供体粪便细菌;
(3) 可将肠道菌群准确移植到受影响的肠段,并进一步将细菌保留在目标肠段;
(4)可以准确地输入足量的供体粪便菌,可以在一定程度上提高治疗效率。
然而,这些手术也伴随着部分不良事件,包括加重肠道反应。
粪菌移植的主要操作方式及优缺点


Zhang YW,et al.J Bone Miner Metab.2022
▼
粪菌移植的安全性主要与两方面因素有关,即供体的生物安全性、移植的操作安全性。
•供体的生物安全性
供体不仅要筛查排除乙肝、丙肝、梅毒、艾滋病、这些已知传染病,还要排除寄生虫、条件致病菌感染,甚至连饮食习惯、排便习惯、情绪状态、肿瘤家族史都要进行调查,不符合要求的都要排除。
供体的筛查可以说是“百里挑一”,要成为一个合格的供体是非常困难的。所有这些筛查工作都是为了保证供体的生物安全性。
•移植的操作安全性
移植的操作安全性主要与移植的方式有关。经胃镜、结肠镜的移植操作风险与常规胃镜、结肠镜检查的风险基本一致。
注:在后面的章节我们会详细讲述粪菌移植操作的具体要求。

为了评估粪菌移植的安全性,上海同济粪菌移植工作组主任等研究人员对8547名采用过粪菌移植的患者进行跟踪研究,得出以下结论:
1、回顾性分析数据表明,粪菌移植的短期和长期(从2周到5年)安全性都是比较好的,患者不要担心;
2、短期不良事件与粪菌移植给药方法有关,多为轻中度、较短暂,包括恶心、呕吐、腹泻等,只有42名(0.5%左右,很低比例)患者发生胃肠症状恶化;
3、长期随访期间的新发疾病主要是胃肠道和呼吸道疾病,发病率与一般人群无异;
4、 随访期间无粪菌移植相关感染传播事件,11例死亡与粪菌移植无关;
5、通过规范适应症和给药时机后,短期安全性和患者满意度升高。
所以,粪菌移植还是比较安全的。
粪菌移植已被证明是治疗多种胃肠道疾病的有效方法,然而,粪菌移植的潜在长期影响以及粪菌移植的最佳频率和剂量仍未得到很好的了解。
本章节旨在回顾有关粪菌移植潜在长期影响的现有文献,并为未来的研究和临床实践提供建议。
▼
虽然粪菌移植已被证明在短期内有效,但对粪菌移植长期影响的研究有限,粪菌移植对肠道微生物组和整体健康的长期影响仍不清楚。
✦可能导致抗生素抗性基因的转移
粪菌移植是一种医疗程序,它将健康捐赠者粪便中的微生物移植到受体的肠道中,以恢复或改善肠道微生物平衡。虽然粪菌移植已被证明在治疗某些疾病方面非常有效,但它的使用也存在一定争议。
一些研究表明,粪菌移植可能会产生意想不到的后果,例如抗生素抗性基因的转移或致病菌的引入。
✦病毒或其他感染因子可能通过粪菌移植传播
此外,人们还担心病毒或其他感染因子可能通过粪菌移植传播。需要更多的研究来充分了解粪菌移植的长期影响,并制定安全有效地使用该程序的指南。
▼
✦粪菌移植的监管
粪菌移植的监管因国家而异,一些国家如美国对其使用采取更为严格的方法。在美国,粪菌移植被美国食品药品监督管理局作为生物制品进行监管,只批准用于治疗未对其他治疗方法产生反应的复发性艰难梭菌感染。
在欧洲,粪菌移植通常被视为医疗程序,并由国家卫生部门进行监管。
✦粪菌移植的伦理问题
粪菌移植的伦理考虑包括知情同意、捐赠者选择以及对捐赠者和受体的潜在风险。捐赠者必须接受广泛的筛查,以确保他们健康且没有传染病,并且必须同意他们的粪便用于粪菌移植。
受体也必须提供知情同意,并且必须完全了解该程序的潜在风险和好处。粪菌移植的主要争议之一是该程序缺乏标准化。目前没有粪菌移植的标准化方案,粪便成分在捐赠者之间可能会有很大的差异。
▼
✦粪菌移植的剂量会影响其疗效
目前,粪菌移植的最佳频率和剂量仍不确定。一些研究表明,粪菌移植的剂量可能会影响其疗效,但目前还没有确定最佳剂量的标准。
✦一般需要多次粪菌移植
关于粪菌移植的频次,目前尚无明确的建议。一些研究表明,单次粪菌移植可能无法完全恢复肠道微生物群的平衡,需要多次粪菌移植才能达到最佳疗效。
单次粪菌移植可以有效治疗某些疾病,如复发性艰难梭菌感染。然而,对于其他疾病,可能需要多次粪菌移植才能获得最佳效果。
但是,多次粪菌移植也可能增加感染和其他不良反应的风险。因此,粪菌移植的频次应根据患者的具体情况和病情进行个体化的决策。
总之,粪菌移植的最佳频次和剂量应根据患者的具体情况和病情进行个体化的决策,同时应注意监测和预防不良反应的发生。
需要更多的研究来确定粪菌移植的最佳频率和剂量,以便在临床实践中更好地使用该程序。
根据《上海市菌群移植技术管理规范(2021年版)》,以下是部分粪菌移植操作要求:
▼
(一)严格遵守粪菌移植治疗操作规范和诊疗指南,严格掌握粪菌移植治疗适应证和禁忌证。
▷粪菌移植适用于治疗以下消化系统疾病
(1)艰难梭状芽胞杆菌感染:复发性/难治性艰难梭状芽胞杆菌感染。
(2)其他消化系统疾病:溃疡性结肠炎、克罗恩病、功能性便秘、肠易激综合征、菌群紊乱相关腹泻等。
▷粪菌移植治疗的其他疾病
(1)神经系统疾病:帕金森病、阿尔兹海默症、癫痫等。
(2)精神疾病:自闭症、情绪障碍、多动症、抽动症等。
(3)代谢相关疾病:代谢综合征、糖尿病等。
(4)肿瘤相关疾病:免疫治疗及放化疗所致肠炎等。
(二)除复发性/难治性艰难梭状芽胞杆菌感染外,出于其他治疗目的使用粪菌移植治疗均须以临床研究形式开展,并通过本机构伦理审查与备案。
(三)实施粪菌移植治疗前应当向患者及其家属告知治疗目的、风险、注意事项及可能发生的并发症等,并签署知情同意书。
(四)医疗机构应当建立完整的电子化临床数据库及严格的术后随访制度,在完成每例粪菌移植治疗后应当按照有关规定将治疗相关信息上报卫生健康行政部门。
▼
(1)有明确与肠道菌群紊乱有关的肠道内疾病,如肠功能障碍性疾病、肠道感染性疾病(复发性艰难梭菌感染)、肠道炎症性疾病及假性和麻痹性肠梗阻等;
(2)有明确与肠道疾病或者肠道菌群紊乱有关的肠道外疾病,如便秘合并帕金森病、便秘或腹泻合并自闭症、便秘或腹泻合并过敏性皮炎;
(3)患者需充分理解粪菌移植治疗过程及机制,并签署知情同意书。
!
出现以下情况之一不宜进行粪菌移植:
(1)肠道大面积溃疡、出血;
(2)移植通道梗阻;
(3)移植操作本身(内镜、置管、灌肠、经口饮食)的禁忌证;
(4)未通过伦理审核的临床试验或患者拒绝接受粪菌移植。
(5)生命体征(体温、心率、血压、呼吸)不稳定;
(6)严重免疫系统紊乱;
(7)孕妇或哺乳期妇女;
(8)代偿或者失代偿肝功能障碍。
▼
(1)个人情况:作息规律,饮食健康,家庭和睦,无不良性交,无吸烟、饮酒、吸毒等嗜好,无药物成瘾,近6个月未接种过疫苗或参加药物试验,近6个月未接受纹身或出现皮肤破损,近6个月无热带地区旅居史;无胃肠道病变家族史,无恶性肿瘤家族史,无传染病家族史;非孕期,非经期。
(2)开展病史和身体检查,评估供体的健康状况和是否符合粪菌移植的供体资格;选择无传染病史、无抗生素使用史、无消化系统疾病等方面的供体。
身体状态:年龄18-30周岁,体重指数(BMI)18.5-23.9 kg/m2,儿童医院开展粪菌移植,供体要求可放宽至15-30周岁。
心理状态:心理科医师或心理咨询师访谈认定心理状态良好;抑郁自评量表(SDS)、焦虑自评量表(SAS)、匹兹堡睡眠质量指数(PSQI)等评分正常。
(3)进行多项检测,如血液化验、粪便检测、病原菌检测、URTI检测等,确保找到适合的健康供体;以确保其不携带有害微生物和多重耐药菌。同时,需要考虑供体的肠道微生物组成和短链脂肪酸水平,以选择最适合的供体。
血液学检测:包括血常规、肝肾功能、电解质、C反应蛋白正常,肝炎病毒、HIV、梅毒、EB病毒、巨细胞病毒、COVID-19抗体、线虫、阿米巴等病原检测阴性;
粪便检测:粪便常规检查正常,隐血实验阴性,艰难梭菌、弯曲菌、沙门菌、志贺菌、产志贺毒素大肠杆菌及虫卵、小肠结肠耶尔森菌、致病性弧菌(副溶血弧菌、霍乱弧菌)、气单胞菌、阿米巴、孢子、诺如病毒、轮状病毒和新型冠状病毒(COVID-19)等病原学检测阴性。
(4)评估供体的肠道微生物组成,多样性和稳定性,包括菌群、真菌群和病毒群,并建立数据库;在选择供体时,除了肠道微生物群的总体多样性外,还应考虑到益生菌的丰富度,如乳酸菌、双歧杆菌等,选择粪便丁酸浓度高的供体微生物组。
(5)根据评估结果选择合适的供体,注意进行家族成员偏向,判别成熟度和完整性,同时避免与患者组群共同优势菌种重合等;一些研究表明,供体微生物组成与受体的微生物组成相似,可以提高移植成功率。
(6)移植前需对受体进行评估和准备,包括清洁肠道和停用抗生素等;为了提高移植效果,一些研究还使用抗生素预处理和肠道灌洗等方法来增强供体菌株的移植。
限食耐受性:通过限食实验完成。部分受体存在食物过敏及食物不耐受(如鸡蛋、牛奶等),根据受体要求,供体需在捐赠粪便前5天限制食物种类,如该供体不可耐受,则不可作为本次移植的捐赠者。
(7)进行移植后,对受体的微生物组成进行多次采样和分析,以评估移植效果和持续时间;
①每2个月复查以上条目,仍然符合上述要求;
②每次捐赠的粪便均留样行宏基因或者16s rDNA测序(根据各自单位检测能力),保证菌群组成及多样性稳定;
③每次捐献粪便应留样行代谢组学检测。
(8)监测受体的生理状况和不良反应,及时采取措施进行处理。
▼
(1)粪便采集:应采用无菌容器采集,粪便重量不少于100g,性状为Bristol评分标准中3-5分方为合格,立即进入菌液制作流程,或立即密封后2-8℃保存。
(2)制作流程:每次需取不少于50g粪便与无菌生理盐水以1∶3比例混合,充分搅拌混匀后过滤,从粪便排出体外至菌液制作完成应保证在2小时以内,整个处理过程应在无菌环境下操作。
注:为了避免感染窗口期的问题,菌液或胶囊制备后应待21 天后经过病毒细菌等检验合格后出库,每批产品留样至少6个月以供追溯。
(3)冻存与复融:菌液制备完成后置于-80℃保存,6个月之内使用不影响疗效;如置于-20℃保存,应在1-4周内使用。使用前置于室温复融,6小时内输注;如使用水浴锅复融,则水温不得超过37℃。
(4)质控标准:每次制作完成的菌液应随机抽样,进行致病菌监测、细胞计数及宏基因或16s rDNA菌群测序。
每50mL菌液中活细胞数应不小于2.5×10^12个,同供体同批次样本应达到菌群组成一致,6个月内同供体不同批次样本应达到菌群组成差异无统计学意义。
制作完成的菌液应有独立包装与标签。如制作胶囊,还应取胶囊内容物进行细菌活性监测与活细胞计数,其中细菌活性应不低于85%,每克内容物包含活菌数不小于10^9CFU。
▼
(1)菌液输注前需结合患者自身情况及疾病种类行肠道准备。
(2)应根据指南及操作规范选择上消化道或下消化道进行菌液输注。
(3)操作时应观察患者耐受情况,并及时处理不良反应。
▼
(1)管理原则:医疗安全(不良)事件的管理,遵循预防为主、紧急处理、合理控制、防微杜渐、持续改进的原则。
(2)上报要求:凡科室职工均有权上报医疗安全(不良)事件,可通过网络、书面、电话上报,上报时限要求在不良事件发生后24小时之内。
(3)审核、处置要求:科室负责人24小时内审核、处置科室发生的不良事件,并呈报至相关职能部门。
注意:主管职能部门自接报之日起,24小时内审核、处置不良事件;对于接报的Ⅱ级及以上不良事件,应呈报至主管院领导。
主管院领导对于接报的Ⅱ级及以上不良事件,必要时组织委员会进行讨论分析,并提交院务会。
▼
(1)每次菌液输注后密切观察患者耐受情况24小时,如有不良反应立刻处理并及时上报。
(2)1个疗程治疗结束后4周内应对患者症状学和肠道菌群改善情况进行评估,粪菌移植治疗疾病的随访及疗效判断指标主要遵循原发疾病的治疗指南。移植前后重复检测粪便及血液中传染病相关指标,有条件者可行免疫相关检测。
注意:如第1个疗程结束3周后病情无改善,可尝试更换供体,重新给予1个疗程粪菌移植,若有效可继续治疗,若仍无效则应视为粪菌移植无反应性,此时不应继续给予粪菌移植,以免耽误病情;如患者接受粪菌移植治疗后症状明显改善,可重复3-5次粪菌移植后结束治疗。
随访至末次粪菌移植结束后至少8周,有条件者应进行大于1年甚至5年以上的长期随访。
随着对微生物与宿主相互作用的了解不断加深,肠道微生物群的参与已成为一种新颖、巧妙且不可忽视的调节宿主健康的方式。
粪菌移植是一种治疗胃肠道疾病的有效方法,还有望治疗其他更多疾病。未来的研究应该重点关注粪菌移植对肠道微生物组和整体健康的长期影响,并制定安全有效地使用该程序的指南。
此外,需要进一步研究如何提高粪菌移植的改善效果,供体和受体的配型以及移植肠道菌群的定植和效果的长期性,以便在临床实践中更好地使用。
主要参考文献:
Zhang YW, Cao MM, Li YJ, Zhang RL, Wu MT, Yu Q, Rui YF. Fecal microbiota transplantation as a promising treatment option for osteoporosis. J Bone Miner Metab. 2022 Nov;40(6):874-889.
Biazzo M, Deidda G. Fecal Microbiota Transplantation as New Therapeutic Avenue for Human Diseases. J Clin Med. 2022 Jul 15;11(14):4119.
Quaranta G, Guarnaccia A, Fancello G, Agrillo C, Iannarelli F, Sanguinetti M, Masucci L. Fecal Microbiota Transplantation and Other Gut Microbiota Manipulation Strategies. Microorganisms. 2022 Dec 7;10(12):2424.
Vendrik KEW, Ooijevaar RE, de Jong PRC, Laman JD, van Oosten BW, van Hilten JJ, Ducarmon QR, Keller JJ, Kuijper EJ, Contarino MF. Fecal Microbiota Transplantation in Neurological Disorders. Front Cell Infect Microbiol. 2020 Mar 24;10:98.
Wang JW, Kuo CH, Kuo FC, Wang YK, Hsu WH, Yu FJ, Hu HM, Hsu PI, Wang JY, Wu DC. Fecal microbiota transplantation: Review and update. J Formos Med Assoc. 2019 Mar;118 Suppl 1:S23-S31.
Chen D, Wu J, Jin D, Wang B, Cao H. Fecal microbiota transplantation in cancer management: Current status and perspectives. Int J Cancer. 2019 Oct 15;145(8):2021-2031.
Stallmach A, Steube A, Stallhofer J, Grunert PC, Merkel U, Hartmann M. Fäkaler Mikrobiomtransfer – Indikationen, Risiken und Chancen [Fecal microbiota transplantation: indications, risks and opportunities]. Inn Med (Heidelb). 2022 Oct;63(10):1036-1042. German.

谷禾健康
乳杆菌属(Lactobacillus)是厚壁菌门乳杆菌科下的一类革兰氏阳性菌,最早于19世纪在酸奶中发现。
乳杆菌在自然界中分布很广,在植物体表、乳制品、肉制品、葡萄酒、发酵面团、污水以及人畜粪便中,均可分离到。在人类中,乳杆菌属成员定植于胃肠道、口腔和女性泌尿生殖道。
★ 乳杆菌的特性
• 消化和代谢蛋白质和碳水化合物
• 合成B族维生素和维生素K
• 分解代谢胆汁盐
• 增强先天性和获得性免疫力
• 抑制促炎介质
• 对一系列病原体具有抗菌活性
✦乳杆菌的应用
乳杆菌被广泛应用于各种领域中:
•食品工业
乳杆菌被广泛应用于食品工业中,如酸奶、奶酪的制作中。乳杆菌能够发酵乳糖产生乳酸,使得食品呈现出酸味和口感。
•医药领域
乳杆菌被用于制作口服制剂、肠内营养制剂、抗生素辅助治疗等药物。能够调节肠道微生物群落的平衡,促进食物消化和营养吸收,缓解肠道炎症和感染。
•保健品领域
乳杆菌被应用于保健品领域,如乳酸菌饮料、乳酸菌片等。乳杆菌能够改善肠道菌群平衡,增强免疫力,促进身体健康。
•环境领域
乳杆菌被应用于环境领域,如土壤修复、废水处理等。乳杆菌能够分解有机物质,促进土壤肥力和水质净化。
★ 乳杆菌对人体健康非常重要
随着对微生物群落的研究逐渐深入,乳杆菌的作用也逐渐被人们所认识。
乳杆菌对人体有着重要的作用,在肠道健康及女性阴道健康中扮演不可或缺的角色。具有多种生理功能,能够维护菌群平衡、改善消化功能、提高免疫力,除此之外还会影响一些疾病的进展。
本文主要讨论和介绍乳杆菌的菌属特性,生态代谢特征,对人体肠道健康、阴道健康以及一些疾病的影响,在最后还介绍了一些补充乳杆菌的方法。
目录/contents
Part1:认识乳杆菌
Part2:乳杆菌与肠道健康
Part3:乳杆菌与阴道健康
Part4:乳杆菌与其他疾病
Part5:如何补充乳杆菌
Part6:结语
▸ 乳杆菌、乳酸杆菌和乳酸菌
乳杆菌属(Lactobacillus),有时也会翻译成乳酸杆菌,是革兰氏阳性兼性厌氧或微需氧的棒状细菌,通常是乳酸菌的一种。
乳杆菌属包括多种菌种,如乳酸杆菌、嗜酸乳杆菌、肠道乳杆菌等。它们通常是革兰氏阳性、非芽孢杆菌,能够在无氧或微氧条件下进行发酵,产生乳酸等有益物质。
乳酸菌包括多种菌属,如乳杆菌属、双歧杆菌属、嗜酸乳球菌属等。它们能够在无氧或微氧条件下进行发酵,产生乳酸等有益物质。乳酸菌在人体内具有多种功能,如维护肠道菌群平衡、增强免疫力、促进营养物质吸收等。
因此,乳杆菌属(乳酸杆菌)是乳酸菌的一种,而乳酸菌则是一个更广泛的概念,包括多种菌属。
▸ 形态特征
乳杆菌属于革兰氏阳性菌,呈细长的杆状,但无分枝,无芽孢,一般大小约为0.5-1.5μm×2-10μm(某些大杆菌如保加利亚乳杆菌可能更大)。菌落呈圆形、白色(有时黄色)、半透明。
细胞壁:乳杆菌的细胞壁主要由肽聚糖和多糖组成,具有较强的抗菌作用。
运动性:乳杆菌通常是非运动性的,即不具有鞭毛或纤毛。菌毛首先在鼠李糖乳杆菌(Lactobacillus rhamnosus GG)中观察到,它从细菌细胞中突出,在粘附到上皮细胞中起主要作用。
乳杆菌菌落


Talib N,et al.Molecules.2019
注:a是在MRS培养基上从开菲尔样品中分离的乳酸杆菌菌落; b是在MRS培养基上从开菲尔样品中单筛选分离的乳酸杆菌
▸ 分类及主要种群
乳杆菌属是乳杆菌科中最大的一个属。该属的成员已经有超过200种。具体见附录一。
主要有德氏乳杆菌保加利亚亚种(保加利亚乳杆菌)、嗜酸乳杆菌、干酪乳杆菌、罗伊氏乳杆菌、惰性乳杆菌等。
此外卫生部公布可用于生产普通食品的乳杆菌还有卷曲乳杆菌、德氏乳杆菌乳亚种、发酵乳杆菌、格氏乳杆菌、瑞士乳杆菌、约氏乳杆菌、副干酪乳杆菌、植物乳杆菌、唾液乳杆菌等。
乳杆菌属中最具代表性的物种以其在食品发酵中的应用而闻名,并且特定菌株已被认为具有益生菌特性。乳酸杆菌的健康方面促使许多研究人员积极筛选这些细菌的许多潜在来源。因此,在过去 15 年中观察到新物种的爆炸式增长。
尽管许多早期描述的物种被转移到新创建的属(Atopobium、Carnobacterium、Eggerthia、 Weissella 、Fructobacillus等)该属在系统发育和表型标记方面仍然存在异质性。Pediococcus属、Paralactobacillus 属与Lactobacillus 混合在一起,表明对该属进行进一步的分类学细分很可能甚至是必要的。
▸ 生长环境和条件
乳杆菌广泛存在于自然界中,包括土壤、水体、植物、动物等环境中。乳杆菌作为一种常见的益生菌,也存在于人体的口腔、肠道、阴道等部位。
乳杆菌的生长环境主要包括以下几个方面:
pH值:乳杆菌适宜生长的pH范围一般在4.5-6.5之间(某些耐酸种群可能在3.8仍能生长),其中以5.5-6.0最为适宜。
温度:乳杆菌的生长温度一般在20-45℃之间,其中以30-40℃最为适宜。
氧气需求:乳杆菌是一种厌氧菌,不需要氧气就能生长。
营养物质:乳杆菌需要一定的营养物质才能生长,主要包括碳源、氮源、矿物质、维生素等。
注:不同种类的乳杆菌对营养物质的需求可能会有所不同
▸ 乳杆菌的代谢
乳杆菌是过氧化氢酶阴性革兰氏阳性微生物,主要产生乳酸作为碳水化合物发酵的主要代谢终产物。
参与的KEGG途径具体见附录二。
▸ 乳杆菌的作用
乳杆菌是一种益生菌,对人体健康有着重要的作用。以下是乳杆菌的主要作用:
维护肠道菌群平衡:乳杆菌能够抑制有害菌的生长,维护肠道菌群平衡,防止肠道感染和炎症。
改善消化功能:乳杆菌能够分解食物中的纤维素和其他难以消化的物质,促进食物消化和营养吸收。
提高免疫力:乳杆菌能够增强肠道黏膜屏障的功能,防止有害物质进入血液循环,提高免疫力。
缓解过敏反应:乳杆菌能够调节免疫系统的反应,减轻过敏反应的症状。
降低胆固醇:乳杆菌能够降低血液中的胆固醇水平,预防心血管疾病。
改善口腔健康:乳杆菌能够抑制口腔中有害菌的生长,预防龋齿和口臭。
总之,乳杆菌对人体健康有着多方面的益处,是一种非常重要的益生菌。
• 乳杆菌也有可能是机会性病原体
乳杆菌中主要的病原菌或致病菌包括:
它们也有可能是机会性病原体,可引起脓肿、菌血症、心内膜炎、肺部感染和新生儿脑膜炎等多种感染。
注:大多数由乳酸杆菌引起的疾病都发生在免疫功能低下的人或那些有糖尿病等易感病症的人身上。
乳杆菌在人体的位置及与人体的关系
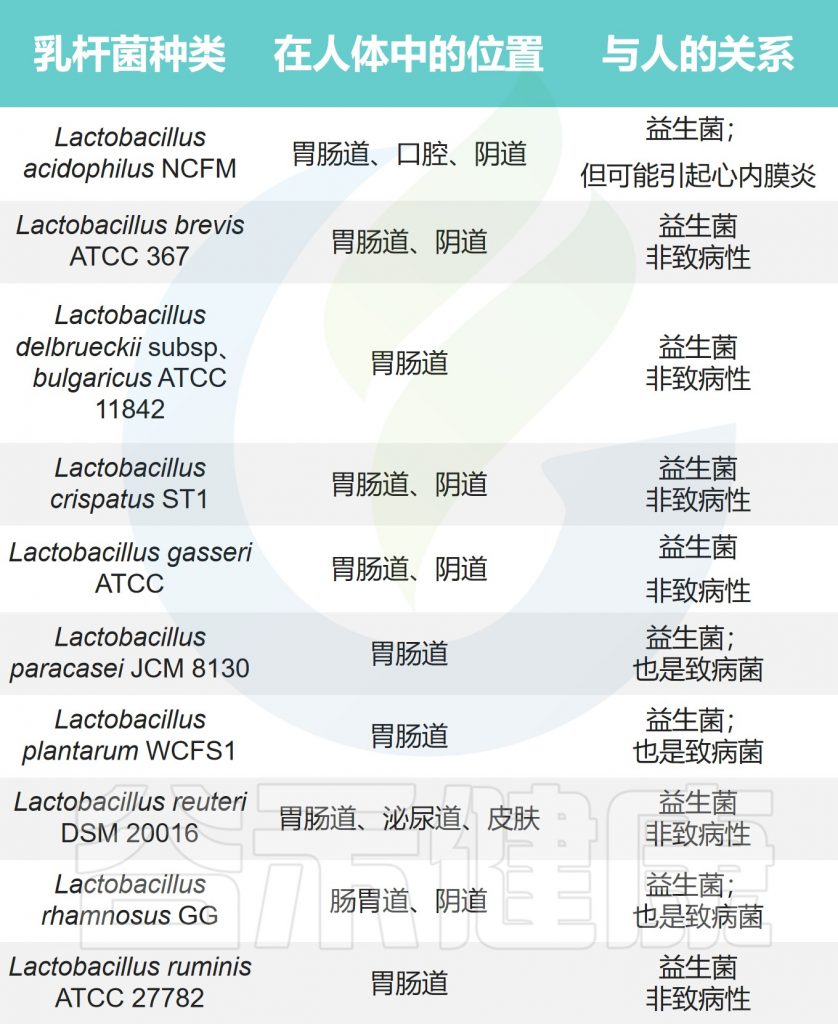

Zafar H,et al.Genes (Basel).2020
▸ 乳杆菌的表面活性分子
乳杆菌的益生菌特性被认为是由乳杆菌表面活性分子 (SAM) 贡献的。据报道,支持益生菌作用的表面活性分子包括肽聚糖、细菌多糖和磷壁酸等。
乳杆菌细胞表面结构
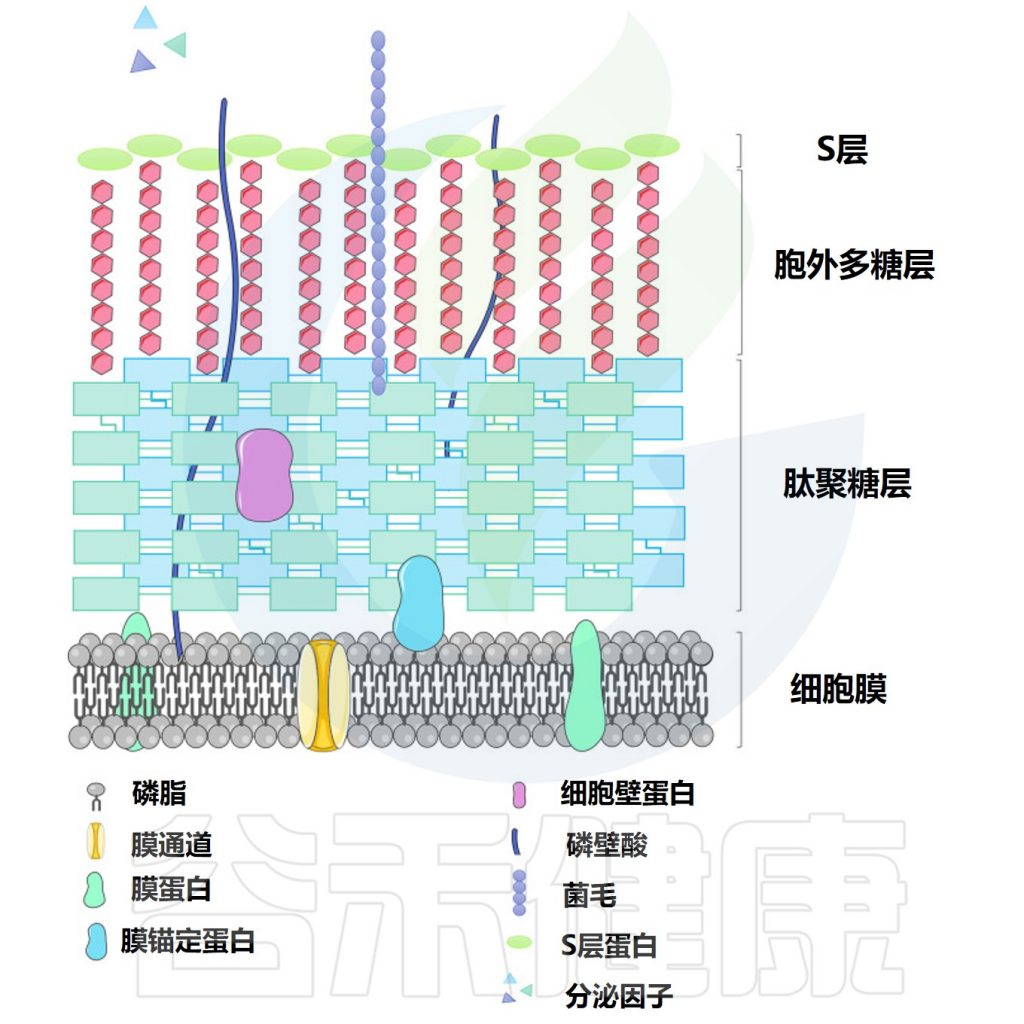

Dempsey E,et al.Front Immunol.2022
通常,乳杆菌的细胞质膜被肽聚糖和其他生物聚合物包围,即磷壁酸、S层蛋白和多糖。
• 肽聚糖
肽聚糖(PG)是一种生物聚合物,包含由N-乙酰氨基葡萄糖(GlcNAc)和N-乙酰胞壁酸(MurNAc)侧链连接的聚糖链,形成革兰氏阳性菌(如乳杆菌和双歧杆菌)的细胞表面。
改善宿主先天免疫
乳杆菌肽聚糖在改善宿主先天免疫反应方面表现出出色的免疫调节活性。例如,干酪乳杆菌(L.casei)肽聚糖能够通过toll样受体2和核苷酸结合寡聚化结构域2(NOD2)诱导小鼠腹腔巨噬细胞产生白细胞介素12。
此外,植物乳杆菌(L. plantarum CAU1055)肽聚糖通过抑制一氧化氮合酶、环氧合酶-2(COX-2)和细胞因子(TNF-α和白细胞介素-6),显示出改善小鼠巨噬细胞中一氧化氮诱导的炎症的能力。
• 脂磷壁酸
乳杆菌肽聚糖通常用磷壁酸或脂磷壁酸修饰。脂磷壁酸由磷酸甘油的聚合产生,并与细胞质膜结合。
调节宿主的信号通路
脂磷壁酸与其他表面活性分子一起调节宿主模式识别受体和宿主的几种信号通路,这些信号通路是乳杆菌的益生菌和抗病原体作用的原因。
植物乳杆菌脂磷壁酸显著抑制粪便大肠杆菌生物膜的形成,并在人类牙本质切片上形成生物膜,这表明脂磷壁酸可以作为粪便大肠杆菌感染的预防和治疗措施。
• 细菌多糖
细菌在细胞表面形成紧密连接的聚合物,并将其作为胞外多糖(EPS)(松散未附着的黏液)释放到环境中。
细菌分泌的胞外多糖对于宿主-微生物相互作用过程中的粘附和细胞识别至关重要。乳杆菌的胞外多糖还能抑制蜡样芽孢杆菌(Bacillus cereus)产生的细菌毒素。
▸ 影响乳杆菌的因素
乳杆菌对人体健康有着重要的作用。然而乳杆菌的数量和种类受到多种因素的影响,包括以下几方面:
饮食因素:饮食中富含益生元的食物,如蔬菜、水果、全麦面包等,可以促进乳杆菌的生长。相反,高脂肪、高糖、高盐等不健康的饮食习惯会抑制乳杆菌的生长。
药物因素:长期使用抗生素、非甾体抗炎药等药物会破坏肠道菌群平衡,抑制乳杆菌的生长。
生活方式因素:缺乏运动、长期处于压力状态等生活方式因素也会影响肠道菌群的平衡,抑制乳杆菌的生长。
年龄因素:婴幼儿期肠道菌群构成不稳定,乳杆菌数量较少,随着年龄的增长,乳杆菌数量逐渐增加。
综上所述,保持健康的饮食习惯、合理使用药物、积极的生活方式等都有助于促进乳杆菌的生长。
乳杆菌已被证明对改善胃肠道健康具有重要作用,本章节将具体展开讲述。
胃肠道粘膜是机体最大也是最关键的屏障部位之一,外来抗原、微生物和潜在病原体都在这里与宿主的免疫系统密切接触。
它是一种半透性屏障,允许吸收营养和免疫感应,同时限制潜在有害抗原或微生物的流入。
胃肠道屏障由四个主要元素组成:共生微生物群、粘液层——含有分泌型免疫球蛋白A(sIgA)和抗微生物肽、肠上皮细胞单层和肠道相关淋巴组织(GALT)构成胃肠道隔室中的各种免疫细胞群。
肠道屏障破坏与多种疾病相关
肠道屏障功能的丧失被认为是各种胃肠道疾病发病机制的早期事件,例如乳糜泻和炎症性肠病,以及包括I型糖尿病、肥胖症和多发性硬化症在内的全身性疾病。所以肠道屏障的完整性对于健康至关重要。
★ 乳杆菌可以增强肠道屏障
肠道屏障功能可以通过摄入非致病微生物来增强,这些微生物可以增强粘液层的物理屏障,增强对病原体的先天防御并降低肠上皮细胞的细胞旁通透性。
作为益生菌食用的乳杆菌被认为可以通过多种作用机制调节天然肠道微生物群并改善健康。如图所,益生菌通过增加粘液的产生、刺激抗微生物肽的释放和分泌性免疫球蛋白A的产生来增强肠道屏障功能,增加肠上皮细胞的紧密连接完整性并提供对病原体的抵抗力。
乳杆菌增强肠道屏障的机制
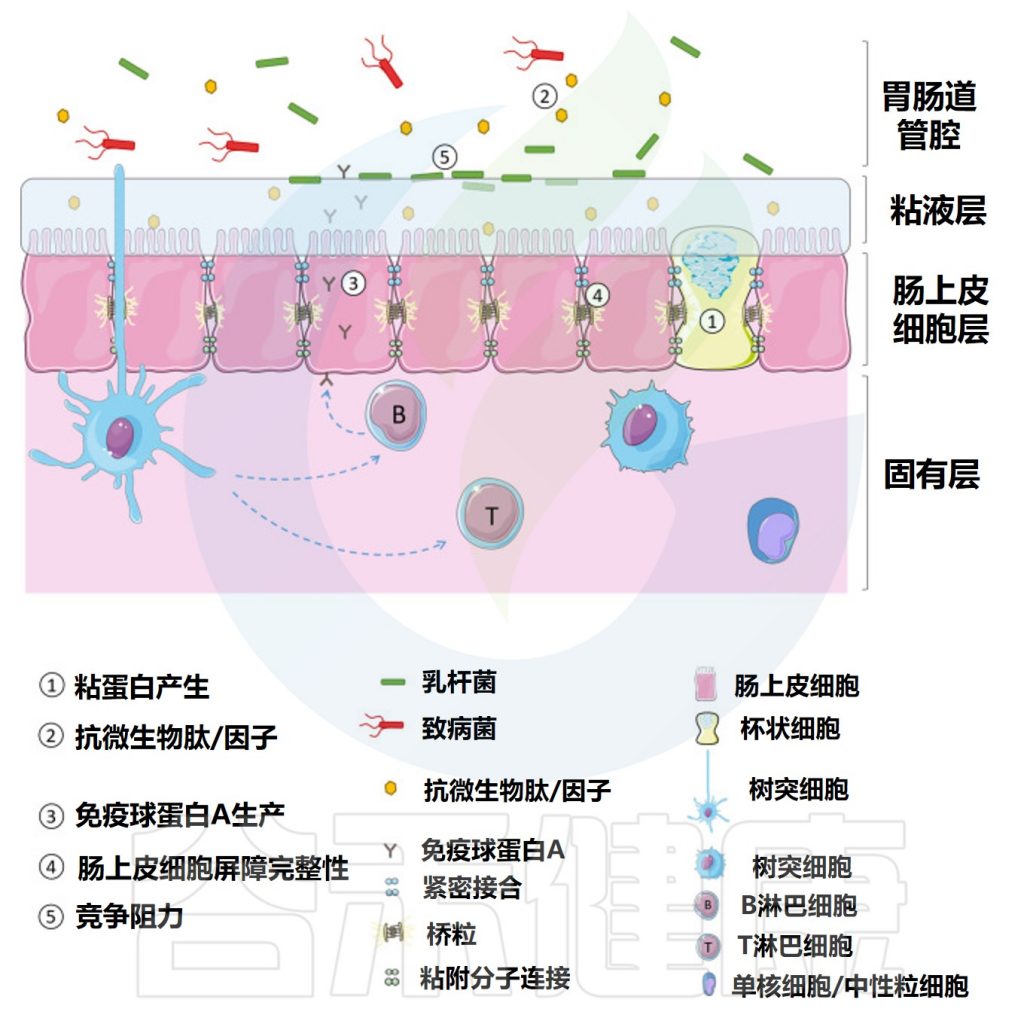

Dempsey E,et al.Front Immunol.2022
1 粘液分泌
胃肠道的杯状细胞表达杆状粘蛋白,这些粘蛋白要么粘附在上皮细胞上,要么释放到胃肠道腔中。这些粘蛋白高度糖基化并通过二硫键连接在一起形成糖蛋白基质,保护肠上皮免受肠腔内容物(含有消化酶)的影响,防止致病抗原/细菌与上皮单层之间的相互作用,并有助于胃肠道运动。
粘液层的厚度通常在50-800µm之间,在健康个体中,最靠近上皮表面的前30µm应该没有微生物。
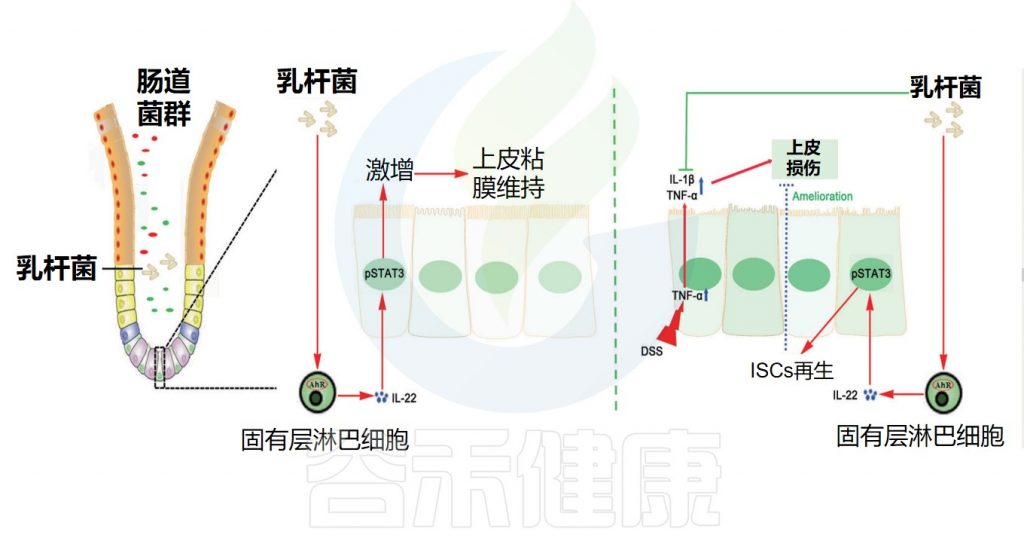
乳杆菌通过促进粘液分泌增强胃肠道屏障
乳酸菌种被认为通过促进粘液分泌来增强肠道屏障防御。体外研究表明,来自干酪乳杆菌(L.casei T21)的条件培养基可以上调受到艰难梭菌攻击的结肠上皮细胞中的粘膜保护性基因。
在体内研究方面, L. rhamnosus CNCM I-3690 最近被证明可以保护或恢复杯状细胞种群并保护小鼠在低度结肠炎症后的粘液层厚度。
类似地,施用两种罗伊氏乳杆菌(L. reuteri R2LC 或4659)菌株之一并暴露于结肠炎的小鼠表现出降低的结肠炎严重程度,归因于给予益生菌的小鼠粘液厚度增加。
2 抗菌肽、抗菌因子
宿主产生的胃肠道抗菌肽通常分为导管素和防御素。抗菌肽可以被丁酸盐激活。
丁酸盐由肠道微生物群产生,但很少有研究检查益生菌对导管素表达的影响。
防御素进一步分为β-防御素,由整个肠道的上皮细胞产生,以及α-防御素,在小肠中表达。防御素在胃肠道中组成型表达,对许多细菌、真菌和一些病毒显示出抗微生物活性。
乳杆菌诱导抗菌肽表达
嗜酸乳杆菌(L. acidophilus PZ1138)和发酵乳杆菌(L. fermentum PZ1162)显示可通过促炎机制在细胞中诱导人β-防御素-2基因的表达。除了上调促炎介质外,给肉鸡施用罗伊氏乳杆菌(L.reuteri)还与盲肠和回肠中的抗菌肽调节相关。
除了宿主衍生的抗菌肽刺激外,共生细菌还产生抗菌因子以帮助宿主屏障防御。这些因素包括短链脂肪酸、过氧化氢和细菌素。
短链脂肪酸刺激紧密连接的形成
乳杆菌产生包括乙酸盐、丙酸盐和丁酸盐在内的短链脂肪酸,这些短链脂肪酸已被证明在体外通过抑制NLRP3炎症小体和自噬来增加跨上皮电阻并刺激肠上皮细胞中紧密连接的形成。
细菌素抑制或杀死其他细菌
细菌素是由多种细菌产生的核糖体合成的热稳定肽,其功能是抑制或杀死其他细菌的生长。
革兰氏阳性菌产生的细菌素通常通过破坏膜功能发挥其抗生素作用,通常针对其他革兰氏阳性菌,但一些革兰氏阴性菌也可能易感。
植物乳杆菌产生多种细菌素,这些细菌素对食源性病原体(如李斯特菌)和食品腐败菌具有抗微生物活性,可用于食品生产以减少化学防腐剂的使用。
3 免疫球蛋白A
免疫球蛋白A的产生是胃肠道在非炎症模式下产生免疫保护的重要方式。IgA二聚体(由位于淋巴结或固有层的肠道B细胞分泌)与上皮细胞基底外侧表面的受体相互作用,转移到上皮细胞表面并作为分泌型免疫球蛋白A(sIgA)释放。
分泌型免疫球蛋白A(sIgA)是一种非常重要的抗体分子,在人体抵抗疾病的第一道防线起决定性作用。
sIgA主要通过结合饮食抗原和粘液中的潜在病原体并下调共生细菌上促炎细菌表位的表达来促进肠道中合适的共生细菌群落的维持。
此外,sIgA通过阻断参与上皮粘附的微生物成分增强肠道屏障,促进上皮内防御病原体和微生物产物。
乳杆菌可以增加小肠中slgA水平
已知包括副干酪乳杆菌(L.paracasei MCC1849)、加氏乳杆菌(L.gasseri SBT2055)和植物乳杆菌在内的各种乳杆菌菌株可增加小肠中的sIgA水平。
在一项针对12至24个月大儿童的临床试验中,补充植物乳杆菌增加了sIgA粪便滴度,并且观察到这与TGF-β1/TNF-α比率之间存在显著正相关。
4 上皮细胞屏障
如前所述,肠上皮细胞形成单层细胞,充当肠腔外部环境和宿主免疫系统之间的物理屏障。该屏障的完整性由紧密连接确保,紧密连接是将细胞紧密结合在一起的多蛋白复合物以及粘附连接、间隙连接和桥粒。
紧密连接位于上皮细胞的顶端侧。它们由跨膜蛋白组成,这些蛋白在细胞外与邻近细胞中紧密连接的类似蛋白质相互作用,在细胞内通过封闭小带蛋白和丝状肌动蛋白与细胞自身的细胞骨架相互作用。
病原菌感染会破坏紧密紧密连接蛋白
在慢性炎症性疾病中观察到紧密连接完整性的丧失,在肠道病原体如艰难梭菌、大肠杆菌、鼠伤寒沙门氏菌、霍乱弧菌等的感染中观察到破坏紧密连接蛋白以突破胃肠道屏障的机制。
乳杆菌增强紧密连接蛋白和闭合蛋白表达
已证明鼠李糖乳杆菌上调Caco-2细胞中的紧密连接蛋白1(ZO-1)的表达。这种益生菌菌株可提高ZO-1的表达水平并增强闭合蛋白(claudin-1)的分布,作为对抗肠出血性大肠杆菌感染的保护机制。
使用各种植物乳杆菌菌株(L. plantarum WCSF1、L. plantarum CGMCC 1258和L. plantarum MB 452)也观察到紧密连接蛋白和闭合蛋白的表达增加。
乳杆菌增强肠上皮屏障保护作用

Hou Q,et al.Cell Death Differ.2018
增加E-钙粘蛋白表达,稳定粘附连接
乳杆菌还可以通过增加E-钙粘蛋白的表达,以及通过增强β-连环蛋白的磷酸化来加强E-钙粘蛋白/β-连环蛋白复合物(将粘附连接到细胞骨架)来稳定粘附连接。
钙黏蛋白E——又名上皮细胞钙粘素和CD324,是一种钙依赖性的细胞粘附分子。
在小肠屏障功能的临床研究中,活检样本表明植物乳杆菌(L. plantarum strain TIFN101)和较小程度上L. plantarum WCFS1和CIP104448调节紧密连接和粘附连接蛋白基因表达的增加。
5 竞争结合位点抵抗病原体
乳杆菌还通过竞争肠上皮细胞、粘液层中的糖蛋白或细胞外基质的纤溶酶原上的结合位点来帮助肠道屏障抵抗入侵病原体。
为了促进与宿主细胞的相互作用,乳杆菌属物种在其外表面展示各种不同的成分。这些可能包括细胞壁蛋白、S层蛋白、菌毛蛋白和月光蛋白。
这些表面蛋白促进乳杆菌与宿主的粘附,例如在几种乳杆菌菌株中发现的LPXTG蛋白是与肽聚糖层共价结合的细胞表面蛋白,可以结合粘液和上皮细胞。
通过阻断病原体与位点结合显示抗病毒活性
嗜酸乳杆菌(L. acidophilus ATCC)的S层蛋白通过阻断病原体与C型凝集素受体(DC-SIGN)的粘附而显示出抗病毒活性
C型凝集素受体(DC-SIGN)是一种强烈促进病毒感染的附着因子
如前文所述,乳杆菌通过促进粘液产生和屏障相关蛋白,分泌抗微生物物质(如短链脂肪酸、细菌素)抑制病原体的生长或杀死病原体,来保护肠道屏障免受感染,并通过调节宿主对病原体的免疫反应,防止病原体粘附和竞争结合位点。
★ 乳杆菌可以抑制病原菌感染
因此,乳杆菌能够预防某些细菌感染引起的肠道损伤。乳杆菌已被证明可以抑制病原菌感染的发展,例如艰难梭菌和产气荚膜梭菌、空肠弯曲杆菌、肠炎沙门氏菌、大肠杆菌、金黄色葡萄球菌和耶尔森氏菌等。
由感染引起的两种主要胃肠道疾病,幽门螺杆菌感染和抗生素相关性腹泻,已被证明可以极大地受益于乳杆菌,概述如下。
幽门螺杆菌感染与乳杆菌
幽门螺杆菌感染是世界上最常见的细菌感染之一,全球一半以上的人口受到感染。幽门螺杆菌感染胃的上皮细胞内壁,导致消化性溃疡病、慢性胃炎和胃癌等疾病。
许多感染者没有症状,20%的感染患者会出现胃炎症状、胃或十二指肠溃疡或非霍奇金胃淋巴瘤。
目前推荐的幽门螺杆菌感染治疗方法包括多种抗生素药物和质子泵抑制剂,但随着幽门螺杆菌抗生素耐药性的上升,这种治疗的有效性正在下降。
在各种随机对照试验中,添加乳杆菌(分别为干酪乳杆菌DN-114 001和干酪乳杆菌Shirota)和嗜酸乳杆菌后生元已被证明可以提高该疗法的疗效。
√使幽门螺杆菌活力丧失
来源于干酪乳杆菌(Lactobacillus caseistrain Shirota)的无细胞废培养上清液在体外表现出对幽门螺杆菌的杀菌活性。约氏乳杆菌(L. johnsonii NCC533)和嗜酸乳杆菌均导致幽门螺杆菌活力丧失。
此外,来自这三种乳杆菌菌株的培养物上清液导致幽门螺杆菌的形态改变为U形或球形体,这是细菌的休眠形式,而球形体定植和诱导炎症的能力较低。
已知约氏乳杆菌NCC 533和干酪乳杆菌Shirota也能产生对幽门螺杆菌有活性的细菌素。
√影响幽门螺杆菌运动
幽门螺杆菌是一种螺旋形细菌,具有多个鞭毛,使其能够在胃粘液层中游动并与上皮细胞相互作用,这是在胃中定植所需的能力。
干酪乳杆菌已被证明会导致幽门螺杆菌失去鞭毛运动,这是由于其转化为没有鞭毛的休眠形式,以及通过分泌抑制游泳能力的小型抗微生物化合物。
类似地,约氏乳杆菌(L. johnsonii NCC533)也分泌抑制幽门螺杆菌游泳能力的化合物。
√阻止幽门螺杆菌粘附
为了在胃的低pH中生存,幽门螺杆菌将脲酶表达为一种表面蛋白,以中和周围的酸性环境。嗜酸乳杆菌和约氏乳杆菌的培养物上清液已被证明可降低幽门螺杆菌的脲酶活性。
就粘附性而言,嗜酸乳杆菌阻止了幽门螺杆菌粘附到人HT-29细胞上,导致粘附细胞死亡,并降低了导致其裂解的剩余粘附细胞的脲酶活性。
HT-29细胞——人结直肠腺癌细胞
抗生素相关性腹泻与乳杆菌
抗生素相关性腹泻 (AAD) 是由抗生素破坏肠道正常微生物群引起的,其症状从轻度腹泻到更严重的疾病,如伪膜性结肠炎 (PMC) 。
抗生素相关性腹泻发生在5-30%接受抗生素治疗的患者中,无论是在抗生素治疗期间还是在治疗停止后长达2个月内。
与抗生素相关性腹泻相关的主要病原体之一是艰难梭菌,它导致10-30%的轻症病例和90-100%的重症病例。
注:尽管产气荚膜梭菌、金黄色葡萄球菌和催产克雷伯氏菌等其他微生物与这种疾病有关,但它们并不常见。
√乳杆菌治疗是预防抗生素相关性腹泻的有效措施
虽然在这种情况下益生菌的作用机制尚不清楚,但它们的功效似乎是维持肠道菌群、战胜病原菌、保护肠道屏障功能和潜在的免疫调节。用包括鼠李糖乳杆菌和格氏乳杆菌(L. gasseri)在内的几种乳杆菌菌株治疗已被证明是治疗抗生素相关性腹泻的有效预防措施。
一项分析总结了51项随机对照试验,发现鼠李糖乳杆菌比其他益生菌更有效,但干酪乳杆菌对艰难梭菌感染最有效。另一项研究显示了类似的结果,得出结论认为安全地给予鼠李糖乳杆菌以预防抗生素相关性腹泻并另外控制急性胃肠炎的症状。
前文中有提到乳杆菌在宿主的胃肠道中发挥免疫作用,可增强肠道屏障并保护免受潜在病原体的侵害。
✦乳杆菌的免疫调节作用
乳杆菌的免疫调节作用是通过释放细胞因子实现的,包括白介素、肿瘤坏死因子、干扰素、转化生长因子和来自免疫细胞的趋化因子。
炎症过程取决于促炎细胞因子与抗炎细胞因子,益生菌可以通过这种方式以免疫调节或免疫刺激的方式发挥作用。
降低炎症反应
免疫调节益生菌可降低炎症反应,保护宿主免受自身免疫性疾病、炎症性肠病和过敏的侵害,其特点是产生白细胞介素-10和调节性T细胞。
白细胞介素-10是由单核细胞、T细胞、B细胞、巨噬细胞、NK细胞产生的抗炎细胞因子,可抑制促炎细胞因子、趋化因子和趋化因子受体,从而防止肠道炎症。
免疫刺激性益生菌通过产生白细胞介素-12来激活NK细胞和发育Th1细胞,保护宿主免受感染和癌症发展,并通过平衡Th1和Th2的产生来保护宿主免受过敏。
越来越多的证据表明,乳杆菌有可能预防或治疗某些炎症性疾病:
下调白细胞介素12的产生
L.casei Shirota、L.johnsonii JCM 2012 和L.plantarum ATCC 14917的肽聚糖已被证明可通过Toll样受体2下调白细胞介素-12的产生。
增强了免疫反应
来自鼠李糖乳杆菌(L.rhamnosus CRL1505)的肽聚糖在小鼠鼻腔给药时表现出先天性和适应性免疫反应的增强,改善了Th2型免疫反应。
引发抗炎反应
植物乳杆菌的脂磷壁酸已被显示通过抑制 白细胞介素-8在人和猪肠上皮细胞中引发抗炎反应。
鼠李糖乳杆菌的菌毛的敲除突变体表明,这些菌毛不仅对粘附至关重要,而且具有免疫调节作用。
在小鼠中,源自德氏乳杆菌保加利亚亚种发酵酸奶的胞外多糖具有免疫刺激作用,激活自然杀伤细胞并诱导脾脏产生γ干扰素。
✦乳杆菌代谢物的免疫调剂作用
短链脂肪酸的抗炎作用
一些免疫调节作用是由乳杆菌的代谢物介导的,例如短链脂肪酸,特别是丙酸盐、乙酸盐和丁酸盐。这些后生元与肠上皮细胞上的特定受体结合,以抑制嗜中性粒细胞和巨噬细胞的促炎活性和Treg抑制作用。
丁酸盐灌肠剂已被证明有效并成为转移性结肠炎的公认治疗方法,尽管这被认为是由于对平滑肌的松弛作用。
减少氧化应激作用
乳酸杆菌还能够产生抗氧化剂,如谷胱甘肽 (GSH),并能减少氧化应激。在结肠炎大鼠模型中,保加利亚乳杆菌的两个菌株已被证明可以减少脂质过氧化,增加抗氧化酶的测量值,并减少氧化应激。
在胃损伤小鼠模型中,发酵乳杆菌(L.fermentum Suo)显著降低了丙二醛(一种氧化损伤的量度)浓度和IL-6、IL-12、TNF-α 和IFN-γ的血清浓度。
在健康的人类受试者中,干酪乳杆菌与益生元菊粉一起服用,显著降低了丙二醛和谷胱甘肽二硫化物(另一种氧化测量)的浓度,并增加了抗氧化指标的浓度。
✦分泌蛋白化合物调节免疫
乳杆菌还可以通过分泌蛋白质化合物来调节免疫系统。从鼠李糖乳杆菌(L. rhamnosus GG ATCC 53103)释放的蛋白质p40和p75都激活了Akt信号通路,抑制了肿瘤坏死因子诱导的人和小鼠结肠上皮细胞和小鼠结肠外植体细胞凋亡。
减少炎症损伤
在诱导小鼠结肠炎之前用鼠李糖乳杆菌进行预处理可显著减少结肠炎症和损伤,抑制细胞因子诱导的细胞凋亡并减少过氧化氢诱导的紧密连接破坏。
在鼠李糖乳中发现的两种可溶性蛋白质p40和p75的消耗消除了这些抗炎作用。鼠李糖乳杆菌通过分泌可溶性肽增加小鼠结肠细胞中热休克蛋白HSP25和 HSP72 的产生,这些肽通过激活MAPK信号转导途径发挥作用。
乳杆菌属不仅在人体肠道健康中起着重要作用,在女性阴道健康与相关疾病中的作用也不容忽视。
阴道微生物组在人类阴道健康方面起着重要作用。使用高通量宏基因组和16S rRNA测序,已在人类阴道中鉴定出超过250种细菌。
其中,乳杆菌是健康阴道中最常检测到的微生物,包括卷曲乳杆菌、惰性乳杆菌、詹氏乳杆菌和格氏乳杆菌。
几十年来,乳杆菌通过产生有机酸、细菌素和其他抗菌化合物来防止病原体入侵,被认为对阴道生态有益。
健康阴道中主要乳杆菌的相对丰度决定了细菌群落群的类型,称为群落状态类型(CST)。群落状态类型分为CST I、II、III、IV、V五种。
每个群落均由卷曲乳杆菌(L. crispatus)、加氏乳杆菌(L. gasseri)、惰性乳杆菌(L. iners)、包括乳杆菌和细菌性阴道病相关细菌(BVAB)在内的多种微生物群落占主导地位。
✦阴道微生物群会随时间和外因变化
人类阴道微生物群组成在不同的生命阶段会发生变化,包括婴儿、青春期、怀孕和更年期。
荷尔蒙的变化、抗生素的不受控制的使用、月经和阴道冲洗是导致人类阴道微生物群暂时变化的常见因素。
阴道生态系统破坏的特征是乳杆菌种类的枯竭和非乳杆菌微生物的过度生长。通常,厌氧菌的过度生长会导致异常情况,例如细菌性阴道病以及妊娠相关并发症。
与肠道菌群相比,健康个体阴道微生物环境的一个典型特征是较低的细菌多样性。阴道生态失调的定义是细菌多样性高和厌氧菌混合,通常与多种妇科疾病有关。
惰性乳杆菌(L. iners)存在于健康女性的阴道中,或者存在于阴道生态失调的女性中,例如细菌性阴道病,甚至在接受抗菌治疗的女性中。
▷乳杆菌的丰度与阴道健康有关
许多研究报告说,卷曲乳杆菌(L. crispatus)在阴道中的存在与身体健康有关,而以惰性乳杆菌(L. iners)为主的群落无法提供足够的保护以防止阴道生态失调。
惰性乳杆菌与炎症反应有关
惰性乳杆菌的存在与较高水平的促炎因子有关,例如白细胞介素-1α、白细胞介素-18、巨噬细胞迁移抑制因子和肿瘤坏死因子,它们负责激活阴道中的炎症反应。
惰性乳杆菌甚至被认为在阴道生态失调中起作用,尽管惰性乳杆菌的确切作用仍有争议。然而,惰性乳杆菌的丰度似乎保持相对恒定,惰性乳杆菌不易被病原体或感染条件取代。
惰性乳杆菌在各种条件下的卓越生存能力表明该物种可能是宿主防御的重要成员,并且可能是一种持久的共生乳杆菌可以维持和恢复阴道微生物组的物种。
细菌性阴道病是育龄妇女最常见的阴道炎类型。它的特点是乳杆菌种类显著减少或消失,伴随着以厌氧和兼性厌氧细菌为主的更多样化的微生物群的出现,例如加德纳菌属、普雷沃氏菌属和阴道曲霉。
注:惰性乳杆菌通常是唯一可以在细菌性阴道病期间检测到的与细菌性阴道病相关细菌共存的阴道乳杆菌。由于其能够响应和调节其基因组功能,它可以在细菌性阴道病急剧变化的阴道环境下持续存在。
▷惰性乳杆菌作为微生物指标
由于惰性乳杆菌在细菌性阴道病中共存,惰性乳杆菌的流行可以作为预测细菌性阴道病发病或中期状态的微生物指标。
此外,惰性乳杆菌对甲硝唑具有抗性,即使在用甲硝唑处理后也是阴道中主要的乳杆菌属物种。惰性乳杆菌显示出更强的竞争优势并共存于被破坏的微生物组中。
因此,惰性乳杆菌的持久性可能会导致长期的阴道生态失调,尤其是在重复治疗周期后。需要进一步的研究来阐明该物种是否只是阴道微生物群转变的生物标志物或细菌性阴道病的促成因素。
尽管如此,最近的一项研究发现了一种名为副格氏乳杆菌(Lactobacillus paragasseri)的人类肠道菌株产生的三种细菌素活性肽。
这些细菌素对惰性乳杆菌具有很强的选择性抑制活性,而卷曲乳杆菌(L. crispatus)、加氏乳杆菌(L. gasseri)和詹氏乳杆菌(L. jensenii)仅受到轻微抑制,表明这些乳杆菌衍生的惰性乳杆菌有效抑制剂可以与甲硝唑联合使用,以改善目前的治疗
▷抗生素和乳杆菌组合可显著降低复发率
传统的治疗方法是使用甲硝唑等抗生素。事实上,口服甲硝唑治疗后的复发率非常高,全身使用抗生素有很大的副作用。在这种情况下,急需新的治疗策略。
益生菌的使用可以改善阴道菌群,增加有益菌,减少有害菌的数量,进一步维持阴道菌群环境的稳定。如今,越来越多的证据表明益生菌可有效治疗 细菌性阴道病。
在对30项研究的荟萃分析中,发现益生菌干预降低了复发率,提高了治愈率。
在另一项荟萃分析中,发现与单独使用抗生素相比,抗生素和益生菌的组合使用可显著降低细菌性阴道病的复发率。
乳杆菌在治疗细菌性阴道病的临床研究


Mei Z,et al.Front Cell Infect Microbiol.2022
生物膜是紧密附着在细菌表面的结构,已知它们比一般细胞对宿主免疫反应和抗生素治疗更具抵抗力。
▷生物膜与阴道感染相关
已经表明,阴道上皮细胞上的生物膜形成与阴道感染密切相关。有足够的证据表明细菌性阴道病与多种微生物生物膜的存在有关。
据推测,加德纳菌属(Gardnerella spp.)启动了生物膜的形成,这支持其他细菌性阴道病相关细菌(BVAB)附着在阴道上皮上,进一步增强了生物膜厚度。
此外,加德纳菌生物膜是抗生素的屏障,通过阻止抗生素的渗透来保护其他其他细菌性阴道病相关细菌。人们普遍认为,细菌性阴道病的高复发率是由于生物膜的形成,生物膜可以保护细菌免受抗生素治疗。
▷乳杆菌可以在一定程度上清除生物膜
先前的研究使用了乳杆菌,试图清除多微生物生物膜,并抑制人类阴道中的感染。
据报道,植物乳杆菌可显著降低HT-29细胞系中大肠杆菌、鼠伤寒沙门氏菌、金黄色葡萄球菌和铜绿假单胞菌的粘附,这使其成为治疗细菌性阴道病的潜在抗生物膜剂。
卷曲乳杆菌(L.crispatus)大大降低了来自健康女性和患有细菌性阴道病女性的阴道加特纳菌(G.vaginalis)对宫颈上皮细胞的粘附。
需要注意的是,惰性乳酸杆菌(L. iners)显著降低了健康女性阴道加特纳菌的粘附力,但增强了致病性阴道加特纳菌的粘附。表明惰性乳酸杆菌可以与细菌性阴道病相关的加特纳菌共存,并可能有助于阴道加特纳菌主导的生物膜形成。
进一步了解阴道共生乳杆菌与生物膜的结构和功能之间的相互作用,对于确定生物膜相关感染的新治疗方法至关重要。
宫颈癌是女性生殖道的恶性肿瘤,每年约有30万人死于宫颈癌。
▷感染人乳头瘤病毒后破坏阴道微生态
感染人乳头瘤病毒(HPV)后,可破坏阴道微生态平衡,使乳杆菌数量减少,增加异常菌群的粘附定植。这进一步导致人乳头瘤病毒蛋白表达上调,促进宫颈上皮内瘤变(CIN)的发展,甚至导致宫颈癌的发生。
研究发现HPV阳性女性的阴道细菌多样性更为复杂,阴道微生物群的组成也不同。持续的高危型HPV感染和宫颈微环境的改变加快了宫颈癌前病变的发生发展。
▷乳杆菌对抵抗宫颈癌具有抵抗作用
一项研究首次证明口服卷曲乳杆菌(Lactobacillus curlicus)可以改变阴道群落状态类型并增加HPV清除率。
乳杆菌作为一种阴道益生菌,不仅可以酸化阴道环境,稳定阴道菌群,增强阴道上皮细胞功能,还可以杀灭宫颈癌细胞。增加益生菌摄入量与减缓癌症进展之间存在重要联系。
乳酸菌激活免疫系统,通过分泌多种抗肿瘤代谢物,包括磷酸化多糖和细胞外多糖,抑制恶性肿瘤的增殖。
抑制癌细胞增殖
乳杆菌吸附并占据阴道上皮,防止引起恶性肿瘤的侵袭性病原菌的粘附。乳酸菌可以通过分泌肽聚糖和胞外多糖来抑制癌细胞增殖。
促进细胞因子产生
乳杆菌主要是增强机体的免疫过程,促进细胞因子的产生,抑制单核细胞的增殖。最近的研究表明,乳杆菌如干酪乳杆菌和鼠李糖乳杆菌通过激活自然杀伤细胞和树突状细胞的成熟发挥抗癌作用。
影响体液免疫和细胞免疫
乳杆菌还能影响细胞免疫和体液免疫,促进胸腺源性细胞的增殖和分化,进一步促进骨髓源性细胞的免疫识别和增殖。
对宫颈癌细胞具有细胞毒性
此外,乳杆菌代谢物对宫颈癌细胞也有细胞毒作用。乳酸杆菌的增加属与高危亚型HPV感染、宫颈上皮内瘤变和癌检出率下降有关。
微生物群在癌症的治疗中发挥着越来越重要的作用。乳酸菌作为一种很有前途的非化疗替代疗法,在恢复和维持正常阴道菌群和治疗宫颈癌方面引起了广泛关注。下面总结了一些乳杆菌对宫颈癌细胞的作用:
乳杆菌在宫颈癌中的作用研究


编辑
Mei Z,et al.Front Cell Infect Microbiol.2022
月经周期是扰乱阴道微生物组多样性的最重要因素之一。卷曲乳杆菌(L.crispatus)通常在育龄妇女的阴道中占据主导地位,而惰性乳杆菌(L.iners)在月经周期期间过度生长并取代卷曲乳杆菌。
▷月经期间阴道微生物变化显著
最近的一项研究报告说,惰性乳杆菌(L.iners)是卵泡期最常见的微生物;惰性乳杆菌和CST IV类型(微生物多样性)在排卵期占主导地位;在黄体期,最常见的类型也是CST IV。
事实上,惰性乳杆菌的丰度在月经期间显著增加,通常与阴道加特纳菌或阴道阿托波氏菌(Atopobium vaginae)的增加有关;然而,在没有干预的情况下,它们随后会在月经后减少。
▷感染人乳头瘤病毒后破坏阴道微生态
由于阴道微生物的动态变化,在同一个体月经周期的不同时间具有不同特征,因此月经周期的采样时刻对于阴道群落分析非常重要。
在怀孕期间保持阴道微生物群中乳杆菌的自然和健康平衡尤为重要。早期的研究证实,怀孕期间阴道中的高雌二醇水平和随之而来的高糖原水平会导致阴道酸化更强,从而随着妊娠的进展促进乳杆菌属的流行。
▷怀孕期间阴道微生物可能存在失调
许多研究表明,以惰性乳杆菌(L.iners)为主的阴道微生物组在怀孕期间更有可能转向生态失调。研究发现,在健康孕妇的孕中期和孕晚期,惰性乳杆菌的丰度显著下降,而卷曲乳杆菌(L.crispatus)的丰度在孕中期与孕早期相比有所增加。
▷乳杆菌丰度与早产直接存在关联
越来越多的证据表明细菌性阴道病是导致不良妊娠结局的主要原因之一,尤其是早产。惰性乳杆菌占主导地位的阴道微生物组,被推测是早产的危险因素。
怀孕早期健康女性的阴道涂片中检测到的惰性乳杆菌可能与早产有关。一项研究报告称,在妊娠16周时,由惰性乳杆菌主导的阴道微生物组是短宫颈和早产(<34周)的一个风险因素。
在更具种族多样性的队列中,卷曲乳杆菌的主导地位对早产具有保护作用。来自不同国家的最新研究也表明,乳杆菌与早产发病率增加之间存在显著关联。


先前的研究报告称,在体外受精辅助生殖失败的患者中,高达40%的患者有异常的生殖道微生物组。
阴道生态失调,包括pH值升高、菌群多样性增加、细菌性阴道病、外阴阴道念珠菌病和滴虫性阴道炎,被认为是不孕的危险因素。
▷惰性乳杆菌丰度与不孕相关
一项研究报告称,惰性乳杆菌的丰度与不孕率的增加有关。最近还报道了以惰性乳杆菌为主的阴道微生物组与输卵管性不孕症和沙眼衣原体感染有关。人们认为,以惰性乳杆菌(L. iners)为主的阴道微生物组是怀孕的不利因素。
乳杆菌属作为人体数量最多的细菌之一,不仅仅只是影响了肠道和阴道健康,在全身其他疾病中也有一定作用。
特应性皮炎(AD)是一种慢性炎症性皮肤病,患者经常会因并发过敏性疾病而出现并发症。
尽管特应性皮炎的发病机制尚不清楚,但数十年的研究表明,特应性皮炎的发病机制可能与遗传因素、环境暴露、皮肤屏障受损、免疫功能异常和微生物失衡有关。
•乳杆菌在预防和治疗特应性皮炎中发挥作用
肠道在免疫反应中起着重要作用。乳杆菌是应用最广泛的益生菌。已经研究了几种乳杆菌用特应性皮炎预防和治疗。
乳杆菌通过对致病菌的竞争性排斥和抗菌活性,刺激先天免疫,促进微生物群落平衡。
据报道,这些乳杆菌可以产生多种物质,如有机酸、过氧化氢、低分子量抗菌剂、细菌素和粘附抑制剂。
乳杆菌的给药降低了免疫球蛋白E(IgE)的血清水平,并实现了Th1/Th2的平衡。乳杆菌加速免疫系统的成熟,维持肠道稳态,改善肠道微生物群,最终改善特应性皮炎症状。
用于治疗和预防特应性皮炎的乳杆菌


Xie A,et al.Front Cell Infect Microbiol.2023
肠道屏障、免疫功能和皮肤屏障在给予乳杆菌后得到了改善。下面列出了一些乳杆菌的作用机制。乳杆菌对患有特应性皮炎的动物和人类都显示出一定的作用。
乳杆菌治疗特应性皮炎的作用


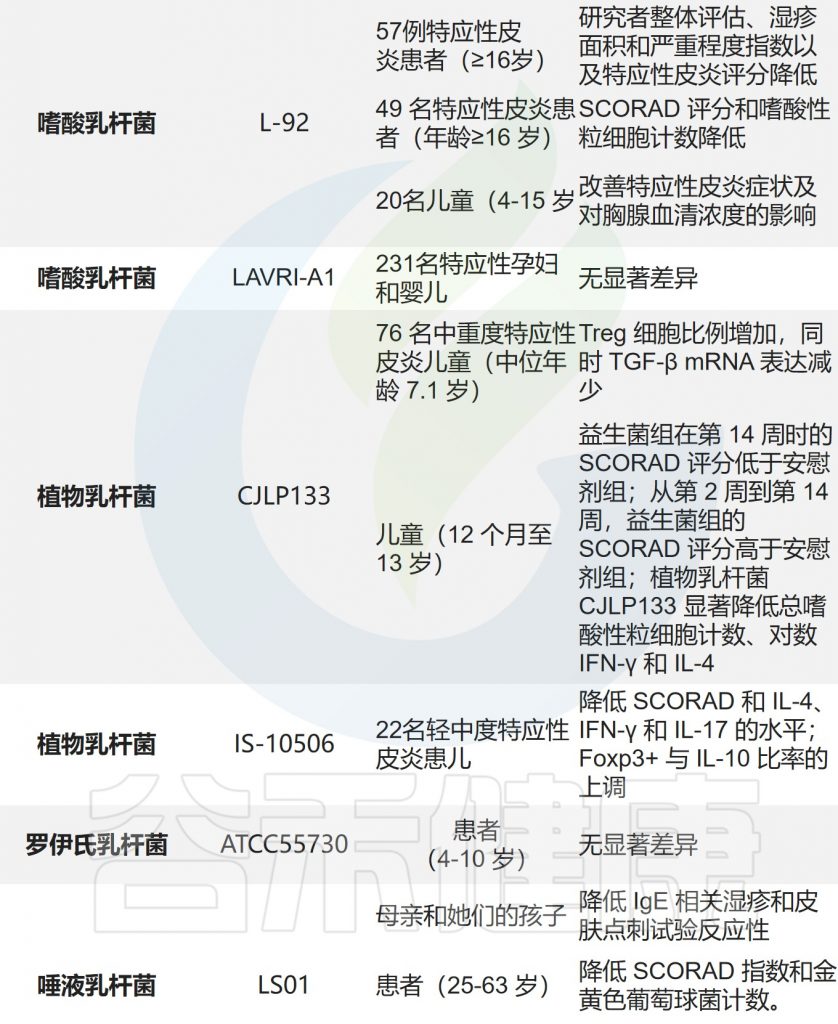



编辑
Xie A,et al.Front Cell Infect Microbiol.2023
肠道微生物群在调节影响全身健康的宿主代谢中起着关键作用。迄今为止,多项研究证实了微生物群与宿主相互作用、调节免疫、控制稳态环境和维持全身状况。
据报道,植物乳杆菌(L. plantarum)具有将亚油酸转化为共轭亚油酸的潜力。根据分子和化学结构,乳杆菌通过多不饱和脂肪酸过程产生的代谢物为10-羟基-顺式-12-十八烯酸(HYA)、10-羟基-十八烯酸(HYB)、10-羟基-反式-11-十八烯酸 (HYC)、10-氧代-顺式-12-十八碳烯酸(KetoA)、10-氧代-十八烷酸(KetoB)和10-氧代-反式 -11-十八碳烯酸 KetoC )。
•乳杆菌代谢物有益于牙周稳态
乳杆菌衍生的生物活性代谢物能带来牙周稳态。HYA和KetoC具有抗氧化、抗炎、抗菌和上皮屏障连接改善剂的作用。
乳杆菌的生物活性代谢物有助于牙周稳态
Sulijaya B,et al.Molecules.2020
• 乳杆菌也可能引起龋齿
虽然链球菌家族细菌(例如变形链球菌)是蛀牙的最常见原因,其他种类的微生物也会引起龋齿。例如,一些乳杆菌属物种与龋齿病例有关。乳酸具有腐蚀牙齿的能力,唾液中的乳杆菌数多年来一直被用作“龋齿测试”。
这是支持在牙膏中使用氟化物的论据之一。乳杆菌的特征是导致现有的龋齿病变进展,尤其是那些在冠状龋齿中的病变。
然而,这个问题很复杂,因为最近的研究表明益生菌可以让有益的乳酸杆菌在牙齿上生长,防止链球菌病原体占据并导致蛀牙。
乳酸菌与口腔健康相关的科学研究是一个新领域,目前发表的研究和结果还很少,需要更多研究。
呼吸道感染是世界上发病率和死亡率最高的疾病之一。尽管许多呼吸道感染具有轻微和自限性,但它们每年在全世界造成400万人死亡。
最近几年严重急性呼吸系统综合症冠状病毒2(也称新型冠状病毒) 在全球肆虐。许多呼吸道病原体没有有效的疫苗,耐药微生物的增加使得呼吸道感染的有效治疗极具挑战性。因此,找到一种安全有效的方法来降低呼吸道感染的风险是很重要的。
•乳杆菌通过肠肺轴影响呼吸系统健康
近年来,许多研究报道肠和肺之间存在串扰,这种被描述为肠-肺轴的联系似乎是双向的 。口服一些益生菌,尤其是乳杆菌,可以通过肠肺轴促进呼吸系统健康。
乳酸杆菌通过肠肺轴调节呼吸免疫的潜在机制

Du T,et al.Front Immunol.2022
最近,乳杆菌已被用于对抗各种呼吸道感染,包括病毒和细菌感染。许多乳杆菌显示可以预防流感病毒感染,包括以下菌株:
L. rhamnosus GG、
L.casei Shirota、
L. plantarum DK119、
L. paracasei MCC1849、
L. gasseri SBT2055、
L. fermentum CJl-112、
L. kunkeei YB38。
此外,乳酸菌由于其安全性和生物技术优势,作为疫苗或佐剂在预防流感病毒感染方面也具有出色的作用。
值得注意的是,补充相同乳杆菌菌株(如鼠李糖乳杆菌CRL1505)的活菌和灭活菌通常具有相似的效果, 表明活力对于乳酸菌达到保护性免疫调节作用不是必需的。
下面是一些通过施用乳杆菌来预防细菌或病毒性呼吸道感染的研究:
乳杆菌预防细菌和病毒性呼吸道感染的研究
Du T,et al.Front Immunol.2022
系统性红斑狼疮是一种慢性自身免疫性疾病,影响大约一半患者的肾脏。狼疮性肾炎(LN)是系统性红斑狼疮发病和死亡的重要危险因素。
狼疮性肾炎与肠壁充血、代谢性酸中毒、频繁使用抗生素有关,所有这些都会对肠道紧密连接产生影响,并导致穿过肠道屏障的细菌代谢产物增加。此外,胃肠道尿素输出增加导致肠道微生物失调。
•乳杆菌混合物有助于恢复黏膜屏障减少肾脏病变
研究发现,5种乳杆菌的混合物(Lactobacillus oris、Lactobacillus rhamnosus、Lactobacillus reuteri、Lactobacillus johnsonii和Lactobacillus gasseri) 通过增加调节性T细胞和抑制致病性Th17细胞来恢复粘膜屏障功能并减少肾脏病变。
表明乳杆菌和乳杆菌衍生的生物活性代谢物在治疗系统性红斑狼疮中具有潜在有益功能。
乳杆菌及其代谢物在系统性红斑狼疮中的有益作用


Wang W,et al.Front Immunol.2022
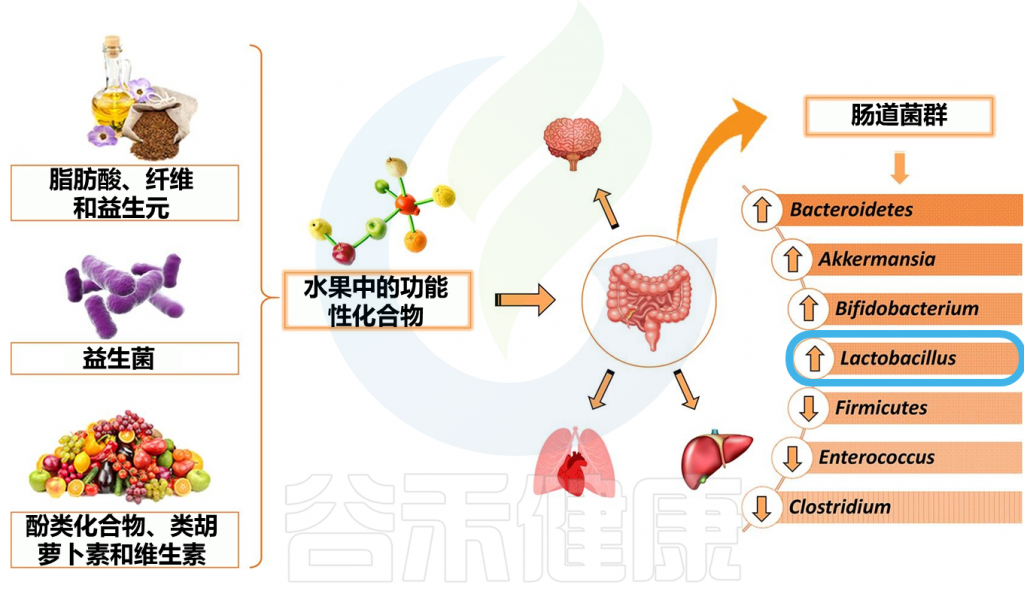

doi.org/10.1016/j.foodres.2022.111809
乳杆菌在人体健康以及一些疾病中起着重要的作用,作为人体不可缺少的一种细菌,我们应该如何补充呢?以下是一些补充乳杆菌的方法:
常见水果
水果中的生物活性物质,包括如胡萝卜素、多酚化合物、维生素和纤维素等,富含这些生物活性物质的水果如香蕉、木莓(树莓)、苹果、芒果、柠檬、石榴、木瓜、猕猴桃等,可以促进乳杆菌属的生长。


食用发酵食品
酸奶、奶酪、酸菜、泡菜、开菲尔、味噌、豆豉等发酵食品中含有丰富的乳杆菌,可以通过食用这些食品来补充乳杆菌。
益生菌
市面上有很多乳杆菌制剂,如乳杆菌素、乳酸菌片等,可以通过口服来补充乳杆菌。
由于补充剂含有不同数量的细菌,因此请阅读包装上的说明。根据补充剂中微生物的数量适量补充。
如果正在服用抗生素,请在抗生素服用2小时之前或者2小时之后服用乳杆菌制剂。
益生元
益生元是一种可以促进肠道内有益菌生长的物质,可以通过食用含有益生元的食品来促进乳杆菌的生长。例如低聚半乳糖(GOS)、低聚果糖(FOS)和母乳低聚糖(HMO)等。
从牛蒡中提取的菊粉促进了有益细菌的生长。菊粉显着增加了小鼠中的乳酸杆菌和双歧杆菌。
将龙舌兰中菊粉等果聚糖添加到正常小鼠饮食中,发现小鼠体内乳酸菌、双歧杆菌数量增加。
全麦谷物可以增加双歧杆菌和乳酸杆菌的相对数量。
一项临床试验(NCT02227602),10名参与者连续8周每天服用200-400克芒果果肉,显著增加乳杆菌、植物乳杆菌、罗伊氏乳杆菌和乳酸乳杆菌的丰度。
还包括其他一些常见食物,例如大蒜、洋葱、韭菜、芦笋、全谷物等,都有助于促进肠道中乳杆菌的生长。


合生元
一项临床试验(NCT 编号:03123510),20 名受试者在接受合生元3个月后,乳杆菌和双歧杆菌显著增加。
注:该研究中的合生元:益生菌成分含有嗜酸乳杆菌DDS-1、乳酸双歧杆菌UABla-12、长双歧杆菌UABl-14和两歧双歧杆菌UABb-10的专利菌株的混合物,益生元成分是一种反式低聚半乳糖混合物。
其他补充剂
乳清和豌豆蛋白的摄入可以增加双歧杆菌和乳酸杆菌的丰度。
灵芝孢子油具有很强的免疫增强活性,有助于肠乳杆菌丰度升高。
绿茶与异麦芽低聚糖联合使用,通过预防小鼠肠道生态失调来对抗高脂肪饮食诱导的代谢改变,双歧杆菌、乳杆菌、罗氏菌属丰度升高。
补充维生素A、维生素C都可以使乳酸杆菌增加。
姜黄素、白藜芦醇、杏仁/杏仁皮可以增加乳酸杆菌。
膳食褐藻糖胶可以增加乳酸杆菌和瘤胃球菌科的丰度。
补充2周葡萄多酚显著提高了断奶后小鼠的乳酸杆菌和Akkermansia的丰度。


中医
对溃疡性结肠炎的患者应用益阳愈溃汤结合针刺治疗,对照组仅给予益阳愈溃汤,治疗后实验组双歧杆菌、乳酸杆菌等均高于对照组。
在连续给药3周后,黄芩促进大鼠结肠的乳酸菌的生长。栀子对乳酸菌双向调节。
黄柏对大鼠肠道乳酸菌的影响:整体表现为促进。
黄连提取物显著促进乳酸菌、双歧杆菌的生长。
吴茱萸碱(EVO)是从吴茱萸中分离出的喹诺酮类生物碱,通过增加嗜酸乳杆菌水平和乙酸盐产生对溃疡性结肠炎具有治疗效果。
山药多糖灌胃健康小鼠一段时间后,发现小鼠盲结肠内的双歧杆菌、乳酸菌均增殖。
马齿苋多糖可使衰老小鼠肠道双歧杆菌及乳酸杆菌数量增加。
蒲公英多糖、黄芪多糖能改善小鼠菌群失调,显著增加双歧杆菌和乳酸杆菌数量。
麦冬多糖 MDG-1 对膳食诱导型肥胖小鼠肠道益生菌群有增殖作用,尤其是一些鼠乳杆菌和台湾乳杆菌。
魔芋低聚糖对三硝基苯磺酸 (TNBS) 诱导溃疡性结肠炎大鼠有保护机制,结肠内乳酸杆菌和双歧杆菌数量增多。
泰山蛹虫草多糖对环磷酰胺 (CY) 免疫抑制小鼠的作用,发现多糖组双歧杆菌、乳酸菌数量均较 CY 模型组增加。
七味白术散能促进小鼠肠道乳酸菌和双歧杆菌的增殖(P<0.05),恢复肠道菌群平衡。
肺癌患者放射治疗后服用养阴清肺汤,治疗12周后,乳酸菌和双歧杆菌含量较对照组均上升(P<0.05)。


辣椒素可降低2型糖尿病小鼠体内乳酸杆菌的丰度。
高盐饮食会使乳酸杆菌的丰度降低。
一些疾病的存在可能与乳杆菌降低相关:
包括阿尔茨海默症、自闭症、炎症性肠病、慢性肝病、过敏疾病、肿瘤、肥胖、牙周病、免疫缺陷病、干燥综合征、系统性红斑狼疮、类风湿关节炎、肾脏疾病等。
注意
虽然乳杆菌在大部分情况下对人体都是有益的,但是过量补充也是不可取的。那么我们如何知道自己体内的乳杆菌含量以及补充是否足够呢?
✔肠道菌群检测可以直观地反映体内乳杆菌丰度
肠道菌群检测是目前较为直观地反映补充剂和食物对肠道乳杆菌影响的方法,通过使用高通量测序技术评估肠道中乳杆菌的菌群丰度。
如果通过检测发现乳杆菌处于正常水平,则不需要额外补充,过高的乳杆菌水平并不一定代表更健康;如果发现乳杆菌缺乏,那么可以通过前文所讲的方法进行补充,有助于营造更健康的身体。
补充乳杆菌并不是一劳永逸的事情,需要长期坚持。同时,对于某些人群,如免疫力低下、肠道疾病患者等,应在医生的指导下进行补充。
总之,乳杆菌是一类非常重要的益生菌,它们能够帮助我们维护肠道和阴道的健康、提高免疫力、改善消化功能,并且可有效治疗各种疾病,包括细菌性阴道病、特应性皮炎和呼吸道感染,对人体健康有着重要的作用。
我们可以通过食用富含乳杆菌的食物或者补充乳杆菌制剂来增加体内乳杆菌的数量,从而更好地保护我们的健康。
此外,乳杆菌对于食品、化工业、保健品和医药行业也都具有重要价值。有必要进一步研究乳杆菌,在未来发掘它更深层的价值。
附录一:乳杆菌种类
· Lactobacillus acetotolerans
· Lactobacillus acidifarinae
· Lactobacillus acidipiscis
· Lactobacillus acidophilus
· Lactobacillus agilis
· Lactobacillus algidus
· Lactobacillus alimentarius
· Lactobacillus alvei
· Lactobacillus alvi
· Lactobacillus amylolyticus
· Lactobacillus amylophilus
· Lactobacillus amylotrophicus
· Lactobacillus amylovorus
· Lactobacillus animalis
· Lactobacillus animata
· Lactobacillus antri
· Lactobacillus apinorum
· Lactobacillus apis
· Lactobacillus apodemi
· Lactobacillus aquaticus
· Lactobacillus aviarius
· Lactobacillus backii
· Lactobacillus bifermentans
· Lactobacillus bombi
· Lactobacillus bombicola
· Lactobacillus brantae
· Lactobacillus brevis
· Lactobacillus brevisimilis
· Lactobacillus buchneri
· Lactobacillus cacaonum
· Lactobacillus camelliae
· Lactobacillus capillatus
· Lactobacillus casei group
· Lactobacillus catenefornis
· Lactobacillus ceti
· Lactobacillus coleohominis
· Lactobacillus collinoides
· Lactobacillus composti
· Lactobacillus concavus
· Lactobacillus coryniformis
· Lactobacillus crispatus
· Lactobacillus crustorum
· Lactobacillus curieae
· Lactobacillus curvatus
· Lactobacillus delbrueckii
· Lactobacillus dextrinicus
· Lactobacillus diolivorans
· Lactobacillus equi
· Lactobacillus equicursoris
· Lactobacillus equigenerosi
· Lactobacillus fabifermentans
· Lactobacillus faecis
· Lactobacillus faeni
· Lactobacillus farciminis
· Lactobacillus farraginis
· Lactobacillus fermentum
· Lactobacillus floricola
· Lactobacillus florum
· Lactobacillus formosensis
· Lactobacillus fornicalis
· Lactobacillus fructivorans
· Lactobacillus frumenti
· Lactobacillus fuchuensis
· Lactobacillus furfuricola
· Lactobacillus futsaii
· Lactobacillus gallinarum
· Lactobacillus gasseri
· Lactobacillus gastricus
· Lactobacillus ghanensis
· Lactobacillus gigeriorum
· Lactobacillus ginsenosidimutans
· Lactobacillus gorillae
· Lactobacillus graminis
· Lactobacillus guizhouensis
· Lactobacillus halophilus
· Lactobacillus hammesii
· Lactobacillus hamsteri
· Lactobacillus harbinensis
· Lactobacillus hayakitensis
· Lactobacillus heilongjiangensis
· Lactobacillus helsingborgensis
· Lactobacillus helveticus
· Lactobacillus herbarum
· Lactobacillus heterohiochii
· Lactobacillus hilgardii
· Lactobacillus hokkaidonensis
· Lactobacillus hominis
· Lactobacillus homohiochii
· Lactobacillus hordei
· Lactobacillus iatae
· Lactobacillus iners
· Lactobacillus ingluviei
· Lactobacillus insectis
· Lactobacillus insicii
· Lactobacillus intermedius
· Lactobacillus intestinalis
· Lactobacillus iwatensis
· Lactobacillus japonicus
· Lactobacillus jensenii
· Lactobacillus johnsonii
· Lactobacillus kalixensis
· Lactobacillus kefiranofaciens
· Lactobacillus kefiri
· Lactobacillus kimbladii
· Lactobacillus kimchicus
· Lactobacillus kimchiensis
· Lactobacillus kisonensis
· Lactobacillus kitasatonis
· Lactobacillus koreensis
· Lactobacillus kullabergensis
· Lactobacillus kunkeei
· Lactobacillus larvae
· Lactobacillus leichmannii
· Lactobacillus letivazi
· Lactobacillus lindneri
· Lactobacillus malefermentans
· Lactobacillus mali
· Lactobacillus manihotivorans
· Lactobacillus mellifer
· Lactobacillus mellis
· Lactobacillus melliventris
· Lactobacillus mindensis
· Lactobacillus mixtipabuli
· Lactobacillus mobilis
· Lactobacillus mucosae
· Lactobacillus mudanjiangensis
· Lactobacillus murinus
· Lactobacillus nagelii
· Lactobacillus namurensis
· Lactobacillus nantensis
· Lactobacillus nasuensis
· Lactobacillus nenjiangensis
· Lactobacillus nodensis
· Lactobacillus odoratitofui
· Lactobacillus oeni
· Lactobacillus oligofermentans
· Lactobacillus oris
· Lactobacillus oryzae
· Lactobacillus otakiensis
· Lactobacillus ozensis
· Lactobacillus panis
· Lactobacillus pantheris
· Lactobacillus parabrevis
· Lactobacillus parabuchneri
· Lactobacillus paracasei
· Lactobacillus paracollinoides
· Lactobacillus parafarraginis
· Lactobacillus parakefiri
· Lactobacillus paralimentarius
· Lactobacillus paraplantarum
· Lactobacillus pasteurii
· Lactobacillus paucivorans
· Lactobacillus pentosus
· Lactobacillus perolens
· Lactobacillus plantarum
· Lactobacillus pobuzihii
· Lactobacillus pontis
· Lactobacillus porcinae
· Lactobacillus psittaci
· Lactobacillus rapi
· Lactobacillus rennanquilfy
· Lactobacillus rennini
· Lactobacillus reuteri
· Lactobacillus reuterii
· Lactobacillus rhamnosus
· Lactobacillus rodentium
· Lactobacillus rogosae
· Lactobacillus rossiae
· Lactobacillus ruminis
· Lactobacillus saerimneri
· Lactobacillus sakei
· Lactobacillus salivarius
· Lactobacillus sanfranciscensis
· Lactobacillus saniviri
· Lactobacillus satsumensis
· Lactobacillus secaliphilus
· Lactobacillus selangorensis
· Lactobacillus senioris
· Lactobacillus senmaizukei
· Lactobacillus sharpeae
· Lactobacillus shenzhenensis
· Lactobacillus sicerae
· Lactobacillus silagei
· Lactobacillus siliginis
· Lactobacillus similis
· Lactobacillus songhuajiangensis
· Lactobacillus sp.
· Lactobacillus sp. 66c
· Lactobacillus sp. 7_1_47FAA
· Lactobacillus sp. Akhmro1
· Lactobacillus sp. BL302
· Lactobacillus sp. C30An8
· Lactobacillus sp. C4I18
· Lactobacillus sp. C4I5
· Lactobacillus sp. CR-609S
· Lactobacillus sp. NRCT-KU 1
· Lactobacillus sp. S16
· Lactobacillus sp. TAB-22
· Lactobacillus sp. TAB-26
· Lactobacillus sp. TAB-30
· Lactobacillus sp. Thmro2
· Lactobacillus sp. oral taxon 052
· Lactobacillus sp. oral taxon 461
· Lactobacillus sp.A A18
· Lactobacillus sp.A A21
· Lactobacillus sp.A A25
· Lactobacillus sp.A A29
· Lactobacillus sp.A A35
· Lactobacillus sp.A A44
· Lactobacillus sp.A A45
· Lactobacillus sp.A A48
· Lactobacillus sp.A A49
· Lactobacillus sp.A A85
· Lactobacillus sp.A A96
· Lactobacillus sp.B A100
· Lactobacillus sp.B A101
· Lactobacillus sp.B A102
· Lactobacillus sp.B A103
· Lactobacillus sp.B A12
· Lactobacillus sp.B A13
· Lactobacillus sp.B A14
· Lactobacillus sp.B A16
· Lactobacillus sp.B A19
· Lactobacillus sp.B A20
· Lactobacillus sp.B A23
· Lactobacillus sp.B A31
· Lactobacillus sp.B A33
· Lactobacillus sp.B A34
· Lactobacillus sp.B A36
· Lactobacillus sp.B A37
· Lactobacillus sp.B A41
· Lactobacillus sp.B A42
· Lactobacillus sp.B A52
· Lactobacillus sp.B A53
· Lactobacillus sp.B A64
· Lactobacillus sp.B A65
· Lactobacillus sp.B A76
· Lactobacillus sp.B A78
· Lactobacillus sp.B A81
· Lactobacillus sp.B A86
· Lactobacillus sp.B A87
· Lactobacillus sp.B A89
· Lactobacillus sp.B A90
· Lactobacillus sp.B A91
· Lactobacillus sp.B A95
· Lactobacillus sp.B A97
· Lactobacillus sp.B A98
· Lactobacillus sp.B AB1
· Lactobacillus sp.B CG1
· Lactobacillus sp.B CG3
· Lactobacillus sp.B CG53
· Lactobacillus sp.B CG63
· Lactobacillus sp.B CG71
· Lactobacillus sp.B CG76
· Lactobacillus spicheri
· Lactobacillus sucicola
· Lactobacillus suebicus
· Lactobacillus sunkii
· Lactobacillus taiwanensis
· Lactobacillus thailandensis
· Lactobacillus tucceti
· Lactobacillus ultunensis
· Lactobacillus uvarum
· Lactobacillus vaccinostercus
· Lactobacillus vaginalis
· Lactobacillus vermiforme
· Lactobacillus versmoldensis
· Lactobacillus vini
· Lactobacillus wasatchensis
· Lactobacillus xiangfangensis
· Lactobacillus yonginensis
· Lactobacillus zymae
附录二:参与的KEGG途径
主要参考文献
Chee WJY, Chew SY, Than LTL. Vaginal microbiota and the potential of Lactobacillus derivatives in maintaining vaginal health. Microb Cell Fact. 2020 Nov 7;19(1):203. doi: 10.1186/s12934-020-01464-4. PMID: 33160356; PMCID: PMC7648308.
Zheng N, Guo R, Wang J, Zhou W, Ling Z. Contribution of Lactobacillus iners to Vaginal Health and Diseases: A Systematic Review. Front Cell Infect Microbiol. 2021 Nov 22;11:792787. doi: 10.3389/fcimb.2021.792787. PMID: 34881196; PMCID: PMC8645935.
Dempsey E, Corr SC. Lactobacillus spp. for Gastrointestinal Health: Current and Future Perspectives. Front Immunol. 2022 Apr 6;13:840245. doi: 10.3389/fimmu.2022.840245. PMID: 35464397; PMCID: PMC9019120.
Mei Z, Li D. The role of probiotics in vaginal health. Front Cell Infect Microbiol. 2022 Jul 28;12:963868. doi: 10.3389/fcimb.2022.963868. PMID: 35967876; PMCID: PMC9366906.
Chen CM, Wu CC, Huang CL, Chang MY, Cheng SH, Lin CT, Tsai YC. Lactobacillus plantarum PS128 Promotes Intestinal Motility, Mucin Production, and Serotonin Signaling in Mice. Probiotics Antimicrob Proteins. 2022 Jun;14(3):535-545. doi: 10.1007/s12602-021-09814-3. Epub 2021 Jul 29. PMID: 34327633; PMCID: PMC9076750.
Sulijaya B, Takahashi N, Yamazaki K. Lactobacillus-Derived Bioactive Metabolites for the Regulation of Periodontal Health: Evidences to Clinical Setting. Molecules. 2020 Apr 29;25(9):2088. doi: 10.3390/molecules25092088. PMID: 32365716; PMCID: PMC7248875.
Xie A, Chen A, Chen Y, Luo Z, Jiang S, Chen D, Yu R. Lactobacillus for the treatment and prevention of atopic dermatitis: Clinical and experimental evidence. Front Cell Infect Microbiol. 2023 Feb 16;13:1137275. doi: 10.3389/fcimb.2023.1137275. PMID: 36875529; PMCID: PMC9978199.
Wang W, Fan Y, Wang X. Lactobacillus: Friend or Foe for Systemic Lupus Erythematosus? Front Immunol. 2022 May 23;13:883747. doi: 10.3389/fimmu.2022.883747. PMID: 35677055; PMCID: PMC9168270.
Du T, Lei A, Zhang N, Zhu C. The Beneficial Role of Probiotic Lactobacillus in Respiratory Diseases. Front Immunol. 2022 May 31;13:908010. doi: 10.3389/fimmu.2022.908010. PMID: 35711436; PMCID: PMC9194447.
Sergeev IN, Aljutaily T, Walton G, Huarte E. Effects of Synbiotic Supplement on Human Gut Microbiota, Body Composition and Weight Loss in Obesity. Nutrients. 2020 Jan 15;12(1):222. doi: 10.3390/nu12010222. PMID: 31952249; PMCID: PMC7019807.
Kim H, Venancio VP, Fang C, Dupont AW, Talcott ST, Mertens-Talcott SU. Mango (Mangifera indica L.) polyphenols reduce IL-8, GRO, and GM-SCF plasma levels and increase Lactobacillus species in a pilot study in patients with inflammatory bowel disease. Nutr Res. 2020 Mar;75:85-94. doi: 10.1016/j.nutres.2020.01.002. Epub 2020 Jan 10. PMID: 32109839.
Wang MX, Lin L, Chen YD, Zhong YP, Lin YX, Li P, Tian X, Han B, Xie ZY, Liao QF. Evodiamine has therapeutic efficacy in ulcerative colitis by increasing Lactobacillus acidophilus levels and acetate production. Pharmacol Res. 2020 Sep;159:104978. doi: 10.1016/j.phrs.2020.104978. Epub 2020 May 30. PMID: 32485282.
Lu F, Li Y, Wang X, Hu X, Liao X, Zhang Y. Early-life polyphenol intake promotes Akkermansia growth and increase of host goblet cells in association with the potential synergistic effect of Lactobacillus. Food Res Int. 2021 Nov;149:110648. doi: 10.1016/j.foodres.2021.110648. Epub 2021 Aug 20. PMID: 34600650.
Borgonovi TF, Virgolin LB, Janzantti NS, Casarotti SN, Penna ALB. Fruit bioactive compounds: Effect on lactic acid bacteria and on intestinal microbiota. Food Res Int. 2022 Nov;161:111809. doi: 10.1016/j.foodres.2022.111809. Epub 2022 Aug 27. PMID: 36192952.
朱晨,段学清,段智璇,谢鑫,张其,田维毅.清热解毒中药黄芩和栀子对大鼠不同肠段菌群的影响[J].中华中医药学刊,2023,42(03):52-57.DOI:10.13193/j.issn.1673-7717.2023.03.012.
朱晨,陈瑞,谢鑫,张其,瞿慧琴,段学清,田维毅.黄柏对大鼠不同肠段主要菌群的影响[J].时珍国医国药,2021,32(12):2851-2855.
崔祥,陶金华,江曙,魏晓燕,徐君,钱大玮,段金廒.黄连提取物与肠道菌群的相互作用研究[J].中草药,2018,49(09):2103-2107.
周欣,付志飞,谢燕,王晓明,李楠,张鹏.中药多糖对肠道菌群作用的研究进展[J].中成药,2019,41(03):623-627.
谭周进,吴海,刘富林,蔡莹,蔡光先,张华玲,曾奥.超微七味白术散对肠道微生物及酶活性的影响[J].生态学报,2012,32(21):6856-6863.
潘婷婷,张爱琴.养阴清肺汤对肺癌患者放疗后皮肤损伤和肠道菌群的影响[J].辽宁中医杂志,2023,50(01):97-100.DOI:10.13192/j.issn.1000-1719.2023.01.028.
谷禾健康

消化是人体生命活动的重要组成部分,分解食物提供能量、促进生长发育、参与免疫功能,然而越来越多的人出现了消化不良。
★ 消化不良在人群中很常见
消化不良在全球范围内都是一种常见的疾病,其发病率在不同地区和人群中有所不同。在中国,消化不良的发病率较高,尤其是城市人群中更为常见。据统计,中国城市居民中消化不良的发病率约为20%-30%。
消化不良主要分为器质性消化不良和功能性消化不良(本文主要讲述功能性消化不良)。功能性消化不良其广义上是一种胃肠道多种症状的综合征,主要包括上腹痛或灼热感,餐后饱胀感及早期饱腹感。
随着对功能性消化不良了解的深入,目前研究发现,消化不良病理生理与脑–肠–微生物群轴紊乱、内脏高敏感性、局部低度炎症、胃肠道感染等因素相关。
肠道菌群可以帮助人体消化吸收,同时还可以合成一些对人体有益的物质,在消化过程中扮演着重要角色。肠道菌群作为新的研究切入点,将有助于探索新的改善功能性消化不良的方法。
在了解消化不良前,我们先来看看人体消化的生理过程:
膳食摄入的消化反应

Livovsky DM,et al.Nutrients.2020
在禁食期间,胃肠道会进行周期性的运动,这种运动被称为迁移运动复合体(MMCs)。迁移运动复合体的功能是将残留物从小肠推进到结肠,以便在新的食物进入之前清空肠道。迁移运动复合体包括静止期和强烈的运动和分泌活动期之间的交替,这种刻板模式有助于保持肠道的健康和功能正常。
在进食开始之前,消化系统已经开始做准备工作,例如,预期即将进食的膳食会刺激唾液和胃液的分泌。当食物进入口腔并被吞咽后,消化系统会进入消化间期运动模式。
食物进入胃后,胃壁会收缩并几乎塌陷,这种主动放松被称为胃适应。固体颗粒通过蠕动活动激活胃窦,启动研磨过程,将食物转化为食糜。
餐后阶段胃会逐渐重新收缩,将食糜推入小肠。胃和小肠的活动会适应消化过程的要求,从口腔开始并延伸到末端回肠,食物被消化并随后被吸收。未被吸收的残留物最终到达结肠,成为肠道微生物群的底物,这些微生物会影响宿主的生理和消化功能。
因此,消化系统的正常活动对于人体的健康和营养摄入非常重要。如果消化系统不正常,可能会导致消化问题和其他健康问题。
消化不良是一种常见的临床症状,主要分为器质性消化不良和功能性消化不良。
器质性消化不良是指由某种器官疾病引起的消化不良。一般通过检查,可以明确诊断是由某个器官的病变引起的消化不良症状,例如肝病、胆道疾病、胰腺疾病、糖尿病等。
✦疾病影响消化
这些疾病会影响到消化道内部的消化酶分泌、肠道蠕动、食物吸收等消化过程,导致消化不良症状的出现,如腹胀、腹泻、便秘、恶心、呕吐、胃痛等。

功能性消化不良是最常见的消化系统疾病,大部分消化不良患者都是属于功能性消化不良。功能性消化不良没有明显的器质性病变,但出现了消化不良症状。
由于功能性消化不良患者病情的反复性与迁延不愈,其生活质量明显降低,同时,所带来的社会医疗成本的增加和生产力的下降也不容忽视。
▸ 发病率
在全球范围内,功能性消化不良女性发病率普遍高于男性。不同地区功能性消化不良的患病率也存在明显差异 :西方国家总体较高,约为10%~40%,亚洲国家较低约为5%~30%。
▸高发人群
•饮食不规律的人群:饮食不规律、暴饮暴食、偏食等不良饮食习惯容易导致消化不良。
•长期服用药物的人群:长期服用某些药物,如抗生素、非甾体抗炎药等,可能破坏肠道菌群平衡,导致消化不良。
•精神压力大的人群:精神压力大、情绪波动较大的人群容易出现消化不良症状。
•高龄人群:随着年龄的增长,人体消化功能逐渐下降,高龄人群容易出现消化不良症状。
•婴幼儿:婴幼儿是消化不良的高发人群之一。由于婴幼儿的消化系统尚未完全发育成熟,消化酶的分泌不足,肠道菌群的构成不稳定等因素,使得婴幼儿容易出现消化不良的症状。

▸ 诊断标准与症状
功能性消化不良的诊断标准:
•餐后饱胀感:即在正常量的膳食后感到不舒服;
•早饱:进食后不久即有饱感,以致摄入食物明显减少,无法完成正常量的膳食;
•上腹疼痛或灼痛:上腹胀多发生于餐后,或呈持续性进餐后加重。
注:满足以上一项或多项,并且没有可以解释这些症状的任何结构性疾病(包括上消化道内窥镜检查)的证据;则可以判断为功能性消化不良
根据主要临床症状可大致分为上腹痛综合征和餐后不适综合征两种亚型。
✦持续时间长,并伴有精神症状
在病程中症状也可发生变化,起病多缓慢,经年累月,持续性或反复发作,不少患者有饮食,精神等症状。
早饱和上腹胀常伴有嗳气。恶心、呕吐并不常见,往往发生在胃排空明显延迟的患者,呕吐多为当餐胃内容物。
不少患者同时伴有失眠、焦虑、抑郁、头痛、注意力不集中等精神症状。这些症状在部分患者中与“恐癌”心理有关。
▸ 功能性消化不良和其他胃肠道疾病的关系
伴有肠易激综合征
许多功能性消化不良患者报告伴有肠易激综合征症状,并且这两种症状经常一同出现在更严重的患者中。
注:虽然这两种情况或多或少都与特定的胃肠道症状有关,但没有结构或生化异常可以解释这些症状。
胰腺功能异常
少数但一定人群的功能性消化不良患者存在胰酶异常或胰腺外分泌功能障碍。胰酶异常和胰腺外分泌功能障碍是否直接影响功能性消化不良症状尚不清楚。
肝脏疾病与消化不良直接存在关联
肝脏疾病如肝硬化、肝炎等也可能导致功能性消化不良症状,如腹胀、食欲不振等。这些疾病可能会影响肠道菌群的平衡,导致消化不良。
功能性消化不良的发病机制尚未完全阐明,可能与胃肠运动异常、内脏超敏反应、肠道菌群紊乱、病原微生物感染、遗传因素、社会心理和神经因素、环境因素等多方面调控异常有关。
需要注意的是,肠道菌群数量和紊乱是功能性消化不良的重要病因之一,将在下文中重点讲述。
➤ 1
√功能性消化不良患者胃排空受损
功能性消化不良的发病机制涉及胃调节、胃排空和十二指肠运动的紊乱。
胃排空——食物由胃排入十二指肠的过程称为胃排空。
一项随机、双盲对照研究发现,功能性消化不良患者的症状与胃调节受损之间存在密切关系 。几份报告表明,一些功能性消化不良患者胃排空受损,一项荟萃分析表明,几乎35%的功能性消化不良患者胃排空明显延迟。
√内脏高敏感性影响功能性消化不良
内脏高敏感性是导致功能性消化不良发展的关键病理生理机制。内脏超敏反应可以通过对肠道机械化学刺激的感知增加,这通常表现为疼痛和灼痛感加重。
辣椒素受体(Trpv1)的激活由神经生长因子 (NGF)、热刺激、辣椒素、前列腺素、酸性pH值和炎症介质触发,进一步释放加剧内脏痛觉的神经肽。
那么一般哪些因素会引起或导致人体胃排空受损或内脏高敏感性,常见的原因如下:
1.神经调节失衡:人体内部的神经系统对内脏的感知和调节非常重要。如果神经系统出现失衡,会导致内脏高敏感性,从而引起胃排空受损以及消化不良等症状。
2.饮食不当:食物的种类、质量和摄入量都可能影响胃排空。比如,吃得太快、咀嚼不充分、进食过多或者过少都会导致胃排空受损。
饮食中过多的刺激性食物(如辛辣、油腻的食物)或者过多的咖啡因、酒精等刺激物质,都可能刺激内脏,导致内脏高敏感性。
3.精神压力:长期的精神紧张、焦虑、抑郁等情绪问题,也会影响人体内部神经系统的平衡,进而导致胃排空受损或内脏高敏感性。
4.慢性疾病:一些慢性疾病,如炎症性肠病、胃溃疡等,也会引起胃排空受损以及内脏高敏感性,从而导致消化不良等症状。
5.长期用药:某些药物,如非甾体类抗炎药、抗生素等,长期使用也可能导致内脏高敏感性。
➤ 2
社会心理因素会导致功能性消化不良症状。
√心理对功能性消化不良具有重要影响
一项针对瑞典人群的研究表明,在10年的随访后,焦虑会使患功能性消化不良的风险增加近8倍。在日本,儿童时期的被虐待与功能性消化不良和功能性消化不良症状的严重程度有关。
此外,病理生理学研究表明,社会心理因素和精神障碍可能通过调节大脑中的信号处理在功能性消化不良中发挥作用以及应激激素对痛觉的影响。
社会心理因素和应激激素也会影响胃肠道的其他方面,例如运动、免疫系统激活、渗透性和微生物群。
√功能性消化不良对精神疾病也有反作用
多项研究强调,与健康人相比,功能性消化不良患者的焦虑和抑郁患病率显著增加。这些观察结果表明,精神疾病在功能性消化不良的发病机制中起着重要作用。
另一方面,由于低度肠道炎症中的细胞因子反应,功能性消化不良症状被认为会诱发焦虑或抑郁,这在功能性消化不良患者心理困扰的发展中起着重要作用。
➤ 3
家族史和遗传多态性可能与功能性消化不良相关。许多研究报告了功能性消化不良风险与遗传多态性之间的关联。
遗传多态性是在同一群体中,某个基因座上存在两个或两个以上的等位基因,且等位基因的频率大于0.01的现象。
GNB3 825C>T、SCL6A4 5HTTLPR、CCK-1R 779T>C等基因多态性被认为与功能性消化不良相关。最近的一项荟萃分析发现,GNB3 825C>T中的次要等位基因 (T) 与上腹疼痛综合征亚型的易感性增加有关。
➤ 4
运动不足、睡眠障碍、高脂肪摄入和饮食不规律等生活方式因素与功能性消化不良的病理生理学有关。
√不健康生活方式导致功能性消化不良发病率高
研究发现,睡眠障碍和运动不足与功能性消化不良相关;脂肪摄入过多会加重功能性消化不良的临床症状,不规律的饮食模式也与功能性消化不良相关。
➤ 5
研究表明,沙门氏菌、幽门螺杆菌、空肠弯曲菌、蓝氏贾第鞭毛虫和诺如病毒等病原微生物导致的急性肠胃炎均与功能性消化不良症状有关。
其机制可能与导致胃肠道Cajal间质细胞(ICC)和肌间神经节受损有关,其发生率在功能性消化不良患者约为10%。
√病因微生物感染会加重消化不良症状
最近的一项荟萃分析证实,根除病原体后症状改善比未治疗的对照组效果更好。在持续时间超过4-12周且内镜检查已排除器质性原因的消化不良患者中,成功根除幽门螺杆菌可使症状缓解率提高10%至15%(或至少症状改善)。
➤ 6
来自动物和临床研究的证据表明,肠道菌群在功能性消化不良中起着重要的作用,影响许多致病机制,包括胃肠动力受损、内脏敏感性、免疫激活、粘膜通透性增加和肠脑轴功能改变等。
在人体中,微生物(包括细菌、古细菌、病毒和真菌)的数量远远超过宿主细胞的数量。
微生物组在调节生理功能(包括胃肠运动功能、上皮屏障保护以及肠道和中枢神经系统之间的相互作用)中发挥着重要作用。然而,微生物失调在功能性消化不良患者中非常常见。
•普雷沃氏菌丰度降低
一项研究将功能性消化不良患者的胃液成分与健康对照组进行了比较,报告称与对照组相比,功能性消化不良中普雷沃氏菌属(Prevotella)的频率显著降低。
这种生态失调的原因可能是胃排空延迟,这可能会改变胃的酸度、粘液稠度和部分氧合作用,从而改变胃的细菌定植。
扩展阅读:肠道重要基石菌属——普雷沃氏菌属 Prevotella
•拟杆菌与变形菌比例增加
功能性消化不良患者胃液的微生物群显示拟杆菌与变形菌的比例增加,而未检测到酸杆菌(Acidobacteria)。然而,健康人的胃液中含有酸杆菌,并且拟杆菌与变形菌的比例较低。
•细菌代谢物分泌出现异常
与对照组相比,功能性消化不良患者的胆汁酸阳性胃液样本比例增加更多。由于胆汁酸从十二指肠反流到胃中,生理上发生在胃运动期间,功能性消化不良患者可能会出现胃动力障碍。
物种丰富度的增加表明胃液微生物群的数量和多样性足以使细菌的代谢物和成分影响胃。因此,可能表明肠道的有毒细菌细胞成分,如脂多糖,刺激白细胞产生促炎细胞因子,引发胃部炎症,从而增加粘膜通透性,这可能导致胃肠神经系统功能障碍。
由于脂多糖和胆汁酸会增加粘膜的通透性,因此患者的炎症可能是由含有此类潜在毒性物质的液体回流引起的。
当比较功能性消化不良患者和健康对照组的上消化道微生物组时,功能性消化不良组口腔、食道、胃和十二指肠中的链球菌(Streptococcus)水平较高。
链球菌丰度与上消化道不适呈正相关,表明链球菌与功能性消化不良患者的胃肠道症状之间存在联系。此外,功能性消化不良组表现出更高水平的厚壁菌门。
• 厚壁菌、变形菌丰度增加
根据这些发现,在一项比较功能性消化不良患者和健康受试者十二指肠粘膜微生物群的不同研究中,十二指肠粘膜中最普遍菌属中也有链球菌。
此外,通过分析功能性消化不良和肝郁脾虚综合征大鼠的粪便样本的微生物组成,与对照组相比,该模型中厚壁菌门、变形菌门(Proteobacteria)和蓝藻门(Cyanobacteria)的水平升高,而拟杆菌门的丰度较低。
上述研究表明,功能性消化不良中的微生物改变并不局限于胃肠道中的一个部位,突出了稳态失衡在这些疾病的发病机制中的潜在重要性。
小肠细菌过度生长 (SIFO) 的定义是小肠中存在过量的细菌,并且与胃肠道症状有关。
最近的研究分析了功能性消化不良患者十二指肠的细菌属水平。
链球菌相对丰度增加
澳大利亚的一项研究报道,与对照组相比,功能性消化不良患者的链球菌(Streptococcus)相对丰度增加,尽管不显著(这可能是因为样本量小),并且链球菌的丰度与厌氧属普雷沃氏菌、韦荣球菌(Veillonella)和放线菌(Actinomyces)的丰度之间存在负相关,这些菌群功能性消化不良患者中显著减少。
β多样性发生显著变化
此外,报告的十二指肠的β多样性在患者和对照组之间存在显著差异,而α多样性保持不变,表明该疾病可能涉及更复杂的微生物群结构变化,而不是仅特定属的相对丰度变化。
奈瑟菌和卟啉单胞菌丰度降低
最近的一项研究表明,在使用质子泵抑制剂(PPI)治疗之前,功能性消化不良患者和对照组的十二指肠粘膜奈瑟菌(Neisseria)和卟啉单胞菌(Porphyromonas)丰度降低,但微生物负荷没有差异。
总体而言,研究证实功能性消化不良患者中确实发生了微生物数量和多样性方面的明显变化。
功能性消化不良患者的微生物变化




Brown G,et al.Neurogastroenterol Motil.2022
谷禾还检测了一些功能性消化不良人群的肠道菌群,虽然个体直接存在一定差异,但是共同之处是菌群紊乱,核心菌异常较多,多项病原菌超标。
案例一
基本信息和病症:
1岁3个月,食物过敏,厌食,功能性消化不良,生长发育不良
菌群构成:
重要菌群情况:


注:高亮部分是异常菌属。
总结:菌群失衡,核心菌属异常较多,普雷沃氏菌属(Prevotella)缺乏,多种致病菌超标。
案例二
基本信息和病症:
43岁,每天大便不成形;容易急性肠胃炎;消化不良
菌群构成:
重要菌群情况:
<来源:谷禾健康肠道菌群检测数据库>
总结:菌群失衡,菌群多样性低,核心菌属大量异常,致病菌和病原菌多项超标。
功能性消化不良是一种复杂的多因素导致的功能性胃肠疾病,目前其确切的发病机制尚不明确。
★ 肠道菌群在功能性消化不良中起重要作用
多个研究表明,肠道菌群紊乱在功能性消化不良的发生发展起着重要作用,是功能性消化不良的重要发病机制。影响包括胃肠动力受损、内脏高敏感性、免疫激活、粘膜通透性增加和中枢神经系统疾病。
肠道微生物群参与功能性消化不良发病机制
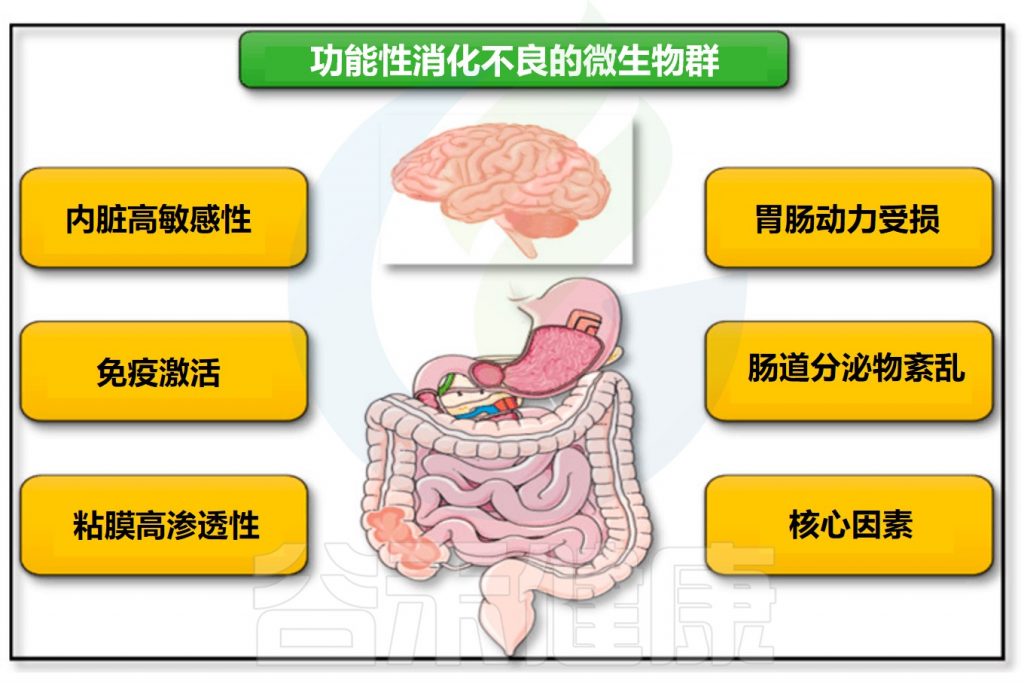

Tziatzios G,et al.Microorganisms.2020
胃肠运动异常是功能性消化不良的基本病理机制,包括胃排空延迟、胃容受性舒张受损及消化期间移行性复合运动(MMC)III期异常。
✦外来菌的增殖抑制胃肠道蠕动
消化期胃和小肠移行性复合运动可将胃肠内容物和致病菌机械性地迁移向远端肠道,而功能性消化不良患者经常出现非传播性和逆传性活动,这可能诱导或加重近端小肠细菌随反流十二指肠液迁移到胃。
而胃排空延迟又导致反流菌能较长时间保留在胃和十二指肠中,引起菌群分布失调的同时,外来菌的增殖可产生内毒素脂多糖刺激免疫应答,进而,抑制胃肠道蠕动,加重功能性消化不良症状。
✦细菌代谢物是胃肠动力的重要来源
短链脂肪酸是肠道共生菌发酵膳食纤维代谢物,除了作为胃肠动力的重要能量来源,也可直接激活肠神经系统以调控肠内分泌细胞合成和分泌某些胃肠激素,如肠激素肽、胆囊收缩素、胰高血糖素样肽等,进而调节胃肠道动力及胃排空。然而在代谢组学研究中发现功能性消化不良大鼠存在短链脂肪酸水平显著降低。
更准确地说,细菌产生的短链脂肪酸不仅调节功能性消化不良中十二指肠碳酸氢盐的分泌,同时它们在十二指肠的快速吸收也可能影响管腔细菌定植抑制。
5-羟色胺影响胃肠道动力
相关研究证实肠道微生物群在调节5-羟色胺(5-HT)合成中起着关键作用,而5-羟色胺水平是影响胃肠道动力的重要因素之一。
大肠杆菌产生的脂多糖延迟胃排空
此外,已发现大肠杆菌(Escherichia coli)产生的细菌脂多糖会导致胃排空显著延迟 ,而双歧杆菌当用作益生菌使用时时可显著增强小肠蠕动。
由上述研究可知,胃肠动力异常可能诱发菌群失调,而菌群失调可反过来进步一步影响胃肠动力,介导功能性消化不良发生发展,但菌群失调与功能性消化不良的因果关系仍需更深入的研究加以验证。
✦心理因素和肠道炎症相互影响
功能性消化不良患者处于十二指肠低度炎症状态。这种慢性低级别的炎症反应会引起疼痛和敏感性。
此外,焦虑或抑郁等心理学因素似乎与慢性炎症反应相互影响,心理压力和由此产生的皮质醇释放激素分泌也会增加十二指肠的局部炎症和全身炎症反应。
✦炎症状态下肠屏障功能发生改变
多项研究报道了功能性消化不良患者肠道中免疫细胞(如肥大细胞、巨噬细胞和嗜酸性粒细胞)数量的增加。
十二指肠活检切片中嗜酸性粒细胞增多

Hari S,et al.Front Allergy.2022
活化的肥大细胞释放细胞因子、组胺、前列腺素和类胰蛋白酶,它们与肠屏障功能障碍和伤害感受通路的改变有关。
嗜酸性粒细胞影响肠道通透性
在生理条件下,十二指肠屏障由粘液层、上皮和下面的固有层组成。在功能性消化不良中,屏障的功能障碍体现为粘膜完整性丧失和通透性增加。
此外,功能性消化不良中嗜酸性粒细胞的募集和激活也发生,这可能使组织损伤和屏障功能障碍持续存在。
正常与功能性消化不良的屏障功能对比
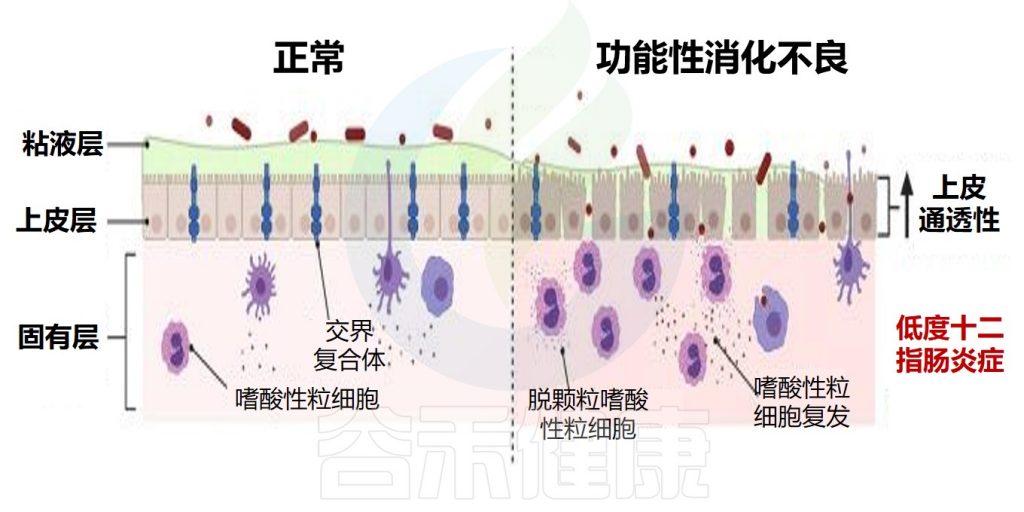

Hari S,et al.Front Allergy.2022
✦肠道菌群也会破坏肠道通透性
肠腔黏膜表面是毒性和免疫原性颗粒面对黏膜相关免疫系统的第一道屏障。肠道微生物菌群失调在功能性消化不良中非常常见。
有害菌破坏紧密连接增加肠道通透性
有研究表明,霍乱弧菌(V.cholera)、艰难梭菌(Clostridium difficile)和产毒素的大肠杆菌的各种菌株已显示可通过直接破坏紧密连接(TJ)、毒素或蛋白酶的产生以及炎症级联反应的活化来增强肠道通透性。
益生菌可以促进屏障完整性
相反,肠道益生菌可以通过增加闭合蛋白、紧密连接蛋白ZO-1和ZO-2的表达来促进屏障完整性。
另外,肠道菌群的某些代谢产物也能够破坏肠道通透性,例如,细菌细胞壁脂多糖易位可以诱导免疫反应和炎症反应,加剧肠道屏障损害并进一步增加肠道的通透性。
扩展阅读:什么是肠漏综合征,它如何影响健康?
除了影响黏膜屏障功能,微生物群也参与调节免疫系统的发育和功能,在先天免疫系统中起关键作用。
✦肠道菌群调节肠上皮免疫功能
研究发现,肠道微生物区系通过激活肠上皮细胞的模式识别受体和内质网应微信号促进免疫系统的发育,从而促进肠上皮细胞的增殖、抗菌肽和黏液的产生,调节与免疫功能相关细胞因子的分泌,如白细胞介素1β、白细胞介素18和白细胞介素25。
✦微生物代谢物也可促进免疫反应
肠道微生物群及其代谢物还通过Toll样受体(TLRs-TLR2、TLR4)干扰信号,并促进促炎细胞因子的产生和免疫反应。
此外,它们的代谢物也可能具有炎症特性或对T细胞分化产生直接影响。
小结
综上所述,肠道菌群失调在影响黏膜生物屏障的同时,可能也导致黏膜机械屏障、化学屏障及免疫屏障等多种屏障功能低下,并可能介导黏膜低度炎症和内脏高敏性的发生。
生物失调的影响
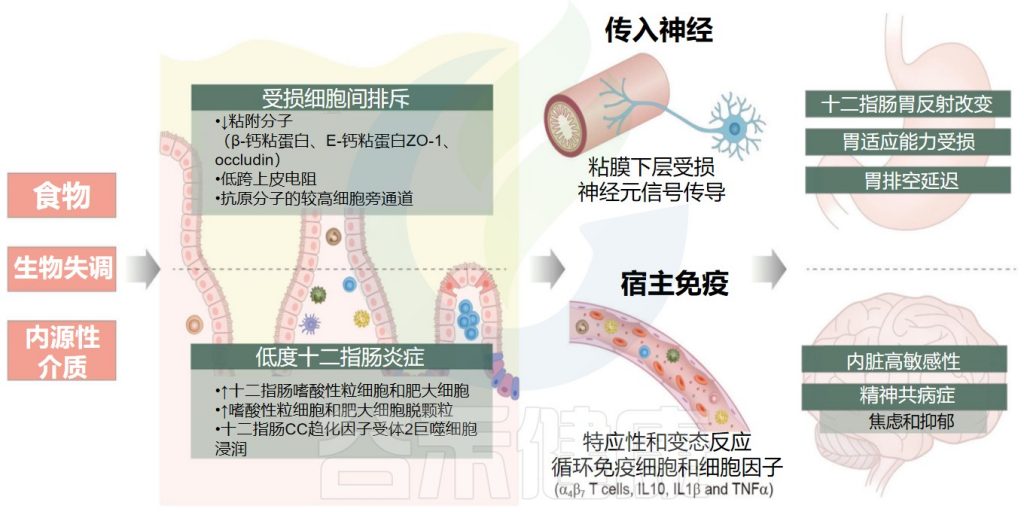

Kim SH.Korean J Gastroenterol.2022
脑-肠轴是连接胃肠道和中枢神经系统的传导通路,包括中枢神经系统、脑和脊髓、自主神经系统、肠神经系统和下丘脑垂体肾上腺轴。
肠道菌群与脑-肠轴存在双向交互作用:
肠道菌群通过神经、内分泌、免疫等至少三种途径与中枢神经系统相通;中枢神经系统调节肠道的运动、分泌及肠道黏膜通透性来影响肠道菌群,或通过肠腔分泌的激素直接调节菌群的基因表达而影响肠道菌群的组成和功能。
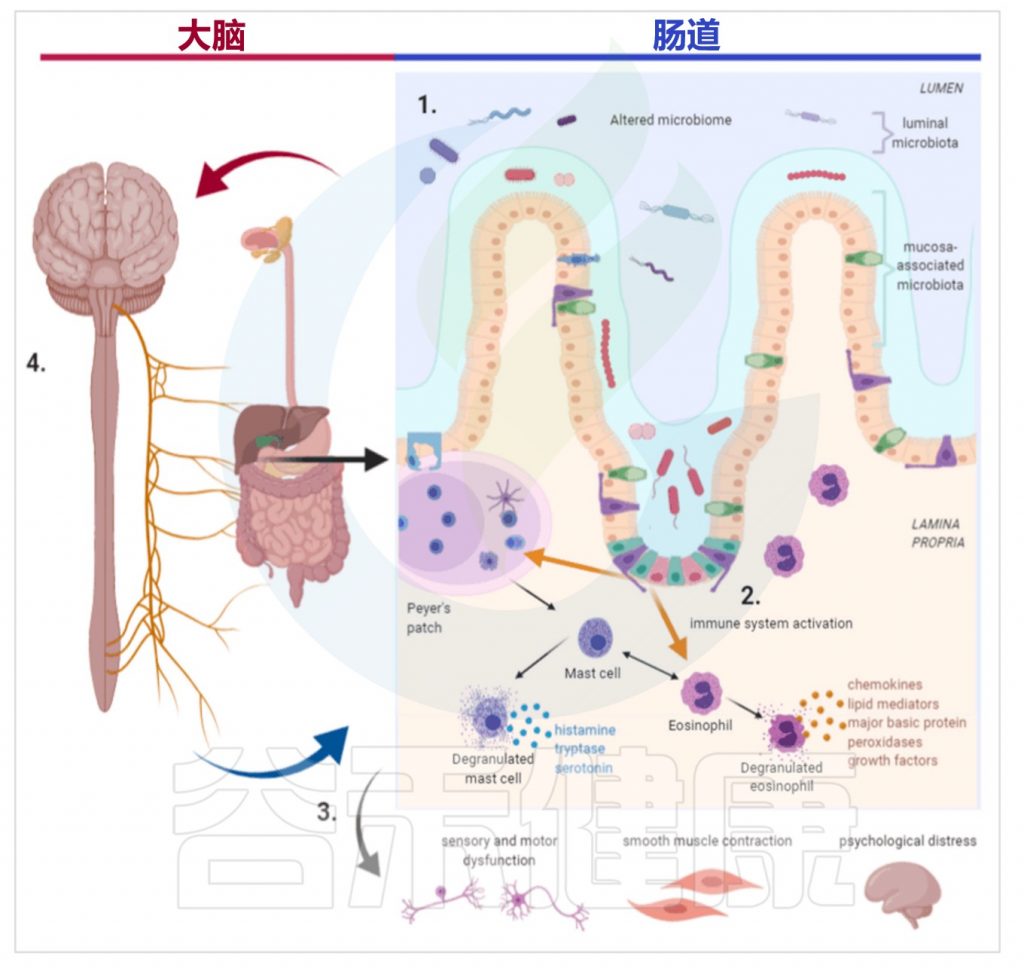

Brown G,et al.Neurogastroenterol Motil.2022
肠道菌群通过肠-脑轴影响5-羟色胺、脑源性神经营养生长因子(BNDF)等神经递质的合成、释放和下丘脑-垂体-肾上腺轴的发育,调节中枢神经系统的发育、功能和行为,影响焦虑、抑郁样情绪障碍和应激反应。
✦功能性消化不良患者伴有精神症状
功能性消化不良属于典型的身心疾病,常有焦虑、抑郁的表现。一项为期12年的前瞻性研究也说明功能性消化不良患者中有1/2~2/3先出现焦虑再有胃肠道症状,提示了功能性消化不良的中枢发病机制。
功能性消化不良患者的中枢神经系统改变
功能性神经影像学,如功能核磁共振,使我们能直接观察到中枢神经系统的改变,证明其与功能性消化不良症状的相关性。
结果显示功能性消化不良患者额叶皮层、感觉皮层、脑岛、前扣带皮层、丘脑、海马体和杏仁核的改变,与功能性消化不良内脏超敏反应、消化不良症状、生活质量及焦虑和抑郁有关。
精神心理应激影响内脏高敏感性
有研究认为功能性消化不良患者在精神心理应激的情况下可通过刺激中枢神经系统的情感活动系统参与患者内脏高敏感性的形成,从而对胃肠道产生影响。
因此,研究人员推测肠道菌群通过脑-肠轴对中枢神经系统的调节,导致消化不良伴焦虑抑郁的发生发展。
✦肠道菌群对肠脑轴具有重要作用
随着研究不断深入,越来越多证据证实肠道菌群与脑-肠-菌群轴功能密切相关。
菌群代谢物影响中枢神经系统信号
一方面,肠道微生物能通过合成和释放短链脂肪酸、次级胆汁酸和色氨酸等代谢物,与肠内分泌细胞,自下而上影响激活内源性中枢神经系统信号机制。
此外,肠道菌群失调会增加脂多糖的产生和分泌,而脂多糖不仅是影响中枢神经系统功能的一个强有力的因素,还会促进其他炎症因子的产生,如肿瘤坏死因子﹣α、白细胞介素﹣1β等,以上炎症因子可影响中枢神经系统的功能, 紊乱神经肽的合成和分泌。
激活肠神经系统信号
另一方面,肠道微生物也可独立完成或促进某些神经活性物质的合成和分泌,如5-羟色胺、γ-氨基丁酸、去甲肾上腺素、多巴胺等,这些小分子物质能通过与肠内分泌细胞相互作用,激活肠神经系统信号机制。
上述研究表明,肠道菌群失调可能诱导脑-肠-微生物群轴功能紊乱,其机制涉及神经、免疫和内分泌信号传导。
✦迷走神经的作用
在生态失调和胃肠道不适的精神状况的背景下,应该注意的一个重要方面是迷走神经的作用。
众所周知,压力会增加肠道通透性并通过各种神经调节剂改变胃肠道微生物组的组成,同时压力可以减少迷走神经活动,这可能会促进胃肠道炎症。
注:虽然目前还没有关于迷走神经刺激对胃肠道微生物组影响的数据,但可以推测迷走神经可能通过其对肠道通透性的影响对肠道微生物组产生影响。
迷走神经的传出可能在肠道中具有抗炎作用,同时降低肠道通透性——这两种作用都可能归因于通过迷走神经活动加强紧密连接。
心理治疗对功能性消化不良患者微生物群和胃肠道症状的影响,也可以用迷走神经的影响来解释。
心理疾病对功能性消化不良的表现有影响
现在人们可能想知道心理疾病的患病率是否在不同的功能性消化不良亚组中有所不同。
已经表明情绪和焦虑障碍在非疼痛主导型功能性消化不良患者中,比在疼痛主导型功能性消化不良患者中更常见。
另一项研究还表明上腹痛与神经质、虐待和躯体化之间存在相关性。
这些发现强调了未来重点关注功能性消化不良患者的心理合并症和特定症状之间的可能联系,研究心理治疗对功能性消化不良患者特别是微生物组的这些有趣影响。
未来随着研究深入,有望更好地理解脑-肠-微生物组轴,功能性消化不良的治疗方法也能更加个性化。
药物经口服途径进入人体,不可避免地与肠道菌群发生相互作用。
多项研究表明,肠道微生物能通过编码多种酶,影响口服药物在体内的吸收、代谢、转化等过程,进而影响药物活性成分的有效性和毒性。
改变微生物群的药物在治疗消化不良中得到了有效证实。
利福昔明是一种广谱抗生素,具有革兰氏阳性、革兰氏阴性、需氧和厌氧覆盖,在胃肠道腔内生物利用度高,全身不良反应极小。
•消化不良症状缓解
在一项随机试验中,利福昔明治疗优于安慰剂治疗,其中79%的患者在服用利福昔明后报告消化不良症状缓解,而安慰剂组在8周时这一比例为47%。
•利于有益细菌生长
在其他胃肠道疾病中,利福昔明治疗已被证明可以保护结肠菌群,增加乳酸杆菌和双歧杆菌(Bifidobacterium)的丰度,从而产生积极效应,利于有益细菌的生长,从而改变整体成分。
扩展阅读:肠道核心菌属——双歧杆菌,你最好拥有它
•益生菌治疗后餐后腹胀改善
益生菌可能在功能性消化不良中发挥治疗作用,三项研究表明,在幽门螺杆菌阳性和阴性消化不良患者中服用益生菌加氏乳杆菌(Lactobacillus gasseri OLL2716)12周后,餐后饱腹感和腹胀症状得到改善。
35.5%的幽门螺杆菌阴性功能性消化不良患者的症状得到了缓解,而安慰剂患者的症状缓解率为17%。
扩展阅读:如果你要补充益生菌 ——益生菌补充、个体化、定植指南
•恢复胃液微生物群
加氏乳杆菌还被证明可以“恢复”功能性消化不良患者异常的胃液微生物群。在服用益生菌12周之前,功能性消化不良患者的胃液显示拟杆菌比变形菌占优势,并且不存在酸杆菌(Acidobacteria),益生菌治疗后,这些比例发生了变化,以反映与健康志愿者相似的微生物群组成。
此外,虽然没有收集小肠细菌数据,但用凝结芽孢杆菌(Bacillus coagulans MY01)和枯草芽孢杆菌(Bacillus subtilis MY02)治疗8周后,功能性消化不良患者的有效率比安慰剂提高了28%。
这些发现提供了初步证据,表明患者症状与肠道微生物群中某些属的丰度有关,这些微生物群可以通过治疗性抗生素和益生菌制剂进行调节。
尽管有这些显著的具体变化,但仍需要对更多不同地理区域的患者群体进行进一步研究,以验证疾病中的微生物组变化。
15项观察性研究和一项随机对照试验的系统回顾发现,高脂肪、小麦、FODMAP(可发酵低聚糖、双糖、单糖和多元醇)和咖啡因等天然食品化学物质含量高的食物与功能性消化不良有关。
扩展阅读:肠道微生物群与健康:探究发酵食品、饮食方式、益生菌和后生元的影响
饮酒与功能性消化不良症状的关联尚不明确,可能取决于酒精的类型。
以下是一些饮食调理建议:
•少食多餐:每天分成5-6餐,每餐食量适中,避免暴饮暴食。
•避免过度饮酒和吸烟:酒精和烟草会刺激胃肠道,加重消化不良症状。
•避免食用刺激性食物:如辛辣、油腻、烧烤等食物,容易引起胃肠道不适。
•增加膳食纤维摄入量:膳食纤维可以促进肠道蠕动,缓解便秘症状。建议多食用蔬菜、水果、全谷类食品等富含膳食纤维的食物。
•注意饮食卫生:避免食用过期食品、生冷食物等容易引起胃肠道感染的食物。
•喝足够的水:保持足够的水分摄入可以促进肠道蠕动,缓解便秘症状。
•食用易消化的食物:如米粥、面条、煮熟的蔬菜等,可以减轻胃肠道负担,缓解消化不良症状。
注意:饮食调理虽然可以缓解功能性消化不良症状,但如果症状持续或加重,建议及时就医进行诊断和治疗。
扩展阅读:20种有效改善肠道健康的科学方法
胃酸抑制的主要药物包括两类药物:质子泵抑制剂(PPI)和组胺H2受体拮抗剂。
•上腹痛和反流症状缓解
一项荟萃分析发现,在两到八周内服用标准剂量的质子泵抑制剂,在减少功能性消化不良症状方面优于安慰剂。
注:治疗益处仅出现在有上腹灼痛和反流样疼痛的患者身上,而不出现在有运动障碍相关问题的患者身上。
•消化不良症状的风险降低
组胺H2受体拮抗剂也是功能性消化不良治疗的一种选择。十二项随机对照试验将H2拮抗剂与安慰剂进行了比较,显示消化不良症状的相对风险降低了23%。
对于以进餐相关症状为特征的功能性消化不良伴餐后窘迫综合征的患者,促胃肠动力药可以减轻症状。
对29项比较促动力药(主要是西沙必利)与安慰剂的试验的回顾发现,功能性消化不良的整体症状显著减少。
对功能性消化不良患者的12项随机对照试验的回顾发现,心理治疗(即认知行为治疗和其他形式的心理治疗)比对照组具有统计学上的显著益处。
认知行为疗法:是一种心理治疗方法,旨在通过调整患者的思维方式和行为习惯,来改善其身体和心理健康问题。在功能性消化不良患者治疗中,认知行为疗法可以帮助患者意识到自己不良的思维和行为习惯,如过度焦虑、负面情绪、饮食习惯不当等,从而通过改变这些不良习惯,减轻和预防消化不良症状的发生。
此外,认知行为疗法还可以帮助患者学会应对压力和情绪问题的技巧,提高其身心健康水平。
•症状严重程度、疼痛强度减弱
认知行为疗法 (CBT)导致症状严重程度、疼痛强度减弱,并进一步降低疾病对患者生活的影响。
•胃动力显著增加
此外,与对照组相比,干预组胃排空率显著增加,胃动力参数发生变化。基于这些结果,认知行为疗法可被视为功能性消化不良患者的有效治疗选择。
注:心理治疗不应作为功能性消化不良的一线治疗,因为数据质量低,缺乏盲法心理干预和主观症状评分导致偏倚风险高。如果药物治疗无效,可以考虑心理治疗。
中药具有多成分、多靶点和多途径的优势,目前中医对功能性消化不良肠道菌群的研究仍处于探索阶段。
中药在治疗功能性消化不良方面具有一定的疗效,可以缓解消化不良症状,改善消化功能,提高生活质量。研究证实,中药能逆转肠道菌群失调,维持肠道微生态平衡。
注:中药治疗功能性消化不良应根据患者的具体情况进行个体化治疗,避免不必要的药物过敏或不良反应。同时,中药治疗应在专业医生的指导下进行。
基于菌群干预后,原先消化不良患者的肠道菌群有了好转:







<来源:谷禾健康肠道菌群检测数据库>
从以上菌群评估报告显示,核心菌属和重要菌属没有异常,整体菌群平衡和多样性良好,健康评分显著提升,检测者反馈消化不良症状大幅减轻。
功能性消化不良是一种常见的消化系统疾病,其症状包括腹胀、腹泻、便秘等,给患者带来很大的困扰和痛苦。目前,传统的治疗方法主要是针对症状进行缓解,但效果并不理想。近年来的研究表明,肠道微生物与功能性消化不良密切相关,肠道菌群干预成为了治疗功能性消化不良的新途径。
谷禾的肠道菌群检测经验和研究表明肠道菌群的紊乱和异常特征可以对部分功能性消化不良患者症状背后的病理进行分类和解析,为针对性干预和改善提供了新的可能。对肠道微生物与功能性消化不良之间的关系的进一步深入研究和应用有望带来更有效的治疗方法。
主要参考文献
Singh R, Zogg H, Ghoshal UC, Ro S. Current Treatment Options and Therapeutic Insights for Gastrointestinal Dysmotility and Functional Gastrointestinal Disorders. Front Pharmacol. 2022 Jan 25;13:808195. doi: 10.3389/fphar.2022.808195. PMID: 35145413; PMCID: PMC8822166.
Martinez-Guryn K, Hubert N, Frazier K, Urlass S, Musch MW, Ojeda P, Pierre JF, Miyoshi J, Sontag TJ, Cham CM, Reardon CA, Leone V, Chang EB. Small Intestine Microbiota Regulate Host Digestive and Absorptive Adaptive Responses to Dietary Lipids. Cell Host Microbe. 2018 Apr 11;23(4):458-469.e5. doi: 10.1016/j.chom.2018.03.011. PMID: 29649441; PMCID: PMC5912695.
Shah A, Talley NJ, Holtmann G. Current and Future Approaches for Diagnosing Small Intestinal Dysbiosis in Patients With Symptoms of Functional Dyspepsia. Front Neurosci. 2022 May 6;16:830356. doi: 10.3389/fnins.2022.830356. PMID: 35600619; PMCID: PMC9121133.
Miwa H, Nagahara A, Asakawa A, Arai M, Oshima T, Kasugai K, Kamada K, Suzuki H, Tanaka F, Tominaga K, Futagami S, Hojo M, Mihara H, Higuchi K, Kusano M, Arisawa T, Kato M, Joh T, Mochida S, Enomoto N, Shimosegawa T, Koike K. Evidence-based clinical practice guidelines for functional dyspepsia 2021. J Gastroenterol. 2022 Feb;57(2):47-61. doi: 10.1007/s00535-021-01843-7. Epub 2022 Jan 21. PMID: 35061057; PMCID: PMC8831363.
Mounsey A, Barzin A, Rietz A. Functional Dyspepsia: Evaluation and Management. Am Fam Physician. 2020 Jan 15;101(2):84-88. PMID: 31939638.
Tziatzios G, Gkolfakis P, Papanikolaou IS, Mathur R, Pimentel M, Giamarellos-Bourboulis EJ, Triantafyllou K. Gut Microbiota Dysbiosis in Functional Dyspepsia. Microorganisms. 2020 May 8;8(5):691. doi: 10.3390/microorganisms8050691. PMID: 32397332; PMCID: PMC7285034.
Brown G, Hoedt EC, Keely S, Shah A, Walker MM, Holtmann G, Talley NJ. Role of the duodenal microbiota in functional dyspepsia. Neurogastroenterol Motil. 2022 Nov;34(11):e14372. doi: 10.1111/nmo.14372. Epub 2022 Apr 11. PMID: 35403776; PMCID: PMC9786680.
Hari S, Burns GL, Hoedt EC, Keely S, Talley NJ. Eosinophils, Hypoxia-Inducible Factors, and Barrier Dysfunction in Functional Dyspepsia. Front Allergy. 2022 May 31;3:851482. doi: 10.3389/falgy.2022.851482. PMID: 35769556; PMCID: PMC9234913.
Rupp SK, Stengel A. Bi-Directionality of the Microbiota-Gut-Brain Axis in Patients With Functional Dyspepsia: Relevance of Psychotherapy and Probiotics. Front Neurosci. 2022 Feb 28;16:844564. doi: 10.3389/fnins.2022.844564. PMID: 35295092; PMCID: PMC8919856.
谷禾健康
播种肠道,喂养心灵
在新冠疫情的影响下,我们的生活方式和社交模式都发生了很大的改变。随着社交距离的要求和封锁措施的实施,我们不得不放弃了很多与朋友和家人的互动,这给我们的身心健康带来了很大的影响。
然而,随着疫情的好转和社交限制的逐渐放松,我们有了更多的机会去重新建立社交联系和友谊。
社会关系对于群居动物(例如我们人类和其他灵长类动物)的健康和福祉至关重要。越来越多的证据表明,肠道微生物组通过所谓的“肠-脑轴”,在我们的身心健康中发挥着关键作用,而且细菌可以通过接触等方式在社交中传播。
本文来了解一下,社会关系与肠道菌群之间的一些有趣的关联。

从动物到人类都有一个微生物组,或者更确切地说,有几个微生物组。消化系统、皮肤和身体的其他部位承载着微生物群落,也就是肠道微生物群、皮肤微生物群等。
在过去的10-15年中,随着DNA测序技术的不断进步,人们有了更清晰的视野来观察微生物组的多样性和复杂性。
一般关于微生物组与社会性之间联系的研究都集中在病原体和感染上。
任何场所,不管是曾经在医院就医,或生过孩子,还是与咳嗽和打喷嚏的人一起乘坐公共交通工具等,微生物都可以通过身体接触或共享环境将疾病从一个人传播到另一个人。
一些研究人员认为,对感染的恐惧可能是人类对陌生人根深蒂固的戒心的一种解释。
越来越多的迹象表明,社会伙伴之间的微生物共享也正在影响其他事情。例如,大黄蜂可能会被一种有害的剧毒寄生虫感染。2011 年,研究人员发现大黄蜂携带一种微生物,这种微生物通过蜂巢进行社会传播,保护蜜蜂免受这种寄生虫的侵害,这是有益社会传播的明显案例。
在一些啮齿动物研究中,特定细菌的存在与否可以决定小鼠是否表现出社交缺陷,避免与同伴互动。肠道细菌在吸引果蝇为其配偶方面发挥了作用。微生物可以影响身体产生催产素等荷尔蒙,后者在亲密关系中起着强大的作用。
扩展阅读:微生物和你的爱情生活有什么关系?
对于人类,我们知道合住一所房子的人也有共同的微生物群。2014年,芝加哥大学和阿贡国家实验室的微生物学家 Jack Gilbert 和他的同事对七个家庭及其住宅进行了六周多的研究,他们发现每个家庭中的微生物群很容易相互区分,每个人都可以被家人识别。研究期间搬家的三个家庭都带着他们的微生物特征。
在婴儿出生后的第一年,肠道中一半的微生物菌株与母亲共享。母亲的影响力随着时间的推移而减弱—— 大约从 3 岁时的 27% 下滑到 30 岁时的 14%,但并没有消失。
其他家庭成员也是肠道微生物的重要来源。4 岁以后,孩子与父亲和母亲共享的微生物菌株数量相似。
分开生活的时间越长,彼此远离的双胞胎共享的肠道微生物就越少。一起长大的双胞胎有大约30%的共享菌株,在分开生活30年后下降到大约10%.
共享甚至发生在几个农村生活组的家庭之间:与来自不同村庄的人相比,来自同一个村庄不同家庭的人往往在肠道微生物方面有更多的重叠。
住在一起的人,往往有相同的口腔微生物菌株,而且住在一起的时间越长,他们共享的越多。

人类有体味,许多动物都有气味标记腺体。几十年前,研究人员提出,产生气味的不是动物本身,而是微生物,微生物产生挥发性有机化合物,这些化合物通过空气传播,从而产生真正的气味。如果我们用气味来相互交流,微生物很可能是产生交流的中间人。
牛津大学实验心理学系教授 Robin Dunbar 博士说:我们不仅在社会世界中进化,也在微生物世界中进化。
社会关系对人类健康和死亡率产生持续影响,高度社会融合和良好的人际关系的影响,远远超过戒烟或保持正常体重等个人行为对死亡率的保护作用。
研究表明,身心健康以及长寿的主要预测指标是我们亲密关系的质量和数量。事实上,饮食、体重、酒精摄入和环境污染物(包括空气质量差)等其他生活方式方面对健康和幸福的影响,要比良好的社交和心情影响要小。

对微生物群数据整合研究已有 60 年历史的威斯康星纵向研究中,发现与家人和朋友的社交与人类粪便微生物群的差异有关。
对配偶 (N = 94) 和兄弟姐妹对 (N = 83) 的分析进一步表明,与兄弟姐妹相比,配偶拥有更相似的微生物群和更多共同的细菌类群。与独居者相比,已婚者拥有更多样化和更丰富的微生物群落。这些结果表明,人与人之间的互动,尤其是持续的亲密婚姻关系,会影响肠道菌群。
研究人员还发现,拥有更强大社交网络的人通常具有更多样化的肠道微生物组,而更孤立、微生物多样性更少的人面临更大压力和焦虑的风险更高。健康的肠道微生物组及其多样性甚至与新生儿时期更高水平的社交能力相关。
而且这种现象不仅限于人类。灵长类动物研究实际上表明,黑猩猩社会中,通过社会互动共享微生物(水平传播)比母婴微生物共享(垂直传播)导致更高的肠道微生物组多样性。

来自佛罗里达州立大学和其他机构的研究人员比较了疫情前和疫情后的数据,发现人类社会中个体的四个特征有所下降:外向性,开放性,宜人性和尽责性。但是第五个特征,神经质,在人群中,尤其年轻人中增加了。最近对 COVID 流行期间近 5000 人进行的一项横断面研究发现,神经质与焦虑和抑郁呈正相关。
研究人员评估了 672 名成年人(23 ~ 69 岁),发现当按上面提到的五大人格特质进行分层时,微生物组的多样性和组成显示出显著差异。
LEfSe分析人格特征肠道微生物丰度的两组差异
doi.org/10.1016/j.bbi.2017.12.012
对于神经质特征得分较高的个体,研究人员发现γ-变形杆菌(Gammaproteobacteria )的水平升高,其中包括多种属,包括潜在的病原体,如肠杆菌(Enterobacter)、埃希氏菌(Escherichia)、嗜血杆菌、克雷伯氏菌(Haemophilus)、假单胞菌(Pseudomonas)、志贺氏菌(Shigella)和弧菌(Vibrio)。这些菌群水平升高也见于责任心得分较低的个体,这与较低的动机和自律水平有关。
作者还在这些高度神经质-责任心较低的个体中发现了 HPA 轴激活和炎症标志物升高。
此外,值得注意的是,肠道屏障通透性增加允许细菌移位到肠腔外,以及循环中革兰氏阴性细菌脂多糖 (LPS) 毒素的存在可能在神经质的生理学中发挥作用。
人类微生物组杂志的一项研究描述了肠道微生物通过神经、免疫、内分泌和神经递质途径对人格特征的影响。该研究包括 655 名平均年龄为 42 岁的成年人(83% 为北美人)。
作者发现,焦虑和压力的增加以及睡眠质量的下降与微生物组组成的改变和多样性的降低显着相关,特定的细菌属与特定的行为特征相关。例如,特定的拟杆菌属菌株与抑制性神经递质 (GABA) 的产生有关,这种神经递质对于抵御压力和抑郁症很重要。

扩展阅读:
近日,来自爱尔兰科克大学的研究人员发现,社交焦虑症中的肠道微生物群发生变化。对 49 个粪便样本(31 个病例和 18 个性别和年龄匹配的对照)分析,通过 β 多样性衡量的总体微生物群组成,发现社交焦虑症组和对照组之间存在差异,并且在属和种水平上发现了一些分类学差异。
在属水平上发现,Anaeromassillibacillus和Gordonibacter在社交焦虑症组中升高,而Parasuterella在健康对照中富集。
在物种水平上,发现Anaeromassilibacillus sp An250在社交焦虑症患者中更丰富,而Parasutterella excrementihominis在对照组中含量更高。肠道代谢模块“天冬氨酸降解I”在社交焦虑症患者中升高。

邓巴数字表明,人类无法在认知上管理大于 150 人的有意义的社会群体。尽管这个数字本身一直存在争议,但很明显,个人的社交需求因人格类型和内在特征而异:
注释:邓巴数字——人类社交网络的节点不会超过150个,即和你保持友好关系的人在150人以内。这个著名的论断是由英国牛津大学人类学家罗宾·邓巴在20世纪90年代提出。
对于我们所有人来说,无论性别、年龄或现有社交网络规模,如果长期处于相对隔离状态,有时甚至是绝对隔离,对我们的社交都显著影响。尽管网络是一种帮助我们保持高效和虚拟连接的工具,但它并没有显著减轻与封锁相关的孤独感和抑郁感,尤其是对于那些可能很少接触和/或不太习惯使用网络的老年人。
尽管很少有关于 COVID 感染对大脑本身影响的纵向研究,但英国的一项此类调查评估了近 400 名在感染 SARS-CoV-2 之前和康复后接受过脑部扫描的人。研究人员发现,与匹配的对照组相比,COVID 感染者的全脑体积和灰质总量减少,默认网络(default network)发生显着变化,这表明社交技能和网络规模的丧失。
注释:默认网络(Default Mode Network)的概念被提出,并引起了很多神经科学家的关注。默认网络是由在脑处于静息状态时相互联系、维持健康代谢活动的若干脑区组成的网络,在个体从事如监控外界环境、记忆提取和控制自身心理状态等多种事务中发挥着重要作用。
默认模式网络会随着年龄的增长而发生变化,反映的是大脑神经细胞自发活动的组织模式,可能会与大脑的学习、记忆及认知等功能相关。(参考自百度百科)。
在急性感染 SARS-CoV-2 病毒后,许多人的健康状况发生了长期变化。事实上,据估计,全世界有超过6500 万人患有所谓的“长新冠”(Long COVID)。与长新冠相关的问题包括大脑特定问题,如心理健康状况、疲劳、睡眠问题和认知障碍。
与长新冠相关的各种各样的大脑相关问题以及数百万人遭受这些问题的困扰,促使研究人员和普通公众研究新冠病毒感染如何以及为什么会影响大脑。
一些可能导致大脑损伤的因素包括:病毒传播到大脑、与感染相关的炎症、代谢问题、微生物组变化和血管损伤。然而,同样重要的是要强调,许多人在疫情期间可能经历过与感染无关的类似大脑问题,这反映了在大规模封锁措施的背景下发生的压力、不健康饮食和生活方式的改变。
扩展阅读:
阳康后是否会二次感染,长新冠与肠道菌群的关联,多种潜在的相关干预措施
在 2023 年 3 月发表在《大脑、行为和免疫》杂志上的一项研究中,研究人员在动物模型中研究了 SARS-CoV-2 病毒,发现该病毒激活了大脑中的免疫细胞,加剧了大脑炎症。重要的是,我们现在知道大脑中的免疫细胞(称为小胶质细胞)可能在情绪、认知等方面发挥作用,而肠道菌群积极参与了这一过程。
肠道微生物影响大脑的方式有很多。例如,有些可能会分泌通过血液传播到大脑的信使分子。其他细菌可能会刺激从大脑底部延伸到腹部器官的迷走神经。细菌分子可能通过最近发现的位于肠道内壁的“神经足”细胞将信号传递给迷走神经,这些细胞感知其生化环境,包括微生物化合物。每个细胞都有一个长长的“脚”,向外延伸,与附近的神经细胞(包括迷走神经细胞)形成突触状连接。
扩展阅读:
间接影响也包括像炎症和免疫。越来越多的研究人员将炎症视为抑郁症和自闭症等疾病的关键因素。肠道细菌是免疫系统正常发育和维持的关键,研究表明,微生物的失调会破坏该过程,并促进炎症。
肠道微生物产物可能会影响肠内分泌细胞,这些细胞位于肠道内壁并释放激素和其他肽。其中一些细胞有助于调节消化和控制胰岛素的产生,但它们也会释放神经递质血清素,它会从肠道中逸出并传播到全身。
神经精神疾病的药物开发已经滞后了几十年,而且许多现有药物并不对所有患者都有效,甚至会引起不必要的副作用。越来越多的研究人员在基于微生物的治疗或“精神益生菌”中看到了一种有前途的替代方法。
事实上,临床研究表明,补充精神益生菌,不仅可以改善情绪和行为,还可以将大脑活动朝积极的方向转变,功能性磁共振成像证明了这一点。常见的精神益生菌例如:
• 干酪乳杆菌 W56(Lactobacillus casei W56)
• 乳酸乳球菌 W19(Lactococcus lactis W19)
• 嗜酸乳杆菌 W22(Lactobacillus acidophilus W22)
• 乳双歧杆菌 W52(Bifidobacterium lactis W52)
• 副干酪乳杆菌 W20(Lactobacillus paracasei W20)
• 植物乳杆菌 W62(Lactobacillus plantarum W62)
• 乳双歧杆菌 W51( Bifidobacterium lactis W51)
• 双歧双歧杆菌 W23(Bifidobacterium bifidum W23)
• 唾液乳杆菌 W24 (Lactobacillus salivarius W24)
精神益生菌的潜在作用模式
Singh S,et al.Microorganisms.2022
详见:环境污染物通过肠脑轴影响心理健康,精神益生菌或将发挥重要作用
当我们都在适应新常态时,逆转隔离对身体的影响与重新参与社交一样重要。一个重要的步骤是积极地重新调整平衡肠道微生物群,并重振肠脑轴。
健康的微生物有助于预防疾病。所以,平衡我们身体内部微生物种群与身体外部的消毒防护之间的关系很重要。继续坚持洗手,但要多吃纤维、发酵食品和益生菌,减少糖、重加工和红肉的过量摄入。
同时,加强锻炼、减少压力,野外能多走走就多走走。环境中的微生物也可以被摄入,并成为我们肠道菌群的一部分,在肠道内一些微生物种群可以帮助促进健康的肠道细胞。
扩展阅读:
“玩泥巴”也有利于健康?接触环境微生物群可能调节肠道菌群和免疫系统
结语
总的来说,肠道菌群与社会关系之间存在复杂的相互关联。
社会关系对肠道菌群的组成和多样性有着直接的影响。一个人的家庭环境、工作环境、与其他人的接触等因素都可能对肠道菌群产生影响。
此外,肠道菌群也可以反过来影响社会行为和认知能力。肠道菌群可以通过与中枢神经系统交流,来影响人体的认知和行为。肠道中特定种类的菌群可以影响情绪,从而表现出焦虑、抑郁等症状。
共享微生物带来的好处,可能会促使我们人与人之间彼此互动,同时我们也可以积极地去寻找和维护社交和友谊,以保持身心健康和肠道微生物群的健康。
主要参考文献
Holt-Lunstad, J., Smith, T. B., Baker, M., Harris, T. & Stephenson, D. Loneliness and social isolation as risk factors for mortality: a meta-analytic review. Perspect. Psychol. Sci. 10, 227–237 (2015)
Yang, Y. C. et al. Social relationships and physiological determinants of longevity across the human life span. Proc. Natl Acad. Sci. USA 113, 578–583 (2016)
Johnson KV. Gut microbiome composition and diversity are related to human personality traits. Hum Microb J. 2020 Mar;15:None.
Christian L.M. Gut microbiome composition is associated with temperament during early childhood. Brain Behav Immun. 2015;45:118–127.
Moeller A. Social behavior shapes the chimpanzee pan-microbiome. Sci Adv. 2016;2
Kim HN, Yun Y, Ryu S, Chang Y, Kwon MJ, Cho J, Shin H, Kim HL. Correlation between gut microbiota and personality in adults: A cross-sectional study. Brain Behav Immun. 2018 Mar;69:374-385.
Johnson KV. Gut microbiome composition and diversity are related to human personality traits. Hum Microb J. 2020 Mar;15:
Valles-Colomer M, Blanco-Míguez A, et al., The person-to-person transmission landscape of the gut and oral microbiomes. Nature. 2023 Feb;614(7946):125-135.
Barrett E., Ross R.P., O’Toole P.W., Fitzgerald G.F., Stanton C. γ-Aminobutyric acid production by culturable bacteria from the human intestine. J Appl Microbiol. 2012;113:411–417
Bruce-Keller A., Salbaum J.M., Berthoud H.-R. Harnessing gut microbes for mental health: getting from here to there. Biol Psychiatry. 2018;83:214–223
Lindenfors P, Wartel A, Lind J. ‘Dunbar’s number’ deconstructed. Biol Lett. 2021 May;17(5):20210158.
Dunbar, R. I. M. The anatomy of friendship. Trends Cogn. Sci. 22, 32–51 (2018)
Butler MI, Bastiaanssen TFS, Long-Smith C, Morkl S, Berding K, Ritz NL, Strain C, Patangia D, Patel S, Stanton C, O’Mahony SM, Cryan JF, Clarke G, Dinan TG. The gut microbiome in social anxiety disorder: evidence of altered composition and function. Transl Psychiatry. 2023 Mar 20;13(1):95.
Pollet, T. V., Roberts, S. G. & Dunbar, R. I. Extraverts have larger social network layers. J. Individ. Differ. 32, 161–169 (2011)
Roberts, S. G., Dunbar, R. I., Pollet, T. V. & Kuppens, T. Exploring variation in active network size: constraints and ego characteristics. Soc. Netw. 31, 138–146 (2009)
Savignac H.M., Kiely B., Dinan T.G., Cryan J.F. Bifidobacteria exert strain-specific effects on stress-related behavior and physiology in BALB/c mice. Neurogastroenterol Motil. 2014;26:1615–1627
Robb, C. E. et al. Associations of social isolation with anxiety and depression during the early COVID-19 pandemic: a survey of older adults in London, UK. Front. Psychiatry 11, 591120 (2020).
Douaud, G. et al. SARS-CoV-2 is associated with changes in brain structure in UK Biobank. Nature 604, 697–707 (2022)
Bzdok, D., Dunbar, R.I.M. Social isolation and the brain in the pandemic era. Nat Hum Behav 6, 1333–1343 (2022).
Bagga D, Reichert JL, Koschutnig K, Aigner CS, Holzer P, Koskinen K, Moissl-Eichinger C, Schöpf V. Probiotics drive gut microbiome triggering emotional brain signatures. Gut Microbes. 2018 Nov 2;9(6):486-496.

谷禾健康

越来越多的证据表明,肠道菌群定植紊乱和微生物多样性减少与全球非传染性疾病 (NCD) 的增加有关。影响儿童和青少年的非传染性疾病包括肥胖及其相关合并症、自身免疫性疾病、过敏性疾病和哮喘。饮食变化也与非传染性疾病的发病机制有关,并且由于饮食是肠道微生物群组成和功能的主要驱动因素之一,因此人们开始关注通过饮食干预,来促进健康的肠道微生物群,最终促进健康。
一些生物活性营养素,如长链多不饱和脂肪酸 (LC-PUFA)、铁、维生素、蛋白质或碳水化合物,已被确定在婴儿出生后的前 1000 天对婴儿生长、神经发育发挥重要作用,以及肠道菌群的建立和成熟。LC-PUFA 是中枢神经系统 (CNS) 的结构成分,对视网膜发育或海马可塑性至关重要。最近,乳脂球膜 (MFG) 的成分被添加到婴儿配方奶粉中,因为它们在婴儿发育中起着关键作用。
大量摄入蛋白质会导致婴儿期体重增加更快,但这与后来的肥胖有关。可消化的碳水化合物提供葡萄糖,这对中枢神经系统的充分运作至关重要;不易消化的碳水化合物 [例如人乳低聚糖 (HMO)] 是肠道细菌的主要碳源。婴儿期缺铁性贫血与精神和精神运动发育的改变有关。与维生素 B6 和 B12 密切相关的叶酸代谢控制表观遗传变化。
从历史上看,重点一直放在早期营养对生长模式和儿童体脂成分的影响上。证据表明,生命早期摄入过多的能量和快速或缓慢的生长模式与不良的发育结果有关;事实上,婴儿期体重快速增加是晚年肥胖的重要预测指标。
肠道菌群与营养失调与多种儿科疾病有关,营养素的摄入和肠道微生物群的定植和成熟是相互关联的,因此通过饮食干预来促进健康的肠道微生物群是一种有前途的方法,可以改善儿童健康结果。

本文讨论和总结评估营养和肠道微生物群对儿童健康结果影响的临床研究的最新发现,并分享使用营养方法有利地改变肠道微生物群以改善儿童健康结果的研究成果。
脂肪酸是许多脂质的主要成分,必须通过婴儿饮食提供必需的脂肪酸,以实现健康成长、神经发育、免疫系统和胃肠功能。
婴儿的脂肪摄入量占比
在生命的头几个月,多不饱和脂肪酸 (PUFAs) 的需求增加,因为快速生长和神经发育。婴儿的脂肪摄入量在母乳喂养期间很高,从开始添加辅食后的第一年下半年逐渐减少。脂肪营养需求量占每日总能量摄入:
细分各类脂肪酸的摄入量
最近,不同的国家确定亚油酸的摄入量应占总能量的 4%,而 α 亚麻酸应占总能量的 0.5%。
长链多不饱和脂肪酸 (LC-PUFAs)、n-3 二十二碳六烯酸 (DHA, 22 : 6n-3) 和花生四烯酸 (ARA, 20 : 4n-6) 是中枢神经系统细胞膜的功能成分,在神经传递具有关键作用。
欧洲食品安全局 (EFSA) 委员会已确定:
0 ~ 24 个月的 DHA 摄入量为 100 毫克/天;
0 ~不到6个月的 ARA 摄入量为 140 毫克/天;
ARA 和 DHA 由母乳提供
婴儿的 DHA 状态是通过母乳提供的,它取决于母亲的 DHA 状态;尽管如此,母乳中的 ARA 浓度始终接近总脂肪酸的 0.5%,通常高于 DHA,与 DHA 相比更稳定。
与大脑发育相关的脂肪酸
大量的 n-3 和 n-6 LC-PUFA 在器官和组织的膜中迅速积累。在胎儿生命的最后三个月和生命的头两年,DHA 在脑组织中积累,特别是在与注意力、运动控制和感觉统合相关的灰质区域,而 ARA 负责海马可塑性。
已经表明,ARA 的延伸产物肾上腺酸 (ADA,22:4n-6) 是细胞膜中的重要成分。ADA构成了大脑中近一半的n-6 LC-PUFA,n-6 LC-PUFA的含量远远超过n-3 LC-PUFA。
均衡摄入DHA和ARA对大脑功能和发育至关重要
事实上,生命早期较高的 DHA/ARA 比率与更好的认知结果相关。已经表明,神经发育结果有利于 DHA 与 ARA 的比例为 1:1 或 1:2,而与1:1和1:2的比例相比,1.5∶1的比例会降低大脑发育过程中红细胞中ARA的浓度。
在脂肪酸摄入量和线性生长之间建立关系的研究得出了不同的结论。其中一些人认为必需脂肪酸对于婴儿期的最佳线性生长很重要,也有研究人员没有发现任何关联。
乳脂球膜蛋白的健康益处
另一方面,脂肪的研究工作表明,乳脂球膜 (MFGM) 蛋白代表母乳的生物活性部分,可提供一些健康益处。这种膜组分由不同的生物活性成分(磷脂酰胆碱、鞘磷脂、胆固醇和脑苷脂、神经节苷脂等)组成,它们对大脑发育和免疫功能有积极影响并保护新生儿胃肠道调节肠道菌群组成。
饮食中脂肪酸的分布与肠道菌群的关联
我们通常认为饮食中脂肪过多会造成肥胖,实际上,饮食中脂肪酸的分布也可能改变肠道微生物群的组成和肥胖状况。最近,表明人乳中的 sn-2 脂肪酸与婴儿肠道微生物群之间存在显着关联;ARA 和 DHA 与拟杆菌属(Bacteroides)、肠杆菌科(Enterobacteriaceae)、韦荣球菌属(Veillonella)、链球菌属(Streptococcus)和梭菌属(Clostridium)有关,参与短链脂肪酸(乙酸盐、丙酸盐和丁酸盐)生产的细菌,具有重要的免疫调节功能,在抵抗肠道病变的发展等方面发挥着关键作用,并且在母乳喂养后 13-15 天显着增加。
扩展阅读:脂肪毒性的新兴调节剂——肠道微生物组
蛋白质在生命的前 1000 天非常重要,因为它们在细胞结构中发挥着重要作用,并且是酶和神经递质的组成部分。
蛋白质推荐量
在出生后的头 6 个月内,每公斤体重/天的蛋白质推荐量为:
0 至 6 个月大时为 0.58 克;
6 至 36 个月大时为 0.66 克。
母乳中蛋白质种类多,有多种功能
母乳含有 400 多种蛋白质,多种功能如抗菌、免疫调节活性或刺激营养吸收等。蛋白质缺乏会导致生长发育不良以及运动和认知发育迟缓;然而,高蛋白质摄入会导致婴儿期体重增加更快,并与以后的肥胖相关。
使用婴儿配方奶粉喂养的婴儿在生命的前四个月内表现出正常的婴儿生长模式,婴儿的总蛋白质减少 1.0 g/dl(类似于母乳)。
辅食中蛋白质影响婴儿生长及肠道菌群组成
补充食品中的蛋白质来源和摄入量会显着影响婴儿生长并可能影响超重风险;以肉类和奶制品为基础的辅食会导致不同的生长模式,尤其是身高。
同时,补充喂养期间相关类型的富含蛋白质的食物,对配方奶喂养婴儿的肠道微生物组成和代谢物有影响;吃肉的儿童肠道群落富含厚壁菌门和粪杆菌属,同时变形杆菌门和双歧杆菌属减少。
扩展阅读:肠道菌群与蛋白质代谢
碳水化合物需求量
每日总能量摄入中的总碳水化合物需求量占比如下:
0 ~ 6 个月为 40-45%
6 至 12 个月以下为 45-55%
12 至 36 个月以下为 45-60%(接近成年人)
葡萄糖
葡萄糖对于中枢神经系统的充分运作起着关键作用,因为它是生长、神经冲动和突触的主要能量来源。葡萄糖由不同的碳水化合物提供给婴儿,例如乳糖,作为母乳中的主要糖分(范围为 6.7 至 7.8 g/dl),以及多种低聚糖,其含量约为 1 g/dl。
母乳低聚糖
母乳低聚糖 (HMO) 构成了婴儿无法消化的母乳碳水化合物的重要部分。母乳低聚糖具有益生元功能,可喂养胃肠道微生物群,并促进有益菌的生长;此外,它们还与多种生物学功能有关,例如对胃肠道发育和全身免疫的影响、双歧杆菌生成活性和抗感染、炎症调节、肠神经元激活和肠道运动,以及中枢神经系统功能的增强。
母乳低聚糖包括酸性低聚糖,主要是唾液酸化 [例如 6′-唾液酸乳糖 (6′-SL)、3′-唾液酸乳糖 (3′-SL)] 或中性低聚糖 [例如 2′-岩藻糖基乳糖 (2′-FL)]。
岩藻糖基聚糖是母乳中最丰富的母乳低聚糖形式 (80–90%) 。
聚糖
聚糖(glycans)是微生物的碳源,对宿主细胞和微生物之间的识别、信号传导和表观遗传调控至关重要,与广泛的免疫和代谢紊乱有关。双歧杆菌属和乳杆菌属与的生长之间存在显着相关性。在哺乳早期和晚期的婴儿肠道中。
几个临床前模型已经证明母乳低聚糖对认知功能的影响,但人类的临床数据尚未公布。
关于糖没有特定推荐量,2岁以下避免添加糖
关于糖,没有针对婴儿期糖的特定的每日参考摄入量。ESPGHAN 营养委员会建议,避免在 2 岁以下儿童的饮食中添加糖分。还建议避免饮用果汁或含糖饮料,因为过早摄入这些饮料会增加日后患 1 型糖尿病的风险。
为什么婴儿在 6 个月左右时需要添加辅食?与铁等营养素的需求有关
在婴儿出生前,胎儿会从母体中吸收铁元素,积累在肝脏中,以备出生后使用。然而,母乳中的铁含量相对较低,因此在婴儿 6 个月左右时,需要从饮食中摄取外源性铁以满足营养需求。
铁的需求量
0 ~ 6 个月为 0.3 毫克/天;
6 ~ 12 个月以下为 6-11 毫克/天;
12 ~ 36 个月以下的需求量为 3.9-9 毫克/天。
缺铁有哪些影响?
缺铁会影响大脑、神经和精神运动发育,因为铁是神经递质所需酶的组成部分。缺铁会导致携氧能力降低,从而导致生长发育所需的葡萄糖转化受限;这些限制可能导致生长迟缓、体重减轻和年龄增长,但与神经发育不同的是,它们可以通过补铁治疗来克服。
缺铁影响肠道菌群组成
母乳是短双歧杆菌的主要来源,它可以在二价金属通透酶和乳铁蛋白的帮助下获得管腔铁,促进这些有益细菌的生长,并从细菌病原体中隔离铁。缺铁导致肠道微生物群落失调,这反映在肠杆菌科(Enterobacteriaceae)和韦荣球菌科(Veillonellaceae)的相对丰度增加,以及与健康对照相比,红蝽菌科(Coriobacteriaceae)肠杆菌科和双歧杆菌科/肠杆菌科的丰度降低。
Coriobacteriaceae被确定为一个潜在的生物标志物,将运动与健康改善联系起来。
扩展阅读:人与菌对铁的竞争吸收 | 塑造并控制肠道潜在病原菌的生长
纯母乳喂养的婴儿摄入的维生素 D 低于最低推荐摄入量,远低于每日参考摄入量。
维生素D推荐摄入量
为避免因维生素D而可能出现的病症,例如骨矿化不足或软骨病,母亲每天补充 400 至 2000 IU 可以增加母乳中的维生素 D 水平;建议纯母乳喂养的婴儿接受阳光照射和补充维生素 D。
0 至 36 个月以下的婴儿维生素 D 营养需求为:
10 微克/天。
缺乏维生素D会引起什么?
维生素D诱导神经生长因子,促进神经突生长,抑制海马神经元凋亡。关键神经发育时期的缺陷会导致生命后期的行为、记忆和学习障碍。
低水平的维生素 D 会导致肠道通透性增加,产生慢性低度炎症状态。
维生素 D 与肠道菌群之间存在关联,在 3-6 个月大的不同种族婴儿的肠道微生物群组成中观察到一些差异,这些婴儿的母亲在怀孕期间补充了维生素 D 以预防其后代的哮喘和过敏症。
扩展阅读:维生素D与肠道菌群的互作
维生素 B12 的需求量
0 ~ 6 个月为 0.4 微克/天,
6 ~ 不到 12 个月为 0.5 至 0.8 微克/天,
12 ~ 36 个月以下为 0.6 至 1 微克/天。
叶酸的需求量
EFSA 推荐:
0 ~ 6 个月的叶酸营养需求为 65 微克/天,
6 ~ 12 个月婴儿的叶酸摄入量为 80 微克/天,
12 ~ 36 个月以下的需求量为100微克/天;
1-17 岁儿童的叶酸 (FA) 摄入量上限已确定为 200 – 800 微克/天。
叶酸和维生素B12的作用
叶酸和维生素 B12(钴胺素)作为参与广泛生物过程的辅助底物和辅助因子发挥着重要作用,例如核酸合成、糖酵解、糖异生和氨基酸代谢。
此外,叶酸和维生素 B12 以及单碳代谢循环所需的其他微量营养素辅助因子的状况可能会影响 DNA 甲基化,从而对健康产生长期影响。
叶酸——必须,但不要过量
众所周知,怀孕期间缺乏叶酸会导致后代出现神经管缺陷的风险更高。然而,高剂量的叶酸与更好的状态无关,与母亲或后代无关;事实上,怀孕期间摄入量高于 400 微克/天并没有明显的好处。母乳喂养期间补充叶酸可导致母乳总叶酸适度增加。
在儿童中,叶酸缺乏与认知发育受损以及腹泻和呼吸系统疾病增加相关;然而,补充叶酸对于减少这些病症并没有明显帮助。
过量摄入叶酸可能会产生潜在的不利影响,包括几种疾病(例如癌症、神经系统疾病、生长综合征、呼吸系统疾病和多发性硬化症)的发病率增加。
目前,由于食用补充剂或强化食品,很多欧洲儿童摄入大量叶酸;目前尚不清楚这些摄入量是否会造成伤害,尤其是在早期发育过程中,而许多组织中正在发生大量表观遗传变化。
缺乏维生素B12有什么影响?
当母亲的维生素B12状况不佳时,母乳中的含量会降低,会影响后代维生素B12的状态。维生素 B12 对中枢神经系统的代谢和维持至关重要,与叶酸一起在同型半胱氨酸代谢和髓磷脂的保护中起着关键作用。因此,维生素 B12 缺乏会导致覆盖颅神经、脊神经和周围神经的髓鞘受损,从而导致神经精神疾病的发展。
B族维生素缺乏影响肠道菌群
通过基因组重建和预测,针对几种B族维生素,预测整个微生物群落的代谢表型,发现微生物群落中有相当一部分是辅助营养物种(它们无法自己合成某些生命所需的化学物质,需要从外部环境中获取这些物质才能生存),它们的生存完全依赖于从饮食和/或原养型微生物中获取一种或多种B族维生素,通过特定的拯救途径(一种代谢途径,通过这种途径,微生物可以从外部环境或其他微生物的代谢产物中回收利用某些生命所需的化学物质,以满足自身生存所需)来实现。
膳食摄入影响:
母体甲基供体的摄入(胆碱、甜菜碱、叶酸、蛋氨酸)会改变其后代的DNA甲基化。观察到这种摄入量,特别是在围孕期,会影响婴儿口腔中与代谢、生长、食欲调节和维持 DNA 甲基化反应相关的基因的 DNA 甲基化。
细菌合成影响:
除了膳食摄入外,细菌叶酸生物合成也备受关注。细菌叶酸生物合成可以提供额外的叶酸来源,对健康结果和/或 DNA 甲基化具有重要意义。
在体外结肠模型中,研究发现补充甲钴胺和乳清可以提高厚壁菌门和拟杆菌属的比例,同时减少变形杆菌属的数量,其中包括一些病原体,如大肠杆菌(Escherichia)和志贺氏菌属(Shigella)等,以及假单胞菌属(Pseudomonas)。此外,研究还发现甲钴胺可以促进肠道细菌对脂质、萜类化合物和聚酮化合物的代谢,诱导外源性物质的降解,抑制转录因子和次级代谢产物(如维生素 B12)的合成。
扩展阅读:如何解读肠道菌群检测报告中的维生素指标?
新生儿肠道菌群的建立及发育
新生儿的肠道菌群既直接来自母亲,也来自分娩后的环境。微生物组在生命的头几个月经历动态演替和成熟,这一过程伴随着身体指标以及器官和神经认知发育的快速变化。
新研究结果强调母乳喂养和婴儿饮食会影响肠道微生物组成和功能。一项使用宏基因组鸟枪法测序的综合研究表明,停止母乳喂养(而不是引入固体食物),可以推动婴儿肠道微生物组的功能成熟,使其接近成人状态。
新生儿肠道菌群的影响因素
新生儿微生物组和免疫系统的不成熟似乎与肠道感染的易感性增加有关,特别是在 LMIC(中低收入国家) 环境中。虽然新生儿获得微生物群的时间各不相同,但多次接触,包括分娩方式、母婴饮食、药物、获得安全水和卫生设施以及多种宿主因素,是微生物群组成的主要决定因素。
母乳对婴儿的发育和成熟起着重要作用,微生物组在断奶时进入过渡阶段,此时微生物组会发生其他变化。
儿童营养不良和生长障碍是由膳食摄入不足和炎症之间复杂的相互作用驱动的,炎症通常是持续和/或反复感染和慢性疾病(包括镰状细胞病、艾滋病毒、先天性心脏病、心理障碍和内分泌或代谢疾病)的结果。
肠病是营养不良的一个重要驱动因素
肠病可能是肠病原体相关性腹泻病的结果,这在 LMIC 环境中的儿童中很常见,并且与死亡率、生长迟缓和认知发育不良的风险较高有关。
而这些环境中的许多儿童在存在或不存在已知肠病原体的情况下患有无症状肠病。这种肠病与非特异性持续粪口污染、反复肠道感染和小肠细菌过度生长 (SIBO) 有关。这种肠病与其他慢性肠道炎症有一些相似之处,包括克罗恩病和溃疡性结肠炎。它与发育迟缓密切相关,可能通过营养吸收不良和食欲抑制间接影响生长,并通过生长激素-胰岛素样生长因子 1 (IGF-1) 轴直接影响生长。
肠道通透性增加也会对发育产生负面影响
肠屏障功能障碍和肠道通透性增加可能导致微生物和/或微生物产物易位,从而激活先天免疫反应并促进全身炎症,从而对生长产生负面影响。
扩展阅读:什么是肠漏综合征,它如何影响健康?
肠道微生物群会影响多种宿主功能,包括代谢调节和信号传导,通过获取膳食营养素和微生物群衍生的代谢物、免疫耐受和对病原体的抵抗力、昼夜节律以及与儿童健康成长相关的其他途径。
肠道菌群失调可能影响儿童生长发育
由于疾病、环境或药物暴露或其他损害而破坏微生物组的正常多样性和组成,可能导致生态失调,这是一种以致病菌大量繁殖、共生体丧失和多样性丧失为特征的状态。在一些人群中,生态失调与肥胖、2 型糖尿病、肝脂肪变性和肠道疾病有关。在儿童和部分人群中,生态失调与生长和神经认知发育不良以及反复感染、免疫力改变和炎症增加有关。
与营养良好的儿童相比,营养不良的儿童拥有“不太成熟”的肠道菌群,其多样性较低。生态失调导致营养提取效率低下、吸收不良、易患肠杆菌科等侵袭性疾病和肠道炎症,从而影响生长。
肠道微生物群与发育迟缓之间存在密切关联,表明存在因果机制
谷禾健康与长沙妇幼儿童保健中心实验室合作发表的临床研究,揭示了肠道微生物群对患有严重急性营养不良 (SAM) 等严重儿科病理状况的儿童的重要性;临床诊断为生长发育迟缓 (FTT) 的受试者和正常生长正常的早产受试者 (NFTT-pre) 在不同年龄段表现出明显的肠道菌群发育轨迹中断,并且其α多样性的发展以及观察到的 OTU 和 Shannon 指数不足,尤其是在具有 FTT 的受试者中。
此外,与正常相比,FTT组中细菌如拟杆菌、双歧杆菌、链球菌和大多数年龄歧视性细菌分类群的顺序定殖和富集及其微生物功能紊乱。我们的研究结果表明,发育迟缓的婴儿肠道菌群发育不全,具有潜在的临床和实践意义。

肠道菌群失调还与共生微生物的易位和系统传播以及对病原体的易感性有关。此外,共生细菌抵抗肠道炎症的功能能力降低,如产生短链脂肪酸和色氨酸分解代谢配体(驱动芳烃受体激活),可导致肠道炎症。
恢复肠道菌群稳态,可促进儿童生长发育
共生菌还维持先天性淋巴样细胞,这是白细胞介素IL-22 的主要来源,IL-22 可刺激抗菌肽,帮助防止病原菌的微生物移位和入侵。恢复稳态微生物组和相关代谢物,有可能逆转与生态失调相关的表型,并促进儿童的生长发育。
确定肠道微生物群落结构和功能的变化(包括确定它们与疾病的因果关系)以制定有效的干预措施,对恢复肠道微生物群落结构并改善健康生长发育至关重要。
确定可以在怀孕、婴儿期和儿童期实施的干预措施,以预防或改善这些导致生长发育不良的驱动因素,对于改善短期和长期健康与发育至关重要。
扩展阅读:
怀孕期间母体肠道菌群的组成和功能似乎与出生结局密切相关,包括体重和胎龄。在健康的非妊娠成人中,肠道微生物群由相对稳定的种群组成,主要由拟杆菌门、厚壁菌门、放线菌门、变形菌门和疣微菌门组成。微生物组的组成和多样性在怀孕期间发生了变化。例如,在怀孕期间,肠道微生物群 α 多样性和产丁酸菌减少,而双歧杆菌、变形菌和产乳酸菌增加。
最近对来自刚果、印度、巴基斯坦和危地马拉的孕妇进行的一项纵向研究表明,怀孕期间肠道微生物群的个体属和 α 多样性(丰富度)有所减少。
妊娠期肠道菌群与新生儿生长关联
最近进行了一项研究,以了解津巴布韦农村地区妊娠期肠道微生物群分类群与代谢功能对胎龄、出生体重和新生儿生长的关联。
结果证明,抗性淀粉降解细菌,主要是瘤胃球菌科、毛螺菌科和真细菌科,是主要的肠道类群,并且是出生体重、新生儿生长和胎龄的重要预测因子。
此外,这项研究表明,与淀粉和能量代谢、信号和维生素 B 代谢相关的细菌功能,与出生体重增加有关。这些结果表明,非洲农村地区母亲食用富含淀粉的饮食的饮食模式,可能会推动选择影响婴儿健康和成长的物种。
扩展阅读:肠道核心菌属——毛螺菌属(Lachnospira)
肠道菌群变化分别与妊娠糖尿病和高脂血症有关
谷禾健康与江南大学食品科学与技术国家重点实验室合作的临床研究成果表明妊娠糖尿病 (GDM) 通常与高脂血症合并症有关。改变的人类肠道微生物群分别与妊娠糖尿病和高脂血症有关,但与合并症无关。发现链球菌(Streptococcus)、粪杆菌(Faecalibacterium)、韦荣球菌(Veillonella)、普雷沃氏菌(Prevotella)、嗜血杆菌(Haemophilus)和放线菌( Actinomyces )在糖尿病加高脂血症人群中显着更高。此外,几种细菌与患有妊娠糖尿病和高脂血症的参与者的空腹血糖和血脂水平相关。
扩展阅读:肠道重要基石菌属——普雷沃氏菌属 Prevotella
母体微生物群的干预:益生菌
针对母体微生物群的干预措施有可能显着影响婴儿健康,因为孕期生态失调和母体暴露会影响微生物群的建立、免疫发育和代谢健康。正在评估妊娠期膳食补充益生菌(对宿主健康有益的活微生物),以预防妊娠相关并发症和不良出生结果,包括早产和极低出生体重。
一些数据表明,益生菌对孕妇或哺乳期妇女在治疗妊娠糖尿病 (GDM)、B族链球菌定植和乳腺炎方面具有有益作用。
鉴于已知的安全性,益生菌作为妊娠干预措施特别有吸引力。然而,迄今为止的研究还没有定论。在新西兰、芬兰、丹麦、瑞典、澳大利亚、伊朗和我国的女性中,补充各种益生菌和混合物(包括乳酸杆菌、链球菌和双歧杆菌菌株)对出生人体测量没有影响。但有一些数据表明益生菌单独或联合使用可能与低收入国家早产儿死亡率、坏死性小肠结肠炎和/或新生儿败血症的降低有关。
新生儿和婴儿是考虑针对微生物组进行干预的关键人群,因为婴儿微生物组在出生后经历快速进化。此外,婴儿期是生长和神经认知发育的关键时期,也是发病率和死亡率最高的时期。
婴儿肠道菌群的定植
来自拟杆菌门和放线菌门的专性厌氧菌会迅速定植婴儿肠道,主要是双歧杆菌属、拟杆菌属和梭菌属,在生命的前 6 个月内,其特点是多样性低。
母体肠道微生物群似乎对婴儿肠道的定植有显着贡献,而阴道和皮肤来源的细菌似乎更短暂,并且不会在新生儿期后持续存在于婴儿肠道中。
婴儿肠道菌群->免疫系统->宿主
婴儿肠道微生物群为免疫系统的发育提供信息,而免疫系统又协调维持宿主-微生物共生的关键特征。因此,肠道微生物组成和代谢的异常可能会破坏正在发育的免疫系统。
母乳喂养->断奶,肠道菌群变化
婴儿期的母乳喂养还通过母乳中微生物种类的直接转移和其他主要成分的调节影响婴儿生长和塑造肠道微生物群,例如人乳低聚糖(HMO – 人类酶无法消化的复合糖),分泌IgA 和抗菌因子。
断奶,即逐渐将固体食物引入婴儿饮食,是婴儿发育的一个重要里程碑。断奶也是肠道菌群快速扩张的时期,包括双歧杆菌、乳杆菌、韦荣球菌(Veillonella)、柯林氏菌(Collinsella)、普雷沃氏菌、粪杆菌属和大肠杆菌属以及参与复杂多糖代谢的其他物种的多样化和扩张。
断奶期微生物群受干扰,可能导致肠道感染的易感性
断奶时微生物群的扩大还与强烈免疫反应的诱导有关,一种“断奶反应”,其特征是与生命后期的免疫成熟和耐受性相关的调节性 T 细胞的扩增。
在小鼠中,断奶期间限制微生物组的成熟会导致免疫发育受损并增加对肠道感染的易感性。此外,在母乳喂养率高且在长时间断奶期间也接受补充饮食的孟加拉国社区队列中,发现了一个独特的“过渡”长双歧杆菌进化枝,它携带利用母乳和食物底物的酶。这种过渡性长双歧杆菌在断奶期间引入固体食物后会扩大,并且在孟加拉国以外的婴儿队列中也得到证实,尽管患病率要低得多。 这些发现表明,底物和混合喂养的持续时间也会影响肠道微生物组的结构和功能。
断奶期过后,肠道菌群高度依赖于饮食习惯
农村地区的儿童表现出拟杆菌门的显着富集和厚壁菌门的枯竭,普雷沃氏菌属的细菌数量独特丰富,显示出利用富含多糖的营养素的能力。
然而,在工业化国家,这些普氏菌肠型不太常见,断奶后微生物组的特征是拟杆菌和瘤胃球菌肠型的存在。
在试图了解微生物群落是如何共同配置的,包括描述组成成员之间的相互作用以及这些群落随着年龄的增长而成熟时,需要较大的样本人群队列,这也是谷禾一直推进的事情。
微生物群是否有一个稳定的架构?
综合众多的研究结果确定了一个由几十个细菌分类群组成的核心“生态群”,这些分类群在孟加拉国、印度和秘鲁的出生队列的健康成员中,在 20 个月及以后表现出一致的协变。研究得出结论,生态群网络是微生物群组织的一个保守的一般特征,建议这样的生态群可以提供一个框架来描述营养不良儿童的生态失调。
我们建议这样的生态群可以用作定量指标,用于定义旨在重新配置肠道微生物群落的靶向干预措施的功效。
婴儿绞痛、反流和便秘常常引起父母的痛苦也是儿科就诊的主要原因。如前所述,母乳喂养婴儿的微生物群通常被认为富含双歧杆菌和乳杆菌等“有益”细菌,以及梭菌等产气细菌的生长减少。
益生菌:罗伊氏乳杆菌DSM 17938减少哭闹
在随机对照试验中,与安慰剂相比,使用罗伊氏乳杆菌(L. reuteri)DSM 17938 治疗绞痛婴儿可显着减少哭闹时间、反流和功能性便秘。
相比之下,报道了与L. reuteri DSM 17938 相比,安慰剂组在治疗 1 个月时的烦躁时间短暂减少和睡眠持续时间更长。这种烦躁增加仅发生在配方奶喂养的婴儿中,而不发生在母乳喂养的婴儿中。
另一项最近的研究评估了L. reuteri DSM 17938 在 1 个月和 3 个月大时通过显着减少哭闹时间来预防绞痛。一般来说,罗伊氏乳杆菌DSM 17938 似乎可以减少患有绞痛的母乳喂养婴儿的哭闹时间,但是,这种益生菌在绞痛配方奶喂养婴儿中的作用需要进一步研究。
同样对于绞痛的预防,使用L. reuteri DSM 17938 似乎是有效的,但这需要在其他研究环境中得到证实。
扩展阅读:认识罗伊氏乳杆菌(Lactobacillus reuteri)
为了促进“有益”细菌的生长,婴儿配方奶粉中添加了特定的益生元,并在临床试验中进行了评估。
益生元:低聚半乳糖降低绞痛和反流的风险
在最近的一项双盲随机对照试验中,摄入补充低聚半乳糖的配方奶显示出与母乳喂养参照组相似的双歧杆菌和乳杆菌发育趋势,并且与接受不含低聚半乳糖的配方奶粉的婴儿相比,降低了绞痛和反流的风险。
合生元:减少哭闹、减轻疾病发作
人们对合生元提供“有益”细菌及其底物的兴趣也越来越大。
与随机分配到安慰剂配方奶粉的对照组相比,给婴儿喂食七种益生菌菌株和低聚果糖的混合物后,婴儿在第 7 天和第 30 天的哭闹时间减少了 50% 以上。
在另一项前瞻性双盲随机对照试验中,评估了含有嗜热链球菌(Streptococcus thermophilus)、保加利亚乳杆菌(L. bulgaricus)和动物双歧杆菌( B. animalis ssp. lactis)的合生酸奶饮料的效果。
与安慰剂相比,乳糖和菊糖对疾病发作(腹泻、上呼吸道感染和发热性疾病)的影响减少了发烧天数。 干预组大便稀便的频率更高,需要照顾孩子的次数也更多,但差异无统计学意义。
注意:
作者强调益生菌的干预并不是适合所有有症状的婴儿,婴儿的肠道菌群变化较快,益生菌及其组合的干预需要充分评估肠道菌群及其功能,了解其肠道菌群网络结构下,选择对应症状的干预方式才能确保安全和发挥干预的效果。
总的来说,在得出任何确定的结论之前,需要更多的研究来评估益生元和合生元在这些在不同类型儿童及其整体肠道微生态条件下的作用。
扩展阅读:
肠道微生物群与健康:探究发酵食品、饮食方式、益生菌和后生元的影响
人们还关注肠道菌群失调在过敏表型发生发展中的作用。
肠道菌群 & 过敏性疾病
肠杆菌科/拟杆菌比率↑ — 食物致敏的风险↑
据报道,在基于人群的加拿大健康婴儿纵向发育 (CHILD) 出生队列研究中,婴儿粪便中低肠道微生物群丰富度和升高的肠杆菌科/拟杆菌比率与随后食物致敏的风险增加有关。
瘤胃球菌科↓
–食物敏感 –特应性湿疹 –炎症性先天免疫反应过度
他们还发现食物敏感的婴儿在 1 岁时瘤胃球菌科的丰度下降。这可能与过敏性疾病高风险婴儿的病例对照研究结果一致,发现瘤胃球菌科的相对丰度较低与未出现任何过敏表现的婴儿相比,随后出现特应性湿疹的婴儿的粪便样本中。
值得注意的是,瘤胃球菌属的相对丰度较低也与炎症性先天免疫反应过度有关。
总的来说,这些发现进一步支持了这样一种假设,即缺乏潜在的免疫调节细菌可能会增加发生过敏表现的风险。由于瘤胃球菌能够降解纤维,并且是成人“核心”微生物组的一部分,未来的研究应该检验其重要性。
益生菌 & 肠道菌群
鼠李糖乳杆菌GG — 产丁酸菌↑
在最近的一项研究中,研究了益生菌对牛奶过敏婴儿肠道微生物组的影响。报道称,添加了鼠李糖乳杆菌GG (LGG) 的深度水解酪蛋白 (EHCF) 配方导致了与丁酸盐生产相关的特定细菌的富集。
丁酸盐是一种已知的结肠细胞底物,与增强肠道完整性有关。与单独使用 EHCF 相比,接受 EHCF + LGG 治疗的婴儿在治疗 6 个月后的丁酸产量呈双峰分布。
已知的丁酸盐生产者,Faecalibacterium,Blautia,Ruminococcus,Roseburia在高丁酸盐样本中富集,而拟杆菌显着减少。与牛奶不耐受的孩子相比,牛奶耐受的孩子Blautia和Roseburia富集。正如作者推测的那样,这些物种可能导致丁酸盐产量增加和肠道完整性增加。
扩展阅读:
肠道核心菌属——经黏液真杆菌属(Blautia),炎症肥胖相关的潜力菌
肠道核心菌属——普拉梭菌(Faecalibacterium Prausnitzii),预防炎症的下一代益生菌
母亲摄入益生菌降低孩子发病率
在该团队随后的2份研究中,其中在一项随机对照试验中,与无菌安慰剂牛奶相比,孕妇在围产期摄入含益生菌的低脂发酵牛奶可降低其孩子 2 岁和 6 岁时的湿疹发病率。然而,临床益处似乎与 3 个月或 2 岁时对肠道微生物多样性的影响无关。
由于益生菌仅给予母亲,另一种解释可能是通过影响母乳成分。在婴儿期益生菌随机对照试验的另一项后续研究中,对长期肠道微生物群的建立没有影响, 这与之前的报道一致。
注意:
虽然说荟萃分析报告,怀孕期间、母乳喂养期间和/或给婴儿服用益生菌可降低婴儿湿疹的风险,但证据仍然薄弱。因此,专家机构未能推出具体的指导方针。然而,在考虑所有关键结果时,世界过敏组织现在建议使用益生菌预防有过敏孩子高风险的孕妇和哺乳期母亲以及有高风险患过敏性疾病的婴儿(基于家族史)。
在他们的指南中,他们强调该建议是有条件的并且基于低质量的证据,并不能给出关于最有效的菌株、剂量或治疗的开始和持续时间的具体指导。因此,仍然需要更具体的指南和研究基础。
肠易激综合症
在一项评估肠易激综合征儿童低发酵底物饮食的初步研究中,该饮食与腹痛频率和严重程度的降低显著相关。与无反应者相比,对治疗有反应的儿童在基线和干预期间似乎具有不同的粪便微生物组。
在一项更大的、双盲、随机、交叉研究中,同一组使用 16S 测序研究了低发酵低聚糖、二糖、单糖和多元醇 (FODMAP) 饮食对肠易激综合症儿童的临床结果和肠道微生物组成的影响。
低 FODMAP 饮食减少了腹痛,并且对饮食有反应的儿童的微生物群具有更强的糖分解能力。作者建议,鉴定具有更强糖分解能力的微生物群可能作为预测对低 FODMAP 饮食反应的生物标志物。
克罗恩病
肠道微生物群环境的变化被认为是克罗恩病患者纯肠内营养治疗特性的中介。令人惊讶的是,与没有炎症性肠病家族史的健康对照相比,克罗恩病患儿在纯肠内营养过程中肠道微生物多样性、普拉梭菌和丁酸盐浓度有所降低。
当参与者恢复正常饮食时,这后来又恢复到治疗前的水平。伴随着这种假定的“不健康”微生物群,矛盾的是临床结果得到改善,结肠炎症标志物减少。然而,这些发现的相关性需要进一步阐明。
乳糜泻
在乳糜泻中,坚持严格的无麸质饮食 (GFD) 有时很困难,患者可能仍会出现临床症状和营养缺乏,随后持续发炎和肠道菌群失调。
由于特定的益生菌已被证明可以减轻炎症,因此在一项双盲探索性试验中,新诊断出患有乳糜泻的儿童被随机分配到摄入长双歧杆菌CECT 7347 或安慰剂组 3 个月。无论治疗如何,对 GFD 的依从性与生长参数呈正相关,与安慰剂组相比,益生菌组的身高有所增加。此外,益生菌处理减少了脆弱拟杆菌的数量组和分泌型 IgA。
在另一项评估两种益生菌短双歧杆菌菌株对 GFD 患儿影响的随机对照试验中,与安慰剂相比,干预减少了炎性细胞因子 TNFα 的产生。
总的来说,这些研究表明益生菌对患有乳糜泻的儿童可能有益,但需要在更大规模的试验中验证。
扩展阅读:双歧杆菌:长双歧杆菌
青年糖尿病环境决定因素 (TEDDY) 研究最近的一份报告中,该研究包括芬兰、瑞典、德国和美国患 1 型糖尿病的高风险儿童,肠道的组成和多样性都存在很大差异。即使在这个具有同源人类白细胞抗原 (HLA) II 类基因型并因此具有相似遗传风险的人群中,根据地理区域也存在显着差异。
这些差异的根本原因尚不清楚,因为即使在对早年生活和饮食变量进行调整后,差异仍然存在。
在同一项前瞻性队列研究中,还检查了早期接触益生菌和膳食可溶性纤维(可能影响肠道微生物群组成和形成免疫反应)与胰岛自身免疫的关系。与后期补充或无益生菌相比,益生菌暴露(≤27 天)与胰岛自身免疫风险降低相关。 相反,儿童早期膳食可溶性纤维的摄入与胰岛自身免疫或 I 型糖尿病无关。未来的研究需要检验这些发现的重要性。
人们一直对肠道菌群失调在影响儿科人群的大量疾病中的作用感兴趣。
儿童生长迟缓和认知发育不良的驱动因素是多方面的,包括饮食摄入量和多样性不足、暴露于反复感染、慢性疾病和肠道病理学,包括肠病和 SIBO。最近的研究表明,肠道菌群失调与发育迟缓之间存在密切关联,表明存在潜在的因果关系。这些研究强调需要确定肠道微生物群落的结构和功能改变,并恢复微生物组稳态和相关代谢物以促进低收入环境或国家儿童的生长发育。
儿童时期的肠道微生物组成高度依赖于饮食习惯。在营养不良的儿童中,与标准营养干预措施(如 RUSF)相比,含有当地可用成分的低热量密度 MDCF 可改善微生物组的成熟度和生长。未来我们需要努力探究不同地理环境和不同饮食习惯下中婴儿期微生物群的多样性,更深入地了解它们与免疫发育和生长的联系。
鉴定具有更高定植效率和临床有效性的适合当地的菌株可能提供巨大的潜力来优化可在怀孕、婴儿期和儿童期实施的干预措施,这可能会导致针对肠道微生物群的治疗和预防策略得到改进,并且也可能成为安全和具体指南的基础。
主要参考文献:
Njunge JM, Walson JL. Microbiota and growth among infants and children in low-income and middle-income settings. Curr Opin Clin Nutr Metab Care. 2023 Mar 6.
Videhult FK, West CE. Nutrition, gut microbiota and child health outcomes. Curr Opin Clin Nutr Metab Care. 2016 May;19(3):208-13.
Cerdó T, Diéguez E, Campoy C. Infant growth, neurodevelopment and gut microbiota during infancy: which nutrients are crucial? Curr Opin Clin Nutr Metab Care. 2019 Nov;22(6):434-441.
WHO. Levels and trends in child malnutrition: key findings of the 2021 edition of the joint child malnutrition estimates. United Nations Children’s Fund (UNICEF), World Health Organization, International Bank for Reconstruction and Development/The World Bank. 2021.
Gizaw Z, Yalew AW, Bitew BD, et al. Stunting among children aged 24-59 months and associations with sanitation, enteric infections, and environmental enteric dysfunction in rural northwest Ethiopia. Sci Rep 2022; 12:19293.
West CE, Renz H, Jenmalm MC, et al. The gut microbiota and inflammatory noncommunicable diseases: associations and potentials for gut microbiota therapies. J Allergy Clin Immunol 2015; 135:3–13.
Troesch B, Biesalski HK, Bos R, et al. Increased intake of foods with high nutrient density can help to break the intergenerational cycle of malnutrition and obesity. Nutrients 2015; 7:6016–6037.
Hiltunen H, Löyttyniemi E, Isolauri E, Rautava S. Early nutrition and growth until the corrected age of 2 years in extremely preterm infants. Neonatology 2018; 113:100–107.
Zheng M, Lamb KE, Grimes C, et al. Rapid weight gain during infancy and subsequent adiposity: a systematic review and meta-analysis of evidence. Obes Rev 2018; 19:321–332.
谷禾健康
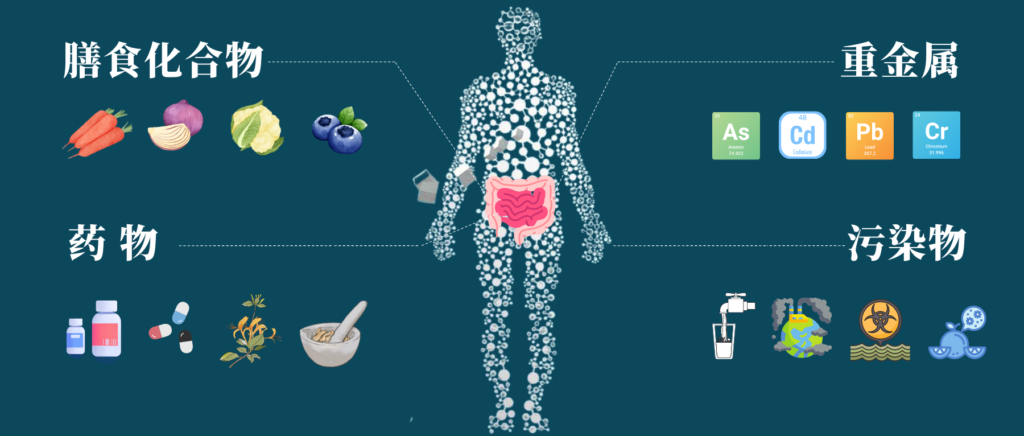
肠道菌群和人体健康息息相关,我们经常讲饮食、生活方式等都可以影响肠道菌群的组成,除了这些耳熟能详的因素之外,其他异源物如环境中的污染物,重金属,药物等都会影响肠道菌群,反过来,细菌也可以通过生物积累或化学修饰影响这些化合物。
通过皮肤接触、吸入或摄入等方式,我们每天都在和异生素接触,可能通过补充剂或药物自愿摄入异源物,或者通过受污染的食物和水被动摄入异源物。
肠道微生物群可以在吸收之前或之后与摄入的化合物相互作用。
了解肠道微生物群如何处理膳食成分的分子机制,是用“功能性食品”、益生菌和益生元治疗不同疾病的理论基础,有利于根据患者的代谢状况和肠道微生物群进行个性化营养定制。
与饮食类似,对肠道微生物群改变重金属、污染物、药物毒性进行研究,可以从肠道菌群代谢角度打开视野,进一步探讨从体内去除有害化合物并预防疾病的方法。
本文主要介绍了肠道微生物群对基本营养物质、重金属、污染物、药物、中草药等产生的代谢作用,并提供了基于肠道菌群的改善方式,以减轻这些有毒元素造成的损害。
什么是异源物?
异源物(xenobiotics),也就是说外源物质,来自希腊语,是指使人体从外界(包括肠道中的细菌作用产物)摄入体内的化学物质,例如药物、毒物、食品添加剂、环境污染物等。
人类肠道微生物群是一个多样而复杂的微生物群落,与宿主一起进化。肠道微生物群是宿主代谢过程的组成部分。
有时候会把肠道微生物群比作“器官”,可以干扰调节代谢的宿主基因。
人体肠道微生物群和异源物之间的大多数相互作用发生在胃肠道粘膜上。由于消化道的范围不同,不同的区域为微生物和代谢过程提供了不同的栖息地。
当我们说异源物的微生物代谢时,应该在宿主代谢过程的背景下进行。后者往往是同时发生的和相互竞争的。
▼ 异源物是怎么进入人体代谢的?
异源物经口服、吸收后在肠上皮细胞之间或通过肠上皮细胞传递,可被宿主酶处理或不被宿主酶处理。之后,它们可以通过门静脉输送到肝脏。
在肝脏中,异源物受到许多代谢酶的影响。因此,外源性代谢产物进入全身循环,从而使其分布到组织中并影响其他器官。
代谢产物可以与消化道的上皮细胞局部相互作用。当外源性药物通过静脉注射时,它们跳过了“第一道”代谢,立即进入全身循环。
而循环的外源性代谢产物进一步代谢并通过胆汁排泄排出,然后通过肠肝循环在小肠中重新吸收,或通过肾脏排出到尿液中。
▼ 异源物怎么和肠道菌群相遇?
可以通过多种途径与肠道微生物相遇。
小肠外源物中未被吸收或吸收不良的物质,继续进入大肠,被肠道微生物转化。
容易吸收或静脉注射的外源性物质最终可以通过胆汁排泄,到达肠道细菌。
▼ 肠道菌群影响异源物的生物转化
除了上面说的代谢过程外,肠道微生物群还可以影响异源物的生物转化,从而影响其功能和毒性。
同样,异源物可以诱导基因表达在人类肠道微生物群的直接参与异源物的代谢。结果表明,即使在短期暴露于异源物期间,这种表达也会上调。
异源物和微生物群之间的复杂相互作用
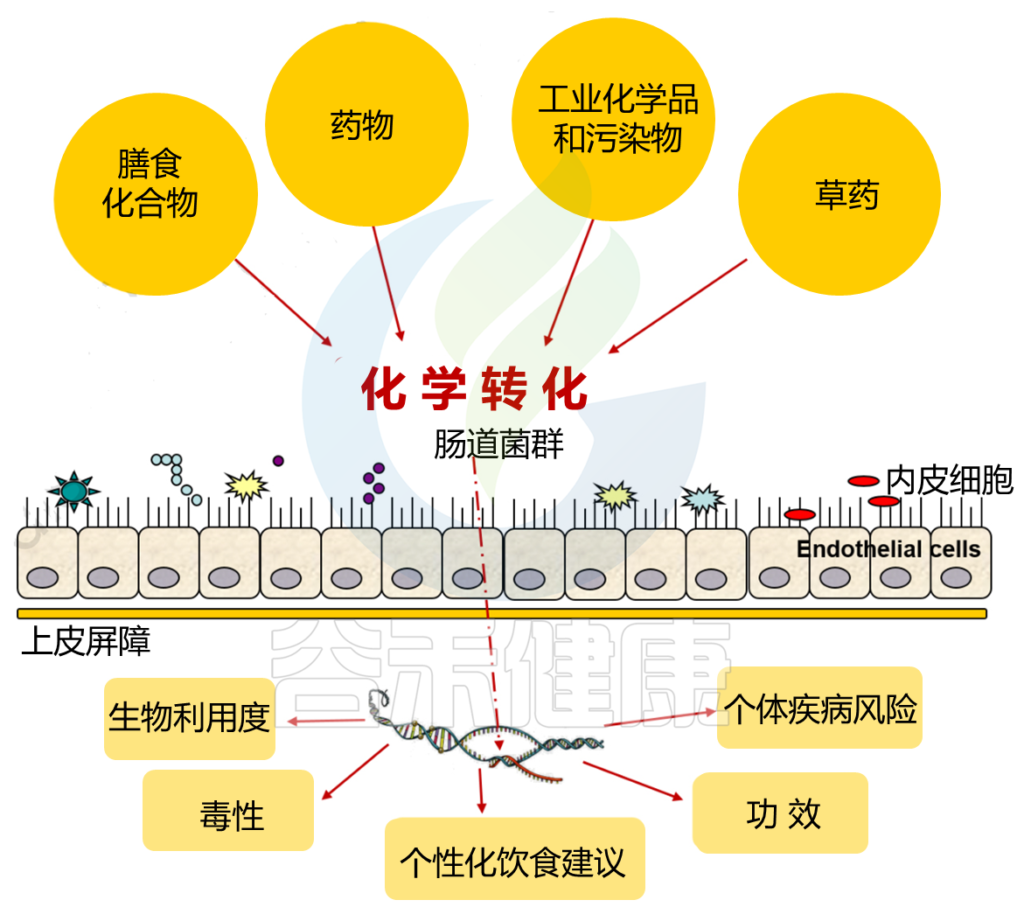
DOI: 10.2174/1389200221666200303113830
肠道微生物群对几种外源性物质的生物转化已通过体外和体内模型进行了广泛研究。然而,由于许多其他因素,例如在到达肠道细菌更普遍的肠道下部之前的有效吸收,或肝脏中的异源代谢,防止肠道微生物群遇到特定代谢产物,人类的异源物生物转化可能会发生很大变化。
同样,暴露于异源物质时,肠道菌群的结构和功能会发生特定变化。
摄入的外源物质的这些直接代谢导致了几种情况:
人类微生物群和异源物质相互作用是一个复杂的代谢网络,影响着双方。
饮食是人类健康的基石,目前已有许多研究将饮食模式与健康结果联系起来,这其中离不开肠道菌群的深度参与。在我们之前的多篇文章中也有相关阐述,这里我们简要回顾一下膳食化合物的代谢,重点关注重金属,污染物,药物代谢等异源物,逐一了解肠道菌群对异源物代谢过程,及其与健康之间的关联。
肠道菌群可以从多种膳食化合物中提取营养和能量,该过程的程度和类型在个体之间差异很大,主要是因为肠道微生物酶的存在和丰度不同,这里主要关注膳食蛋白质、脂类和多酚的转化。
膳食蛋白质可以用来做什么?
它可以体液平衡、细胞修复、血液凝固、激素、产生酶等。它也是肠道菌群的食物,主要作为氨基酸来源,可用于蛋白质合成和代谢能量的产生。
而肠道微生物群参与蛋白质代谢,它们在营养利用和宿主反应之间的相互作用中起着关键作用。
关于肠道微生物群参与蛋白质代谢详见:
小肠中的蛋白酶和许多肽酶对于将膳食蛋白质消化为氨基酸和寡肽是重要的。
肠道微生物群具有利用氨基酸的专门酶。
肠上皮细胞参与氨基酸分解代谢,并通过调节肠道屏障功能在先天免疫和适应性免疫中发挥重要作用。
肠道微生物生活在肠腔中,与粘蛋白密切相互作用,其任何变化都可能改变粘蛋白分泌。据推测,肠道微生物群利用必需和非必需的管腔氨基酸。
➤ 肠道菌群的组成及位置影响氨基酸代谢
肠道菌群的组成及其在肠道中的位置,对于确定膳食蛋白质的生产速率和氨基酸的代谢命运至关重要。
约氏乳杆菌(Lactobacillus johnsonii)
约氏乳杆菌是小肠中的常见居民,这种菌缺乏编码参与氨基酸生产的生物合成途径的基因。
约氏乳杆菌不利用氨,也不参与硫同化的代谢途径。但约氏乳杆菌产生一种胞外蛋白酶、3种寡肽转运蛋白、超过25种胞浆肽酶和20种氨基酸渗透酶型转运蛋白。这些有助于约氏乳杆菌吸收外源氨基酸或肽用于蛋白质合成。
➤不同菌群蛋白水解活性可能直接导致人类疾病
这里拿乳糜泻来举例,肠道微生物群与乳糜泻有关。乳糜泻是一种常见的自身免疫性疾病,其特征是对小麦基食物中的膳食面筋产生炎症反应。
这种富含脯氨酸的蛋白质避免了宿主蛋白酶的完全消化,从而产生了高分子量的免疫原性肽。肠道微生物群可能通过改变面筋蛋白水解来影响乳糜泻。
乳糜泻患者的肠道菌群是如何引发疾病的?
健康人和乳糜泻患者的粪便悬浮液,对面筋蛋白和免疫原性肽的处理方式不同。
一般健康人通过乳酸杆菌产生肽,而乳糜泻患者不一样,乳糜泻患者会多一些铜绿假单胞菌,这种铜绿假单胞菌产生的面筋衍生肽,更加易于穿过小鼠肠道易位,并引发增强的面筋特异性免疫应答。
所以说,鉴定负责肠道微生物面筋加工的特定蛋白酶,不仅可以更好地理解乳糜泻,还可以为该疾病的治疗干预提供信息,包括酶或益生菌治疗。
肠道微生物还可以将从膳食蛋白质中获得的氨基酸(包括l-苯丙氨酸、l-酪氨酸和l-色氨酸)代谢为一系列生物活性产物。例如,肠道细菌可以将l-色氨酸代谢为多种产物,包括抗氧化剂吲哚-3-丙酸、神经递质色胺和吲哚,后者可以通过肝脏酶进行羟基化和硫酸化,生成尿毒症毒素硫酸吲哚。
关于色氨酸代谢详见:
肠道微生物对脂质吸收很重要。
➤ 胆固醇的吸收
饮食胆固醇是西方饮食中与心血管疾病风险增加相关的主要成分。吃进去的胆固醇,被小肠吸收,随后经历胆汁排泄和肠肝循环。
➤ 肠道微生物群对胆固醇的影响
粪甾醇不能被重新吸收并被排出体外。通过肠道微生物的减少,胆固醇产生了不可吸收和排泄的粪甾醇。因此,这种转变有效地去除了循环中的胆固醇。
人类粪便中高达50%的类固醇是粪甾醇。高胆固醇降低患者和低胆固醇降低患者体内微生物群,分别定植的无菌小鼠,产生不同数量的粪甾醇。
还有研究表明,降低胆固醇的细菌可能会降低血清胆固醇。
➤ 降低胆固醇的细菌
对降低胆固醇的肠道细菌——产粪甾醇真细菌的研究表明,粪甾醇合成可能涉及氧化为5-胆甾-3-酮,然后烯烃异构化为4-胆甾-3-烯酮,共轭还原和酮还原。
确定负责这些修饰的酶并确定其在患者体内的丰度可能是关键,因为抑制胆固醇重吸收是降低胆固醇水平的有效方法。
关于脂质代谢,详见:
多酚是天然植物化合物,是人类饮食中最丰富的抗氧化剂,可以预防慢性退行性疾病。
多酚包括植物源性食物中存在的一类次级代谢产物,当它们作为纯化合物或富含多酚的提取物被纳入饮食中时,可以通过调节肠道微生物群的组成产生健康影响。当每天摄入高达2g时,多酚是重要的膳食生物活性成分。
它们的微生物调节是预期的,它们的化学结构,包括许多酚基团,表明了潜在的抗菌作用。
即使是不可吸收的聚合物化合物也会被肠道微生物群转化为可吸收的生物活性代谢物。
膳食多酚的生物利用度主要取决于肠道微生物群的组成:
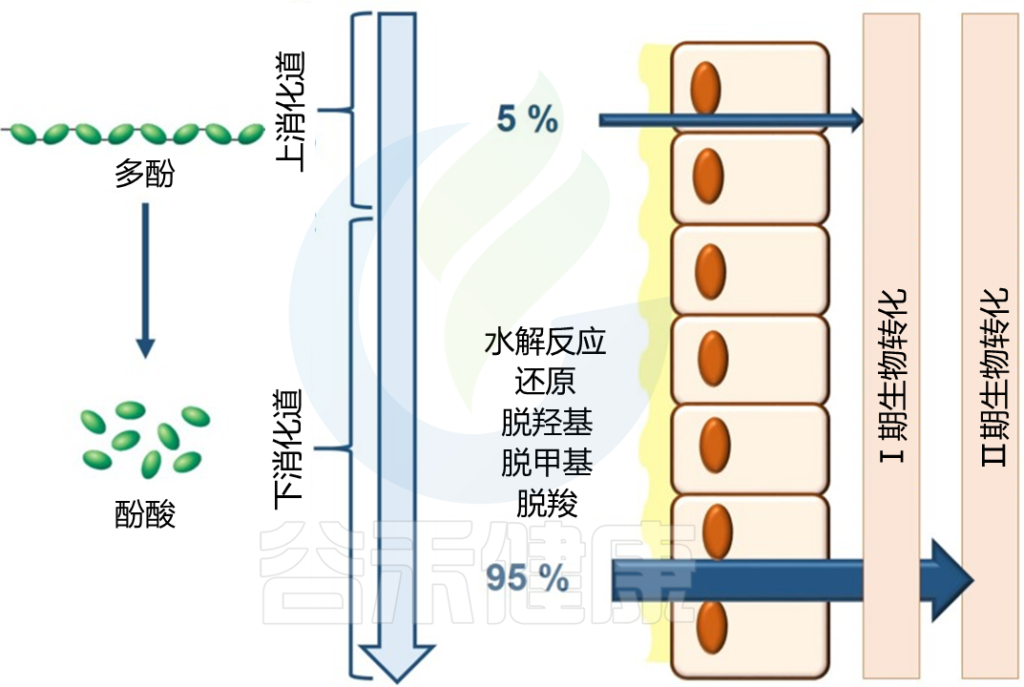
Westfall S, et al., Front Neurosci. 2019
肠道微生物群进行的三个主要分解代谢过程是水解、裂解和还原反应。在这些分解代谢反应之后,释放的苷元可能进行 II 期代谢并被肠道微生物群转化为简单的酚类衍生物,从而促进身体吸收。
关于肠道菌群代谢多酚详见:
近年来,人们越来越关注环境污染物对健康带来的影响。虽然说大多数污染物不直接针对肠道微生物群,但其中一些污染物可以通过不同途径进入人体,并与肠道微生物相互作用。
肠道菌群 ⇔ 污染物
肠道菌群活动可以改变这些化学品的毒性和生物利用度,并延长宿主接触有害化合物的时间。
暴露于环境污染物会改变肠道菌群的组成,导致能量代谢、免疫系统功能、营养吸收和/或产生其他毒性症状。
在评估这些物质的安全性时,必须考虑肠道微生物代谢的后果。这里讨论了几种与人类疾病风险有关的化学物质——重金属、持久性有机污染物和杀虫剂,有证据表明微生物代谢会影响其毒性。
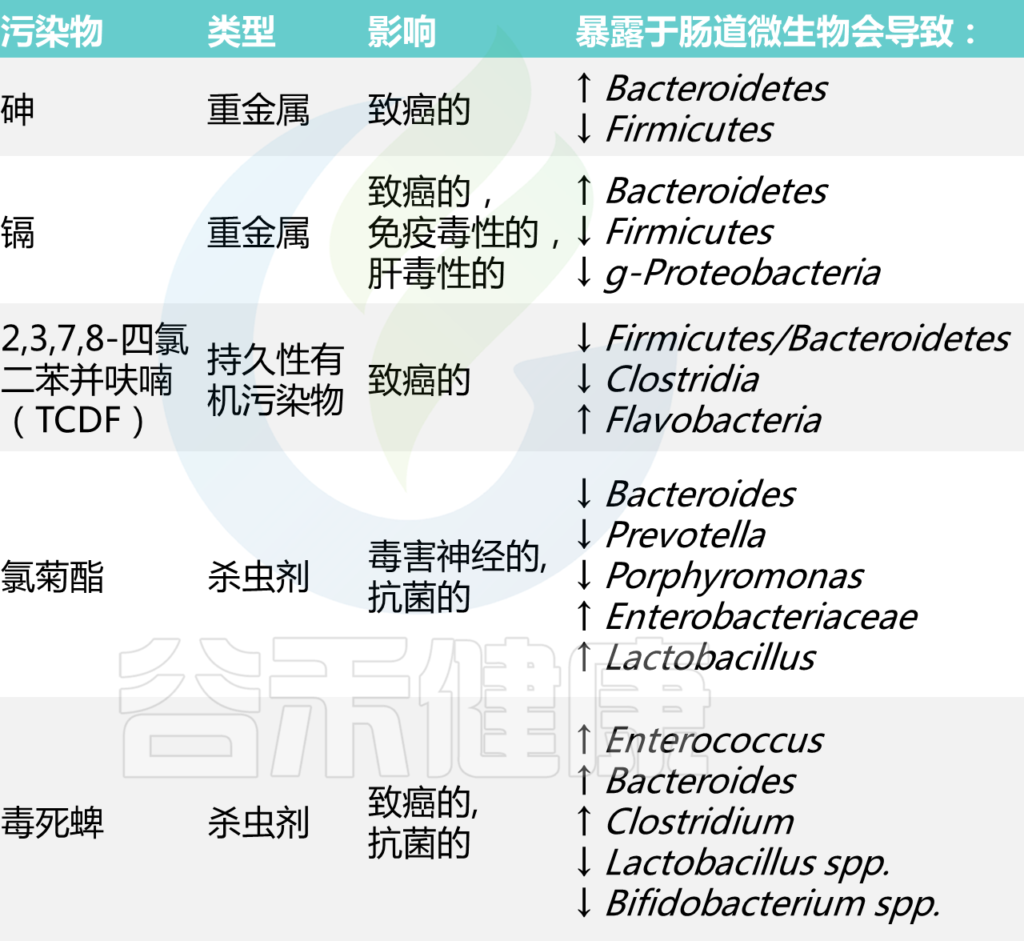
DOI: 10.2174/1389200221666200303113830
环境中的重金属与许多有害影响有关,包括致癌、氧化应激和对免疫系统的影响。几项研究表明,接触重金属也可能导致肠道微生物群失调。我们将以砷、镉、铅为例,展示重金属如何与肠道微生物相互作用,又会带来哪些健康后果。
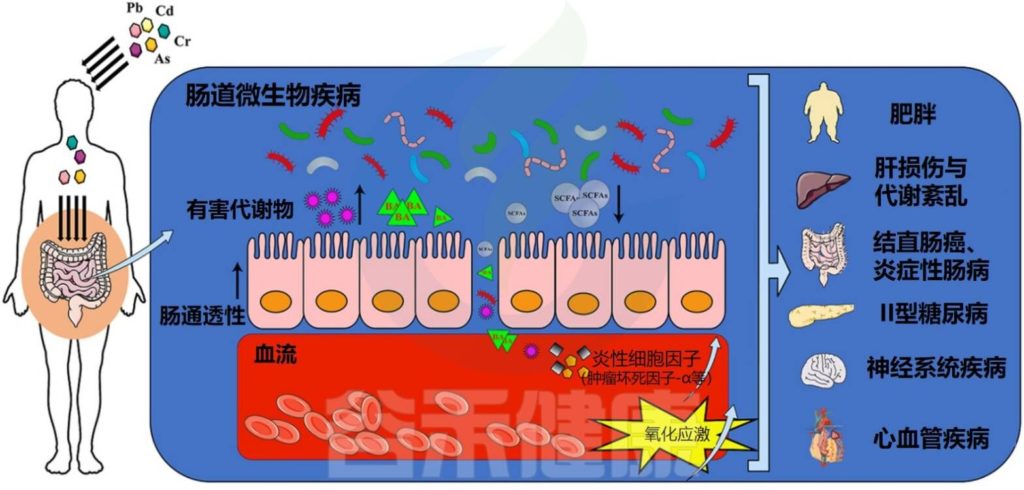
Liu X, et al.,Environ Pollut. 2023
砷(As)
砷(As)是最常见的有毒环境化合物之一。它具有已知的致癌作用。砷能够以砷酸盐[As(V)]的形式附着于固体表面。As(V)摄入并暴露于肠道微生物群后,会诱导硫化和甲基化。
研究发现,小鼠仅4周暴露于10mg/L的砷,就会显著干扰肠道微生物群的组成和代谢组学特征,拟杆菌门的丰度显著增加,而厚壁菌门的丰度则显著减少。这些变化与含吲哚的代谢产物、脂质代谢产物、异黄酮代谢产物和胆汁酸代谢产物有关。
➤砷暴露通过影响菌群及其代谢产物,损伤肝脏,从而影响健康
砷暴露会导致肠道微生物死亡,从而进一步损害肠道中的砷代谢。一旦砷不能代谢出体外,宿主的健康就会受到影响。砷暴露导致的肝脏损伤可能是由于肠道微生物代谢产物(如LPS)的变化所致。这种代谢产物是肝脏损伤的重要指标,并间接激活肠-肝轴。这表明肝脏是砷毒性的主要靶器官之一。
➤砷暴露影响脂质代谢,碳水化合物代谢
砷暴露极大地影响血清和肝脏的脂质稳态,主要影响脂质代谢,脂质代谢失衡与神经系统疾病有关,例如帕金森氏症、阿尔茨海默氏症和肌萎缩侧索硬化症;而肠道菌群在宿主脂质代谢中起着极其重要的作用,因为细菌产生的短链脂肪酸是宿主脂质合成、脂肪酸氧化和脂肪分解稳态的主要调节剂。
扩展阅读:
砷暴露还显着影响碳水化合物代谢途径。在砷暴露小鼠的肠道基因组中,参与淀粉利用系统的基因丰度显著增加。
➤食源性砷暴露不同于水源性砷暴露,会延长砷在胃肠道中的滞留时间
研究发现,小鼠粪便样本中接触30天砷,厚壁菌门和拟杆菌门的比例没有变化,该比率在暴露后 60 天显着降低,并且糖酵解、糖异生和肌醇磷酸代谢等碳水化合物代谢相关基因显著下调。
食源性砷暴露的影响是持久的。砷会导致参与丙酮酸代谢的几种酶水平下降,乙酸激酶和3-羟基丁基辅酶a脱氢酶水平下降,从而导致能量代谢异常。淀粉利用系统中susB、susC、susD和susR基因的增加,虽然缓解了这一情况,但碳水化合物代谢最终受到影响。
镉(Cd)
镉(Cd)用于电池、塑料、金属镀层和颜料的生产。镉以其致癌性、免疫毒性和肝毒性而闻名,它还诱导氧化应激。
研究人员发现雄性小鼠暴露于低剂量的镉(饮用水中为10mg/L)10周后,厚壁菌门和g-变形菌门的丰度降低,盲肠和粪便中拟杆菌门的丰度增加。肠道微生物组组成的这些变化与雄性小鼠血清中脂多糖(LPS)水平升高、肝脏炎症和能量代谢失调有关。
➤镉暴露影响脂肪代谢,引起“肠漏”,减少短链脂肪酸,引发炎症
镉暴露会引起代谢功能的显着变化,影响脂肪代谢并最终导致脂肪堆积。
镉暴露引起的肠道菌群失调会增加 FITC-葡聚糖,FITC-葡聚糖是肠道通透性的重要生物标志物,肠道通透性的增加会改变炎症和新陈代谢。
镉暴露会导致产短链脂肪酸菌数量急剧减少,并降低丁酸和丙酸的含量,短链脂肪酸减少会导致能量代谢受损,也会诱发炎症反应。
铅(Pb)
铅暴露没有明确的阈值。研究发现,血液中任何浓度的铅都会对成人和儿童产生毒性作用。铅中毒可直接影响中枢神经系统、肾脏和血压。
铅通过消化系统在体内蓄积,并对肠道微生物的组成产生显着影响,除了改变肠道多样性、菌群的组成之外,在一定程度上也改变代谢功能。铅对菌群影响主要是,厚壁菌门丰度减少,拟杆菌门丰度增加:
扩展阅读:
铅暴露还会导致某些代谢物和代谢途径发生变化。主要涉及胆汁酸、维生素、氮代谢、氧化应激、防御机制和能量代谢。而且,宿主释放有害代谢产物等变化可能会加剧铅造成的损害。
氮代谢的主要变化是UreE的激活;亚硝酸盐加速向一氧化氮生成,一氧化氮过量可能导致氧化应激、神经毒性、免疫抑制等问题。
注:UreE是一种激活尿素酶的共蛋白,导致尿素酶升高,最终导致肠道细菌中尿素的减少。
重金属暴露后肠道菌群紊乱引起的主要代谢变化
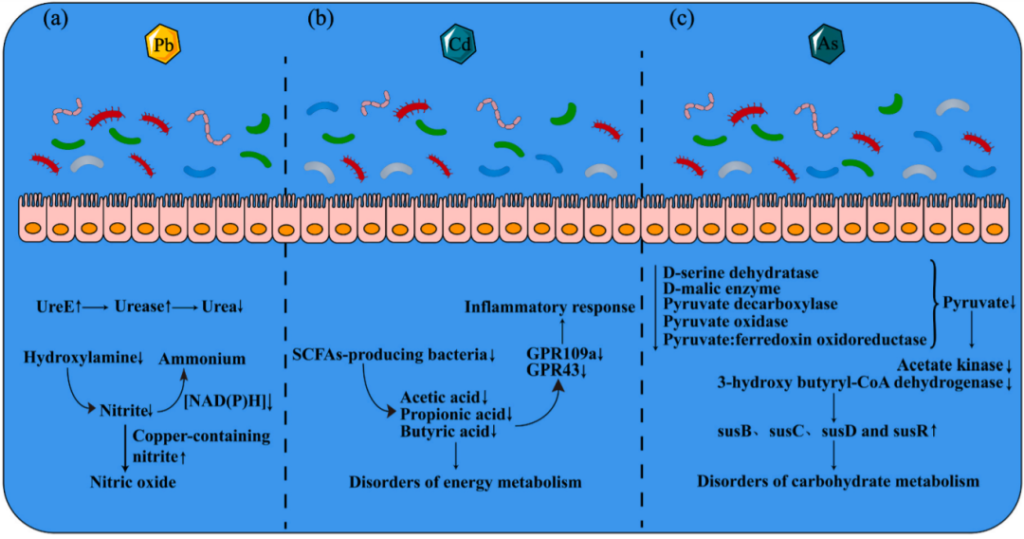
Liu X, et al.,Environ Pollut. 2023
肠道微生物群的研究为评估重金属毒性机制提供了一种新的途径。
持久性有机污染物(POP),包括有机氯农药、多氯联苯(PCB)、多溴二苯醚和多环芳烃(PAHs),是一种持久性和高毒性的合成化合物,可在生物组织中积累,可能与肥胖、糖尿病、自身免疫性疾病和某些发育障碍发病率上升相关。
扩展阅读:
通常,持久性有机污染物在摄入食物或水后会暴露于肠道微生物群,因此需要了解它们对肠道微生物的影响。
多氯联苯(PCB)
多氯联苯具有优异的介电财产和化学稳定性,因此用于制造冷却液、变压器、液压油和润滑剂。多氯联苯被认为是致癌物,可在小鼠暴露2天内显著改变肠道微生物群组成。肠中的产气荚膜梭菌Clostridium perfringens 和贝氏梭菌Clostridium beijerinckii可以降解多氯联苯,通过去除氯原子并打开苯环。
暴露于2,3,7,8-四氯二苯并呋喃(TCDF)(一种PCB)至少5天,可降低厚壁菌门/拟杆菌门的比例,降低梭状芽胞杆菌的水平,并增加盲肠内容物中的黄杆菌Flavobacteria水平。上述肠道微生物群的改变与胆汁酸代谢的变化有关。此外,TCDF还可以抑制法尼素X受体(FXR)信号通路,从而引发细菌发酵导致的宿主的严重炎症和代谢紊乱。
扩展阅读:环境污染物通过肠脑轴影响心理健康,精神益生菌或将发挥重要作用
杀虫剂
在食品材料、水和土壤中检测到多种农药残留,这使得农药成为一个严重的环境问题。一些杀虫剂具有抗菌活性,因此能够改变肠道微生物组的组成。
氯氰菊酯(PEM)
氯氰菊酯(PEM)低剂量用于大鼠,结果降低了拟杆菌、普雷沃氏菌和卟啉单胞菌的丰度,增加了肠杆菌科和乳杆菌的丰度。这些菌群变化可能促进氯氰菊酯的神经毒性。
毒死蜱(CPF)
毒死蜱(CPF)是一种有机磷杀虫剂,通常用于水果、蔬菜和葡萄园。毒死蜱可以通过肝脏或肠道中的细胞色素P450酶代谢。毒死蜱增加了肠球菌、拟杆菌和梭菌的水平,但降低了大鼠肠道中乳酸杆菌和双歧杆菌的水平。
毒死蜱诱导的肠道失调导致粘膜屏障受损,细菌易位增加,并激活先天免疫系统。
以上是重金属等异源物对肠道菌群带来的影响,肠道菌群的变化也会给宿主健康造成影响。基于肠道菌群的层面进行干预,在一定程度上,可以帮助人体免受有害异源物积累带来的健康困扰,这些干预措施包括益生菌,饮食,运动等。
益生菌对重金属的作用
两种重金属解毒策略如下:
✔ 益生菌对重金属诱导的肠道细菌失调的保护
➦ 传统益生菌:
研究表明,鼠李糖乳杆菌(Lactobacillus rhamnosus sp.shermanii JS)和丙酸杆菌(Propionibacterium freudenreichii)能够在体内和体外成功解毒重金属。
有研究发现,含有鼠李糖乳杆菌GR-1 的益生菌降低了孕妇血液中砷和汞的含量。
以下菌株通过肠道重金属隔离和刺激肠道蠕动促进重金属的粪便排泄,从而减少重金属在肠道中的吸收,并逆转重金属引起的肠道菌群变化:
植物乳杆菌TW1-1
植物乳杆菌CCFM8610
乳酸片球菌(Pediococcus acidilactici strain BT36)
蜡样芽孢杆菌
植物乳杆菌CCFM8661
植物乳杆菌CCFM8610
扩展阅读:客观认识植物乳杆菌 (L. plantarum) 及其健康益处
罗伊氏乳杆菌P16
扩展阅读:认识罗伊氏乳杆菌(Lactobacillus reuteri)
短乳杆菌23017
➦ 下一代益生菌:
普拉梭菌Faecalibacterium prausnitzii 是一种可以生产短链脂肪酸能力的人类肠道共生菌,参与微生物的解毒作用。
扩展阅读:肠道核心菌属——普拉梭菌(Faecalibacterium Prausnitzii),预防炎症的下一代益生菌
➦ 重组细菌:
由于一些天然益生菌的不足,重组菌成为重金属污染微生物解毒研究的热点。重组乳酸乳球菌pGSMT/MG1363,产生一种类似于谷胱甘肽 S-转移酶的融合蛋白,具有解毒功能。将重组乳酸乳球菌菌株喂给新生大鼠后,血铅水平显著降低。
Pb结合结构域(PbBD)是已证明具有吸附Pb2+能力的蛋白质之一。通过构建大肠杆菌PbBD并将其引入草鱼体内,结果表明,大肠杆菌PbBD可通过与Pb2+()结合,来减少组织中的Pb积累。
益生菌解毒重金属的机制
与细胞壁上的多糖的金属络合,以降低重金属浓度,通过转运蛋白的重金属离子,与细胞内金属螯合蛋白结合形成沉淀,以及通过酶反应转化为低毒形式等。
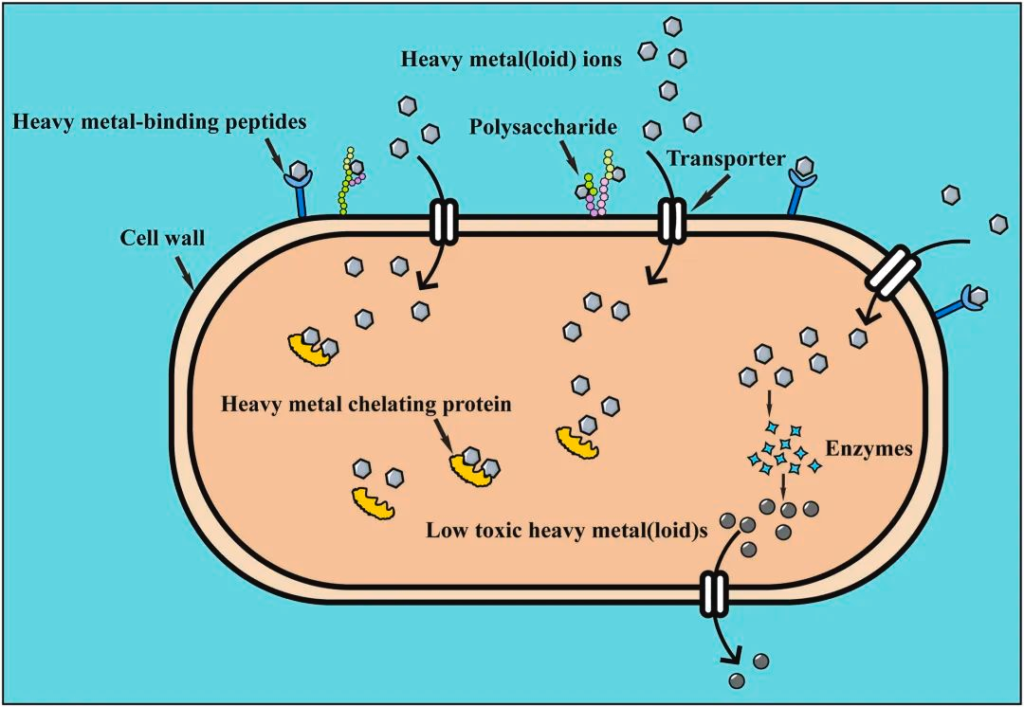
doi.org/10.1016/j.envpol.2022.120780
以上是第一种解毒策略,通过益生菌和重组细菌可以显著减少重金属对肠道微生物的影响,逐渐恢复健康的肠道菌群。接下来看第二张解毒策略:
✔ 肠道菌群防御反应的存在减少重金属的吸收
拮抗菌群的增加,可以减少宿主组织中镉的积累,从而降低发生一系列镉诱导疾病的风险。这种防御反应可能通过增加细胞外磷的浓度来增加铅的沉淀,从而降低肠道中的铅浓度。
——肠道微生物组可以直接影响砷代谢
在暴露于急性砷的小鼠肠道中,第一种反应被发现是一种防御机制,其特征是携带砷抗性基因或参与砷解毒机制的细菌数量增加。
动物研究也表明,肠道细菌通过将砷转化为粪便排泄,限制了砷在宿主体内的积累,降低了砷对宿主的影响。无机砷通过细胞膜上的磷转运蛋白或水通道蛋白进入生物体,并通过生物还原和生物甲基化进一步转化为各种其他形式的砷。
绝大多数无机砷的转化是由肠道菌群进行的,但肠道微生物对砷的处理在肠道的不同位置有所不同:
——肠道菌群抵抗铬吸收,有助于防止铬毒性
肠道微生物中的Cr(VI)耐受细菌,如副乳杆菌CL1107,在体外表现出Cr(VI)的减少,将Cr(IV)还原为Cr(III),其在中性或碱性条件下更难溶解。Cr(III)的低溶解度及其不能透过细胞,增加了粪便中铬的排泄量。
其他可能有用的措施
1、饮食
与排出重金属有关的食物包括:
螺旋藻、香菜、蓝莓等;
富含果胶的食物,如苹果、梨和柑橘类水果;
富含硫的食物,如西兰花、洋葱和大蒜等。
新鲜蔬菜榨汁:
绿叶蔬菜富含叶绿素,叶绿素在身体的解毒过程中非常重要,有助于清除血液中的重金属和毒素。同时,蔬果汁有利于增强肝功能,肝脏能够有效地处理和消除体内的毒素。
2、净化水源
例如:通过滤水器,帮助净化水中的重金属、有毒化学物质和污染物;或者饮用矿泉水等方式。
3、适量运动
出汗是一种自然的身体过程,可以帮助我们清除体内的毒素和废物。确保定期锻炼身体,让身体出汗排毒。
对于身体来说,药物是一种异源物质。身体可能会限制药物在胃肠道的吸收;加剧其在肠道和肝脏中的新陈代谢。
为什么不同人用药效果不一样?
药物明明是用于治病的,为什么可能带来所谓的副作用或者说毒性,甚至更严重后果?
肠道菌群是如何影响药物代谢,从而对身体带来不同影响?
用于神经系统疾病的药物,也与肠道菌群相关?
一个人服用多种药物,为什么会叠加健康风险?
本章节我们用常见的抗炎药,止泻药,癌症药物,神经系统药物,抗病毒药物来举例,从肠道菌群的角度来了解药物代谢的相关问题。
➤ 微生物对前药转化率不同,会影响患者疗效
抗炎药,尤其是针对胃肠道的抗炎药,由肠道细菌直接(化学修饰)或间接(通过微生物群和宿主细胞之间的相互作用)转化。此外,这些药物中的一些作为前药摄入,依赖微生物代谢将非活性前体转化为药物活性化合物。
肠道微生物将柳氮磺吡啶还原为磺胺吡啶后,柳氮磺嗪转化为活性抗炎剂5-ASA,然后进一步代谢为非活性N-乙酰基5-ASA。偶氮还原率和乙酰化率的差异,可以解释柳氮磺吡啶对患者的不同治疗效果。
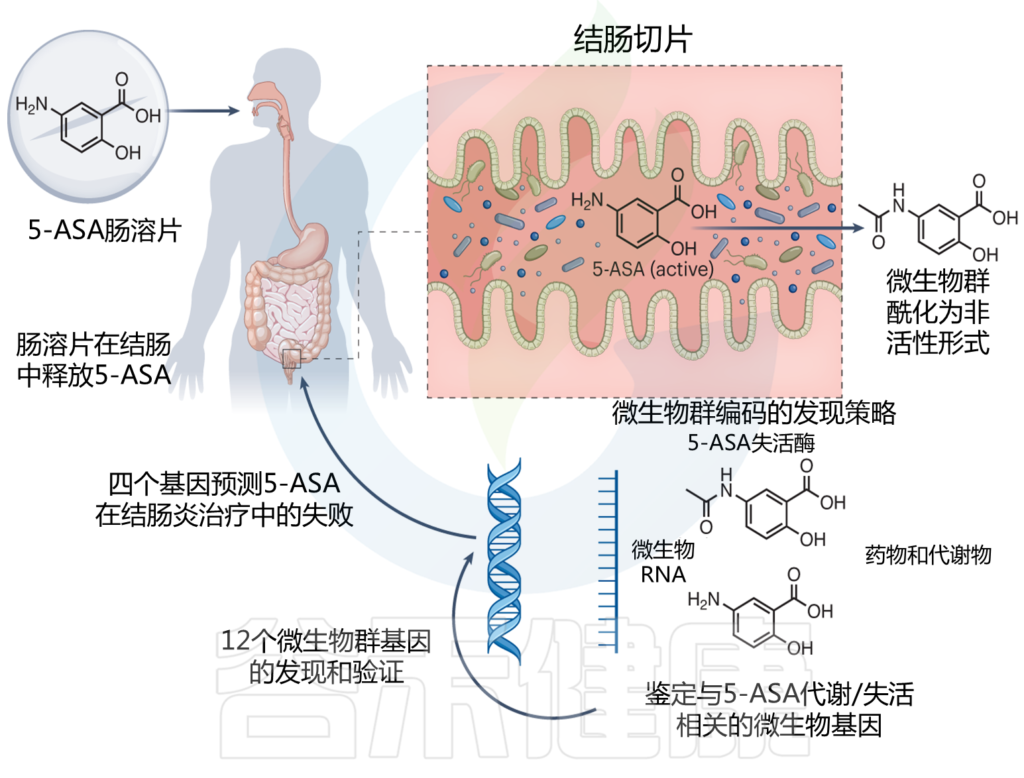
Macpherson AJ, et al., Nat Med. 2023
止泻药,洛哌丁胺的N-氧化物的减少也是肠道微生物活性激活前药的一个例子。
然而,对患者体内负责这些活动的特定生物和酶有更好的理解,有助于管理药物选择和给药等治疗。
个体之间对癌症化疗的治疗反应的一些差异,例如临床疗效和不良反应,也可能是由于肠道微生物群与药物相互作用的现象。肠道细菌可以直接改变化疗药物及其代谢产物的结构,从而改变与宿主细胞的相互作用。
➤ 癌症治疗:药物效果因人而异,可能与肠道菌群或肿瘤相关微生物对药物的结构改变有关
肠道菌群改变常用癌症药物的化学结构。在体外研究中,由于直接的化学修饰,大肠杆菌或魏氏李斯特菌(Listeria welshimeri)等细菌可以提高或降低30种抗癌药物的疗效。大肠杆菌也可以通过类似代谢活动来改变体内化疗的效果。
已知肠道微生物会降低化疗药物吉西他滨的活性浓度。大肠杆菌也可以阻止吉西他滨的疗效,因为它被微生物胞苷脱氨酶转化为其非活性代谢物。有意思的是,胞苷脱氨酶也作为嘧啶挽救途径的一部分存在于人类细胞中。因此,微生物酶在这种抗癌剂代谢中的真正影响仍然不确定。
➤ 神经药物作用于大脑,却也受控于肠道菌群
除了影响局部作用的药物外,肠道菌群代谢还会影响靶向远处器官系统的治疗效果。一些药物,如用于帕金森病治疗的口服左旋多巴,在穿过血脑屏障后,左旋多巴在大脑中被宿主酶局部代谢,以恢复多巴胺水平。然而,在到达大脑之前,药物在肠道中仍有广泛的代谢,导致浓度和患者反应发生显著变化。
一些短乳杆菌菌株,以及粪肠球菌、迟缓埃格特菌(Eggerthella lenta )能够在体外脱羧左旋多巴,但人类中大多数控制左旋多巴转化的肠道微生物和酶尚不清楚,需要更多的研究来揭示肠道菌群与各种神经药物之间的相互作用。
➤ 怎么看待同时服用不同种类药物,死亡风险升高?
拿抗病毒药物举例,如带状疱疹药物索罗夫定。据观察,服用索罗夫定的人,如果同时服用抗癌药物5-氟尿嘧啶,死亡风险会增加,这是为什么?
可能是因为索罗夫定溴戊酸脲的肠道微生物代谢产物减少,使二氢嘧啶脱氢酶失活,这反过来促进5-氟尿嘧啶的积累,导致更显著的毒性。
也可能与不同人群中酶的活性的显著差异有关。因此,同时服用这些药物带来药物不利影响的风险系数较高。
➤ 基于对肠道菌群药物代谢的了解,开发相应的方法提高疗效或减轻不良反应
地高辛主要用于治疗心力衰竭和心房颤动等心脏疾病,但是肠道菌群可以让它失活。地高辛具有狭窄的治疗窗口,用药过量会导致中毒反应,因此在临床上使用地高辛需要仔细监测以防止可能的心脏毒性。
当迟缓埃格特菌(Eggerthalla lenta)转化为没有治疗作用的代谢产物时,心力衰竭和心房颤动的治疗受到损害。
可能的机制与地高辛暴露时,迟缓埃格特菌中双基因细胞色素编码操纵子的上调有关。这种基因表达的蛋白质产物是Cgr1-Cgr2(心苷还原酶)复合物,它可以与地高辛结合并,将其还原为非活性代谢物。
既然我们了解到了这一点,那么有没有什么办法能阻止肠道菌群让地高辛失活?
研究人员认为,高蛋白饮食可以通过精氨酸防止肠道微生物群对地高辛的失活。也就是说饮食控制可以用来改变微生物的药物代谢,因为知道了细菌有独特的营养需求。不过随着对肠道细菌转化药物的机制了解的增加,也可以开发其他治疗方法来提高药物疗效或减轻不良反应。
所有这些研究都强调了揭示药物微生物转化的所有方面的重要性,可以从研究的药代动力学中明显地影响药物的作用。了解异源代谢过程是有必要的,这可能会影响药物的疗效、剂量和避免毒性。
草药在全世界范围内用于预防和治疗各种疾病已有数千年的历史。随着对天然药物的需求不断增长,中草药在世界范围内的重要性正日益增加。
肠道微生物群将中医药化学物质生物转化为代谢物,这些代谢物具有与其前体不同的生物利用度和生物活性/毒性。
接下来,我们从生物利用度和毒性这两方面,来详细了解,肠道菌群在中草药代谢中发挥什么样的作用,如何提高中草药的生物利用度,对毒性产生的正面或负面影响如何,用什么方法可以利用肠道菌群来降低中草药毒性等。
人在服用中草药后可分为两个阶段:
◗ 草药的生物利用度低,容易到达结肠与肠道菌群相遇
在草药中,许多成分的口服生物利用度非常低,因为它们是高极性化合物,亲脂性差。因此血流中的浓度水平有限。比如说,多酚的生物利用度通常低于 10%,人参皂苷的生物利用度可低至0.1%。
由于生物利用度低,这些化合物可以很容易地通过小肠并到达结肠。未被吸收的化合物就不可避免地与肠道菌群接触,并产生相互作用。
◗ 肠道菌群和中草药之间的相互作用是如何发生的?
首先,肠道微生物群与中草药之间的直接相互作用是双向的:
除了直接相互作用之外,中药与肠道菌群之间还会发生间接相互作用。在这类反应中,宿主的免疫和代谢系统起到了连接两端的桥梁作用。
免疫系统可以合成和释放一系列物质,如免疫球蛋白 A 和其他抗菌肽,可以调节肠道菌群组成。吸收的中草药可以调节胃肠道的免疫系统,最终导致肠道菌群发生变化。
比如说,一种中药化合物杜鹃花醇,可以改变牛乳腺上皮细胞中抗菌肽的表达,从而增强对金黄色葡萄球菌感染的防御能力。
◗ 在草药口服生物利用度低的问题中,肠道菌群能发挥作用吗?
出于患者方便和治疗依从性考虑,大多数药物是口服给药的。优化新药开发的主要特性之一是口服吸收度高。
糖苷,如三萜苷和黄酮苷,是许多草药含量中最常见的代表之一。由于氢键增加、糖部分的分子柔性和极性表面积,它们的肠道吸收有限。肠道微生物群可以将这些分子修饰成更亲脂性和极性更小的分子。
◗ 肠道菌群可以根据化合物的结构性质催化一系列代谢反应
草药中的糖苷首先在肠道中通过逐步水解(去糖基化和酯化)代谢。该过程产生的次生糖苷和/或苷元通常具有更好的肠道吸收和更好的生物利用度。它们通过骨架保留修饰(例如三萜苷)、骨架分裂(例如类黄酮苷)或骨架重排(例如环烯醚萜苷)进一步转化。
许多细菌门,其中拟杆菌门和厚壁菌门占优势,编码有丰富的糖苷水解酶基因,因此,它们用来进行糖苷水解。
◗ 不同肠道细菌可以合作处理单个化合物的代谢,单个细菌菌株也能够转化不同的化合物
梭杆菌K-60能使人肠道菌群中的槲皮苷去糖基化,而四种细菌菌株(Pedicoccus Q-5、链球菌S-3、拟杆菌JY-6、双歧杆菌B-9)参与了苷元(槲皮素)的进一步代谢。
丁酸梭菌通过各种反应机制,如脱糖基化、脱水、缩合、分子内环化和/或脱氢,能够转化属于不同结构类型的几种化合物,即紫草素、乌头碱和栀子苷。
然而说到中草药颇有争议,并不是所有人都认可,所谓“是药三分毒”,有人认为它会带来药物性肝损伤,这是一种常见的药物不良反应,可导致肝功能衰竭等问题。
近年来研究表明,中草药可以与肠道菌群相互作用,这种相互作用可以极大地影响其毒性和疗效。
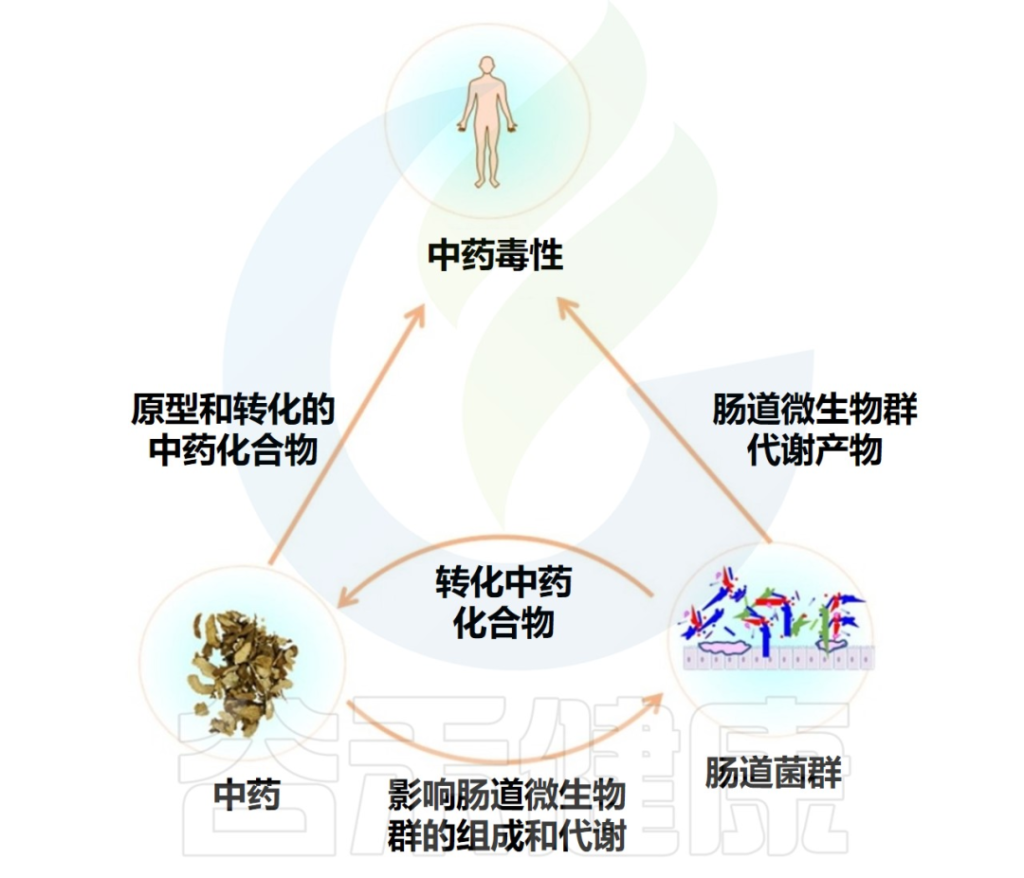
doi.org/10.1016/j.biopha.2020.111047
肠道菌群可以合成和释放一系列具有代谢异源物能力的酶,这些酶可以转化中药化合物,从而直接改变中药的毒性,这种改变包括增加毒性、减少毒性、解毒、激活毒性。
注:这些酶例如,β-葡萄糖醛酸酶、 β-葡萄糖苷酶、 β-半乳糖苷酶、α-鼠李糖苷酶等。
接下来从这四方面展开了解,肠道菌群对中药毒性的直接影响。
► 增加毒性
苦杏仁苷是一种在杏仁中发现的化合物,杏仁是一种用于治疗呼吸系统疾病如支气管炎、肺气肿的中药,但也可能有毒性,表现为恶心、呕吐、胸闷、头晕。
而肠道菌群可以水解苦杏仁苷的糖苷键,从而释放扁桃腈,扁桃腈是一种可以自发分解产生苯甲醛和有毒氢氰酸的化合物。因此,肠道菌群对苦杏仁苷毒性的修饰具有重要作用。
除了苦杏仁苷外,肠道菌群也可以代谢熊果苷、葛根素【与消化链球菌属 (Y-10)有关】、大豆苷【与双歧杆菌属 (K-111)有关】等其他中药成分,产生毒性更强的代谢物。
► 降低毒性
乌头(附子)的侧根是临床广泛用于治疗心力衰竭的中药,但常对神经和心血管系统产生毒性作用。
乌头生物碱是影响附子药理活性和毒性的主要化合物。在临床上有个办法来降低毒性,附子通常经过数小时的煮沸处理,然后再给患者开处方。
在胃肠道中,肠道菌群可以通过脱酰、酯化和去除甲基羟基以类似的方式转化乌头生物碱,参与的菌群与脆弱拟杆菌、肺炎克雷伯菌、丁酸梭菌有关。因此,肠道菌群降低了其毒性。注意肠道的这种减少的程度是有限的,口服大剂量依然会导致急性毒性。
除了乌头生物碱之外,肠道菌群还会对黄芩苷、对羟基苯甲酸丁酯转化为毒性较小的化合物。
► 解毒
这里的排毒,和前面章节的药物代谢机制类似,将活性中药化合物转化为非活性产物。
草酸盐是一种草药化合物,存在于许多中药中,能够诱发高草酸尿症、肾结石和心脏传导障碍。肠道菌群可以在甲酰辅酶 A转移酶和草酰辅酶 A脱羧酶的帮助下,解毒草酸盐。
► 激活毒性
这与前面解毒相反,激活是指肠道菌群将无毒化合物转化为有毒化合物。
苏铁甙是苏铁科植物中的一种偶氮糖苷,具有肝毒性和致癌能力。肠道菌群可以将苏铁甙转化为其苷元(甲氧基甲醇),苷元可以进一步转化为重氮甲烷,一种具有肝毒性和致癌作用的有毒化合物。
中药与肠道菌群的直接相互作用,不仅会导致中药化合物的转化,还会导致肠道菌群的组成和代谢发生变化,从而进一步影响中药的毒性。
以上我们可以看到,肠道菌群对毒性的影响并不是固定的,可能是增加毒性或者减小毒性。那么当发现某些肠道菌群可能会增加中草药毒性时,我们能做些什么尽可能阻止毒性增加呢?
由于肠道微生物群在调节中药毒性方面的重要作用,靶向肠道微生物群已成为控制中药毒性的新前沿。这里总结了靶向肠道菌群降低中药毒性的可能方法。
1、炮制减毒
许多中药在用于临床或制造中药之前要经过煮沸、漂、蒸等特殊处理,也就是“炮制”。
炮制的增效和降毒作用机制是中药现代化的重要问题。研究表明,煮沸等炮制可以将有毒化合物转化为无毒或毒性较小的化合物,从而改变中药的毒性,如前面说的乌头根。
近年来,研究人员证明炮制可以影响肠道微生物群,从而影响中药的药理作用。
甘遂是一种有毒的中药,用于治疗水肿、腹水和哮喘。由于其强烈的毒性,如肝脏和胃肠道毒性,通常用醋煎烤以降低毒性。最近的一项研究表明,醋制甘遂显著减少了甘遂中的10种有毒化合物,如甘遂素B、甘遂素C和甘遂素E。同时,醋制甘遂可以显著改变肠道微生物群的组成,显著提高了短链脂肪酸水平。
注:醋制是是中药炮制中重要的炮制方法。
考虑到肠道微生物群产生的短链脂肪酸具有抗炎和肠粘膜保护作用,可以推断醋制可以调节肠道微生物群的组成和代谢,从而降低甘遂的毒性。
2、中药组合
肠道微生物群释放的β-葡萄糖醛酸酶是一种重要的酶,可以将草药中的糖苷水解为苷元,这一过程可以增加草药化合物和药物的毒性。
非甾体抗炎药和β-葡萄糖醛酸酶抑制剂等化学药物的组合可以降低化学药物的毒性。
一些中药化合物(如黄芩苷)可以抑制细菌β-葡萄糖醛酸酶的活性,其与化学药物(如CPT-11)的组合可以降低CPT-11的毒性。
槲皮素的细菌代谢产物3,4-二羟基苯乙酸可以减少对乙酰氨基酚引起的肝损伤。因此,可以推断,具有抑制细菌酶的能力的中药可以与其他有毒中药或化学药物结合,以降低其毒性。
3、饮食
我们已经知道,饮食在调节肠道微生物群组成和功能中具有重要作用。同样,研究人员也想办法寻找饮食和药物之间的关联。上一章节我们已经了解到,高蛋白饮食可以抑制地高辛失活,并可用于降低某些患者(原本要更高剂量的患者)的地高辛毒性风险。
4、其他(FMT)
其他方法包括粪便微生物群移植(FMT)、益生元和益生菌的补充以及肠道微生物群的工程都有可能降低药物的毒性。
粪菌移植可以显著改善难治性免疫检查点抑制剂诱导的结肠炎,还可以显著改善FOLFOX(一种含有5-氟尿嘧啶、亚叶酸和草酸铂的方案)诱导的大肠癌癌症动物模型中的肠粘膜炎和腹泻。
预防性服用一种益生菌菌株干酪乳杆菌鼠李糖亚种,可显著降低FOLFOX诱导的结肠癌症动物模型中的肠道粘膜炎和腹泻,但不会改变FOLFOX的抗肿瘤作用。
TGF-β阻断剂和大肠杆菌Nisle 1917的组合增强了Galunisertib的肿瘤抑制作用。
肠道微生物群工程是直接治疗疾病和改善药物疗效的新兴前沿,如苯丙酮尿症和左旋多巴。尽管这些方法尚未用于控制中草药的毒性,但可以预见,这些方法将在不久的将来会引入,从而改善中草药毒性。
➭ 肠道微生物组、宿主因素和异源代谢的多方面相互作用是复杂的。微生物异源代谢可导致生物活化、解毒,通过加强肠粘膜屏障来防止吸收。肠道微生物的各种酶反应拓宽了我们对外来生物如何代谢的看法,代谢产物可以发挥新的活性。
➭ 破译异源物的肠道微生物转化,特别重要的是从患者处获取关于饮食或外源性暴露的广泛信息,有助于其与特定健康结果联系起来。
准确评估异源物质引起的肠道微生物群扰动,是将肠道微生物群失调与宿主健康状况联系起来和开发微生物群导向疗法的前提和基础。
➭ 重金属等环境污染物的暴露对肠道微生物群的扰动,会影响代谢和生理功能,部分导致代谢疾病的病因或进展。当然还需要更多深入、系统的研究,来确定重金属等污染物暴露对肠道微生物群的和时间效应。
➭ 了解肠道菌群对药物毒性和疗效的调节作用背后的分子机制,有助于改进药物开发和精准医学,更有效地评估风险,并开发个性化营养。
在不同的临床环境中为个体指导个性化营养膳食及肠道干预,或者制定相应益生菌补充方案,是提高中草药价值的重要研究方向。
将肠道微生物如何转化药物的知识整合到药物开发和实施的所有阶段,将有助于设计合适的治疗方案并改善患者对药物的临床反应。
主要参考文献
Liu X, Zhang J, Si J, Li P, Gao H, Li W, Chen Y. What happens to gut microorganisms and potential repair mechanisms when meet heavy metal(loid)s. Environ Pollut. 2023 Jan 15;317:120780. doi: 10.1016/j.envpol.2022.120780. Epub 2022 Nov 29. PMID: 36460187.
Mehta RS, Mayers JR, Zhang Y, Bhosle A, Glasser NR, Nguyen LH, Ma W, Bae S, Branck T, Song K, Sebastian L, Pacheco JA, Seo HS, Clish C, Dhe-Paganon S, Ananthakrishnan AN, Franzosa EA, Balskus EP, Chan AT, Huttenhower C. Gut microbial metabolism of 5-ASA diminishes its clinical efficacy in inflammatory bowel disease. Nat Med. 2023 Feb 23. doi: 10.1038/s41591-023-02217-7. Epub ahead of print. PMID: 36823301.
Koppel N, Maini Rekdal V, Balskus EP. Chemical transformation of xenobiotics by the human gut microbiota. Science. 2017 Jun 23;356(6344):eaag2770. doi: 10.1126/science.aag2770. PMID: 28642381; PMCID: PMC5534341.
Lindell AE, Zimmermann-Kogadeeva M, Patil KR. Multimodal interactions of drugs, natural compounds and pollutants with the gut microbiota. Nat Rev Microbiol. 2022 Jul;20(7):431-443. doi: 10.1038/s41579-022-00681-5. Epub 2022 Jan 31. PMID: 35102308.
Macpherson AJ, Sauer U. Secrets of microbiota drug metabolism. Nat Med. 2023 Feb 23. doi: 10.1038/s41591-023-02227-5. Epub ahead of print. PMID: 36823300.
Zhang Q, Bai Y, Wang W, Li J, Zhang L, Tang Y, Yue S. Role of herbal medicine and gut microbiota in the prevention and treatment of obesity. J Ethnopharmacol. 2023 Apr 6;305:116127. doi: 10.1016/j.jep.2022.116127. Epub 2023 Jan 2. PMID: 36603782.
Feng W, Liu J, Huang L, Tan Y, Peng C. Gut microbiota as a target to limit toxic effects of traditional Chinese medicine: Implications for therapy. Biomed Pharmacother. 2021 Jan;133:111047. doi: 10.1016/j.biopha.2020.111047. Epub 2020 Dec 4. PMID: 33378954.
Chen F, Wen Q, Jiang J, Li HL, Tan YF, Li YH, Zeng NK. Could the gut microbiota reconcile the oral bioavailability conundrum of traditional herbs? J Ethnopharmacol. 2016 Feb 17;179:253-64. doi: 10.1016/j.jep.2015.12.031. Epub 2015 Dec 23. PMID: 26723469.
Collins SL, Patterson AD. The gut microbiome: an orchestrator of xenobiotic metabolism. Acta Pharm Sin B. 2020 Jan;10(1):19-32. doi: 10.1016/j.apsb.2019.12.001. Epub 2019 Dec 10. PMID: 31998605; PMCID: PMC6984741.
Feng Y, Cao H, Hua J, Zhang F. Anti-Diabetic Intestinal Mechanisms: Foods, Herbs, and Western Medicines. Mol Nutr Food Res. 2022 Jul;66(13):e2200106. doi: 10.1002/mnfr.202200106. Epub 2022 May 11. PMID: 35481618.
Teschke R. Aluminum, Arsenic, Beryllium, Cadmium, Chromium, Cobalt, Copper, Iron, Lead, Mercury, Molybdenum, Nickel, Platinum, Thallium, Titanium, Vanadium, and Zinc: Molecular Aspects in Experimental Liver Injury. Int J Mol Sci. 2022 Oct 13;23(20):12213. doi: 10.3390/ijms232012213. PMID: 36293069; PMCID: PMC9602583.
谷禾健康

肠道微生物群之间编织了一个复杂的相互作用网络,影响人体的营养吸收和代谢,免疫功能等,对我们的健康状态有很大的影响。
我们知道,肠道微生物群具有多样性,平衡性,稳定性等特征,但同时也具有异质性,也就是说不同个体之间存在差异。不同人群对食物,膳食补充剂的健康需求不同。
特定的营养素、食物、整体饮食结构等诸多因素都会影响特定的肠道菌群,从而影响整体健康。
随着当前研究技术的不断进步,我们对肠道菌群和健康之间的关系认知不再停留在初级关联阶段,而是逐步走向精细化调节层面。
怎样通过这些看起来日常的饮食,益生菌补充剂等方式,去操纵肠道菌群以达到获得健康的目的,是一个值得深入探讨的话题。
本文,我们从肠道菌群的角度来了解一下,饮食模式、发酵食品、益生菌、益生元和其他相关化合物对我们健康的影响。
本文主要从五个方面讲述
●发酵食品对肠道菌群和健康的影响
●不同饮食模式与菌群及健康
●益生菌调节肠道微生物群和人体健康
●益生元化合物对肠道菌群的影响
●后生元和精神生物制剂与健康
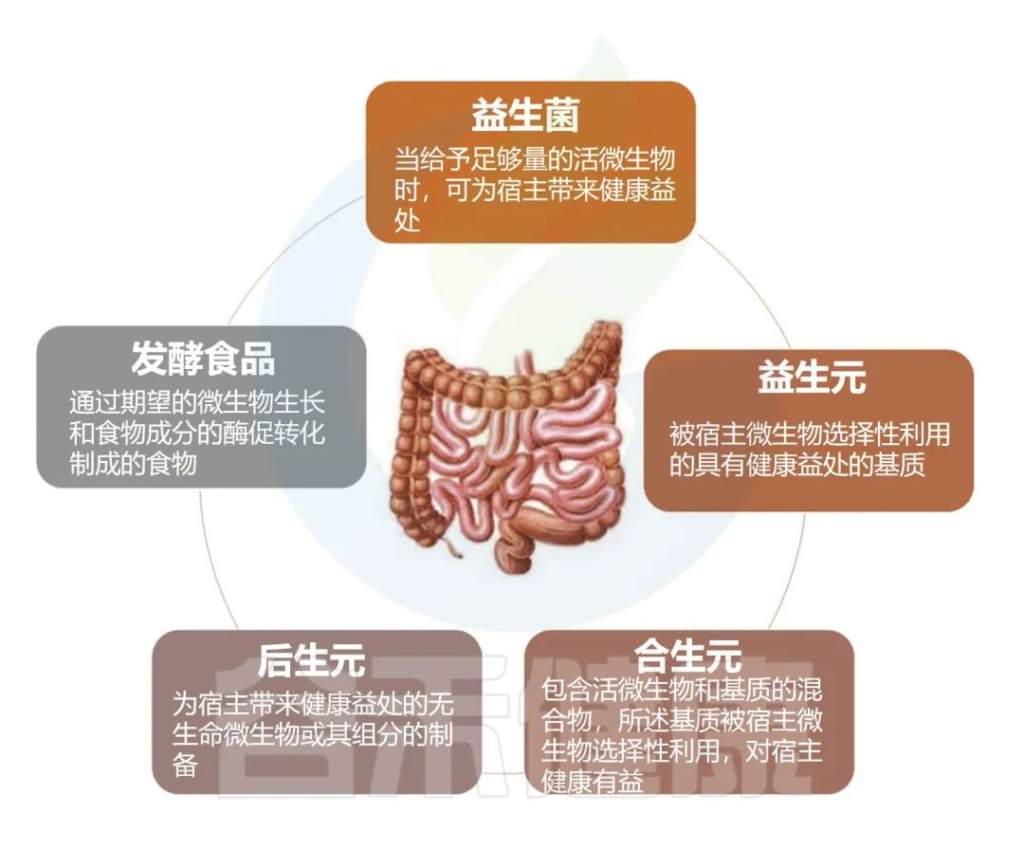
Nma B,et al.Elsevier Inc.2022
发酵食品被定义为“通过理想的微生物生长和食物成分的酶促转化制成的食品”。
许多发酵食品已被证明通过不同的方式对人体具有促进健康的作用,包括生物活性分子的合成、肠道微生物群的调节以及与免疫系统的相互作用。
✦发酵食品产生多种代谢物,促进有益作用
在食品发酵过程中,微生物进行酶促转化,提供多种具有不同生理活性的代谢产物(多肽、低聚糖、游离氨基酸、改性多酚、有机酸等)。
这些转化发生在食物摄入之前,发酵食品也是微生物进入我们肠道的绝佳载体,与我们的微生物群的其他成员一起,可以转化和代谢我们肠道中的食物成分,产生具有不同功能的微生物代谢物(短链脂肪酸、维生素、细菌素等)。
据报道,某些发酵食品或其成分能够以某种方式促进有益的效果,而这些效果有时与我们微生物群的变化有关。在本节中,我们将讨论发酵食品对微生物群的影响及其可能的有益作用。
发酵乳制品对健康的影响已经被深入研究了一个多世纪,但直到最近,随着新的组学技术和大规模测序方法的使用,我们才能够详细了解食用发酵乳制品对微生物群的影响。
1 酸奶

•酸奶有助于改善乳糖消化不良
酸奶是研究最多的发酵乳制品之一。它是通过两种乳酸菌(德氏乳杆菌保加利亚亚种和嗜热链球菌)在牛奶中的联合活性获得的。欧洲食品安全局已经认识到酸奶发酵剂的有益作用,认为活酸奶培养物可以改善乳糖消化不良患者的乳糖消化。
•饮用酸奶增加了嗜热链球菌,减少了拟杆菌丰度
一些动物模型的临床前研究表明,长期摄入酸奶改变了小鼠菌群中拟杆菌门(Bacteroidetes)和厚壁菌门(Firmicutes)之间的比例,并增加了链球菌科(Streptococcaceae)成员的数量,这可能是由于摄入了酸奶中存在的嗜热链球菌(S. thermophilus)。
此外,一些在人体中进行的干预试验表明,饮用酸奶会增加人体微生物群中的一些细菌数量:其中,乳酸菌的丰度较高,而拟杆菌(Bacteroides)的种类则相应减少。
2 奶酪

奶酪,也是一种发酵的牛奶制品,其性质与酸奶有相似之处,都是通过发酵过程来制作的,也都含有可以保健的乳酸菌,但是奶酪的浓度比酸奶更高,近似固体食物,营养价值也因此更加丰富。
•食用奶酪可以增加产丁酸盐细菌丰度,可能缓解特应性皮炎
研究了奶酪和乳制品引发肠道微生物群变化并促进相关健康影响的能力。最近进行了一项健康食品选择与肠道微生物群组成之间的广泛关联研究,表明α多样性与低脂奶酪之间存在强烈的正关联。
临床前研究报告称,给小鼠服用奶油奶酪后,产生丁酸盐的细菌水平增加,T细胞(Treg)介导的免疫反应和IgE水平降低,表明对缓解特应性皮炎有潜在的有益作用。
•奶酪中的发酵菌可以存在人体一段时间
在人类中,两项干预试验评估了食用卡门贝尔奶酪对不同肠道菌群成员的影响,结果表明,食用奶酪后粪肠球菌(Enterococcus faecalis)的丰度显著增加,并且卡门贝尔奶酪中存在的微生物,如乳酸乳球菌(Lactococcus lactis)、肠膜明串珠菌(Leuconostoc mesenteroides)和白地土菌(Geotricum candidum),在食用奶酪期间出现在个体的粪便中。在停止干预15天后,粪便样本中存在肠膜明串珠菌。
3 开菲尔
开菲尔是一种由微生物混合物发酵的产品,通常包括几种乳酸菌和酵母,其发酵和生产过程根据其生产的地理区域和传统的细化过程而显著变化。
因此,很难对开菲尔的效果得出一般性的结论,更谨慎的做法是根据所研究的开菲尔的类型来具体说明这些效果。
•改善机体代谢,抗肥胖
有证据表明,食用开菲尔可能有有益的代谢作用。开非尔能够调节啮齿动物体内的微生物群组成。向肥胖的人类微生物群相关大鼠喂食西藏开菲尔牛奶表明,微生物群变化与胆汁酸和氨基酸代谢有关。
此外,连续四周每天给小鼠服用克非尔显示了对厚壁菌门/拟杆菌门比率的影响,以及对性能和身体疲劳的改善。
另一项研究通过建立社区特征和几种生物标志物之间的相关性,证明了开菲尔在小鼠体内的抗肥胖作用,这表明食用开菲尔诱导的微生物群和真菌群调节可能与预防肥胖和脂肪酸代谢有关。
•改善炎症性肠病、代谢综合征
一项针对炎症性肠病患者的随机对照试验表明,定期食用开非尔可以改善生活质量,减少腹胀。
此外,一项平行组随机对照干预试验显示,代谢综合征患者接受开菲尔12周后,在开菲尔组中放线菌的相对丰度增加,对代谢综合征标志物有一些有利的影响。
•还可能影响宿主行为和免疫状态
对小鼠施用开非尔还被证明可以增加短乳杆菌(Lactobacillus brevis)的流行率和微生物群产生γ-氨基丁酸的能力,影响宿主行为和免疫状态。
这些临床前证据表明,开非尔对宿主的微生物群和代谢有影响,包括肥胖、糖尿病、肝脏和心血管疾病、免疫和神经系统变化。
除了发酵乳制品外,关于其他动物性发酵食品调节微生物群的信息较少。然而,有证据表明植物发酵食品对肠道微生物群的影响,以及其与健康影响的潜在关系。
在这方面,酸菜、泡菜、红茶菌和其他发酵的植物性食物已经被研究过,并显示出对肠道微生物群的不同影响,尽管需要更多的证据来建立它们对人类健康的明确联系。
1 酸面包
•低蛋白饮食相关细菌减少、影响升糖反应
酸面包是一种发酵食品,也显示出了显著的效果。采用元蛋白质基因组学方法分析了酸面包对大鼠微生物组的影响,结果显示低蛋白质饮食相关的细菌类群减少。
在另一项研究中,食用手工酸面包引起的升糖反应与食用工业白面包不同。这些反应是个体特异性的和微生物相关的。
每个人的血糖反应可以根据干预研究前确定的微生物组谱进行预测。
2 发酵豆制品
•促进脂肪酸分解代谢
关于发酵豆制品,一些临床研究显示了微生物群调节活性和各种生理效应。在啮齿动物中研究了大豆发酵产品对代谢过程的影响,表明发酵大豆能够促进小鼠脂肪酸分解代谢和主要细菌门的变化。
•影响其他代谢物水平
鼠李糖乳杆菌发酵的豆浆通过增加一些细菌类群,如拟杆菌(Bacteroides),对小鼠微生物群有影响,这是一种通过尿液排泄的异黄酮代谢物的来源。
此外,用解淀粉芽孢杆菌(Bacillus amyloliquefaciens)发酵的大豆降低了2型糖尿病大鼠模型中的高血糖,引起疣微菌(Verrucomicrobiales)种群的增加和肠杆菌(Enterobacteriales)的减少,以及其他微生物群的变化。
•改善认知功能
值得注意的是,在小鼠中摄入植物乳杆菌发酵的大豆后,观察到对认知功能的积极影响,以及乳酸菌和双歧杆菌种群的增加。
含有植物乳杆菌(L.plantarum C29)的发酵产品在一组轻度认知障碍患者中进行的为期12周的人类临床试验中显示出认知能力的改善。
•可能改善皮肤状况
最后,摄入含有干酪乳酸菌代田株(Lactobacillus casei Shirota)的发酵豆乳对健康绝经前日本女性的皮肤状况有显著影响,乳酸菌和双歧杆菌的数量有增加的趋势,而肠杆菌科和紫单胞菌科(Porphyromonadaceae) 的数量则有下降的趋势。
由于各种食物已经证明了调节特定细菌和肠道微生物群整体结构的能力,因此短期和长期的饮食模式可以影响肠道微生物组的构成和良好功能。
★ 饮食对肠道菌群的多样性至关重要
我们肠道菌群的多样性可能反映了我们饮食的多样性,因此,饮食可能是我们控制和平衡肠道菌群组成和代谢的最强大的武器。
以下部分旨在概述过去几年来产生的关于不同饮食和饮食习惯对调节肠道菌群的潜力的一些现有知识。
西式饮食、地中海饮食和素食这几种可能是世界上最常见的人类饮食模式,每一种都与一些特定的健康/疾病状况有关。
✦不同饮食模式的差异主要是碳水化合物和纤维的含量
宏量和微量营养素方面的显著差异决定了这些饮食模式,其中复合碳水化合物和纤维的含量可能是最显著的之一。
事实上,复杂的碳水化合物和纤维可能是产生最确凿证据的食物成分,它们是肠道菌群的有益调节器,在碳水化合物消耗仅24小时后就会发生快速转变。
西式饮食,是一种以高含量精加工糖和碳水化合物、高含量饱和脂肪酸、高含量动物蛋白以及低含量膳食纤维为特征的一种现代饮食方式,不能否认的是这种饮食好吃还容易上瘾。
✦西式饮食导致易患许多代谢疾病
这种饮食模式在人类历史上是最近才出现的,通常与城市生活方式有关,通常与炎症和代谢疾病有关,包括2型糖尿病和肥胖等。
✦西式饮食下的肠道微生物显著改变
此外,在将祖先生活方式和农业人口的饮食与西方化生活方式的饮食进行比较时,观察到肠道微生物群的组成发生了显著变化,总体特征是纤维降解细菌的减少和蛋白质代谢细菌的增加。
变形菌、拟杆菌等蛋白质代谢菌较丰富
最近的研究已经指出,与这些生活方式相关的一些疾病生物标志物可能是通过西方饮食对肠道微生物群的干扰作用介导的,其特点是多样性低。
蛋白质和胆汁代谢细菌占主导地位,包括产生三甲胺n-氧化物(TMAO)的物种,变形菌和拟杆菌通常更为丰富。
事实上,在动物模型中,西方化饮食的引入会迅速转化为微生物群的变化,并增加感染和代谢疾病的风险,在向新动物移植不良微生物群后,这些表型可以重现。
这表明,饮食模式影响人类健康的一些机制是通过调节肠道微生物群来实现的,并表明通过饮食调节肠道微生物群可能会降低一些疾病风险。
地中海饮食的特点是大量摄入新鲜水果、蔬菜、豆类和全谷物,同时摄入少量动物蛋白、加工食品和饱和脂肪。
✦地中海饮食改善代谢、降低慢性病患病率
长期以来,这种饮食习惯与改善代谢和心血管健康、健康老龄化、降低死亡率和慢性疾病患病率有关。
纤维降解细菌丰度增加、短链脂肪酸增加
其中一些影响与肠道菌群特征及其相关代谢组和免疫调节的特定变化有关。就地中海饮食对肠道菌群的具体影响而言,它通常与纤维降解物种的高度多样性和代表性有关,如普雷沃氏菌(Prevotella)和毛螺菌属(Lachnospira)。
其代谢导致短链脂肪酸的产生;以及一些瘤胃球菌的减少。事实上,严格坚持地中海饮食的人的短链脂肪酸水平似乎更高。
值得注意的是,与地中海饮食相关的有益肠道菌群调节并不完全与其长期食用有关,因为饮食习惯向这种模式转变,即使是相对较短的时间,通常也会改善微生物和健康生物标志物,这在各种炎症、代谢和认知障碍的动物模型以及人类临床试验中都得到了证明。
例如,对一组超重和肥胖的参与者进行了8周的地中海饮食干预,即使在保持能量摄入的情况下,也导致微生物群多样性显著增加,纤维降解细菌的表现包括普拉梭菌、一些拟杆菌属和一些罗氏菌属、颤螺菌属和毛螺菌,这些细菌负责短链脂肪酸的产生,包括丁酸盐生产者。其中一些还被认为具有抗炎特性。
✦有效改善炎症
此外,在饮食干预后,潜在的促炎细菌(如Ruminococcus gnavus)减少,胆固醇水平、炎症标志物和胰岛素抵抗全面改善。
地中海饮食对肠道菌群结构的好处也在其他健康受试者的队列中被揭示出来。在欧洲各国的老年人群中进行了一项大型饮食干预试验,表明在12个月的时间内坚持地中海饮食模式有利于调节微生物群中的关键物种,导致与较低虚弱和改善认知功能标志正相关的类群的丰富,并与炎症负相关,包括以下几种菌属:
普拉梭菌(Faecalibacterium);
罗氏菌属(Roseburia);
优杆菌属(Eubacterium);
多型拟杆菌(Bacteroides thetaiotaomicron);
普雷沃氏菌(Prevotella)。
素食
素食是一种不食肉等动物产品的饮食方式,有时也戒食奶制品和蜂蜜。在另一个极端,许多研究评估了长期素食和杂食饮食对微生物群、代谢组和疾病风险的影响。
✦素食对健康促进的具体作用暂不明确
长期以来,素食饮食一直被认为具有促进健康的作用,但与杂食性饮食相比,素食饮食也可以培养不同的肠道微生物群结构,总体上增加了多样性和丰富性;拟杆菌(Bacteroidetes)经常以较高的相对频率出现。
然而,报告了一些相互矛盾的结果,一些控制良好的喂养研究发现,生活在同一地理区域的杂食性和素食者之间的肠道微生物群只有适度的差异。不过两组在代谢组中都表现出很大的差异,这可能部分归因于微生物群产生的代谢物。
低FODMAP饮食
还有一种与潜在微生物群调节能力相关的饮食模式是低FODMAP饮食。
什么是FODMAP?
FODMAP是一组人体吸收较差的短链碳水化合物,包括果糖、果聚糖、乳糖、多元醇及半乳糖寡糖等。
其特点是可发酵低聚糖、双糖、单糖和多元醇的含量降低,虽然其中一些成分可能作为益生元对健康受试者的肠道菌群进行有益的调节,但它们也可能引发肠易激综合征(IBS)和其他肠道疾病患者的不良反应。
✦缓解肠易激综合征
低FODMAP饮食长期以来可以证明缓解了肠易激综合征患者的症状,但其有益效果似乎强烈依赖于患者的基础微生物群特征。
✦影响健康促进有关细菌的丰度
在微生物区系水平上,FODMAP消耗的减少通常会导致一些与健康促进有关细菌的肠道减少,如:
普拉梭菌(Faecalibacterium Prausnitzii)↓↓↓
双歧杆菌(Bifidobacterium)↓↓↓
嗜黏蛋白阿克曼菌(Akkermansia)↓↓↓
巨球型菌属(Megasphaera)↓↓↓
片球菌属(Pediococcus)↓↓↓
放线菌(Actinobacteria)↓↓↓
以及产生丁酸盐的细菌减少
同时伴随着其他气体消耗细菌的增加,如雌马酚产生者Adlercreutzia,甚至是Ruminococcus torques。细菌通常在肠易激综合征患者中检测到高丰度。
因此,低FODMAP饮食可以导致症状的改善,尽管所涉及的机制尚不清楚,但是同时,也可以诱导更明显的肠道菌群失调。
有趣的是,低FODMAP饲粮降低了结肠pH值和细菌总数,而短链脂肪酸浓度不受影响,这进一步支持了低FODMAP饮食下肠道菌群代谢中存在重要的重组。
注意
虽然人们普遍认为地中海和素食饮食可以改善肠道微生物群的多样性和纤维降解细菌的表现,但在不同的研究中,一些具体的类群有所不同,这阻碍了在饮食模式、微生物群变化和健康反映之间建立具体的因果关系。
这可以归因于不同食物组合的可变影响,以及对饮食干预的反应可能高度依赖于基础肠道微生物群组成,以及其他个体遗传和环境特征、宿主遗传、季节、地理和文化差异。
在考虑将饮食干预作为精确调节肠道微生物群的手段时,另一个需要考虑的关键方面不仅是宏观和微量营养素的具体组成,还包括摄入次数、饮食行为和饮食干预的持续时间。
一般来说,虽然相对短期的饮食干预足以对肠道菌群产生可测量的影响,但在恢复习惯性饮食后,这些变化会迅速恢复,甚至,根据基础菌群配置,可能需要更大的干预才能产生明显的效果;建议永久性的饮食适应,以确保提供持续的基质来源,为有益的肠道种群提供燃料及其相关的长期益处。
因此,虽然在该领域已经取得了重大进展,但为了建立旨在通过调节肠道微生物群来改善人类健康的个性化饮食策略,有必要进行进一步的研究。
益生菌被定义为“当施用足量时,对宿主的健康有益的活微生物”。
益生菌已被认为在不同水平上发挥其有益功能,包括:
•营养物质的代谢,以促进消化,产生维生素或具有全身作用的分子;
•神经系统信号的改变;
•免疫调节的诱导;
•对生理应激的保护;
•直接和间接的病原体拮抗;
•改善肠道上皮的屏障功能;
•调节微生物
益生菌可以在不同程度上对宿主产生有益作用
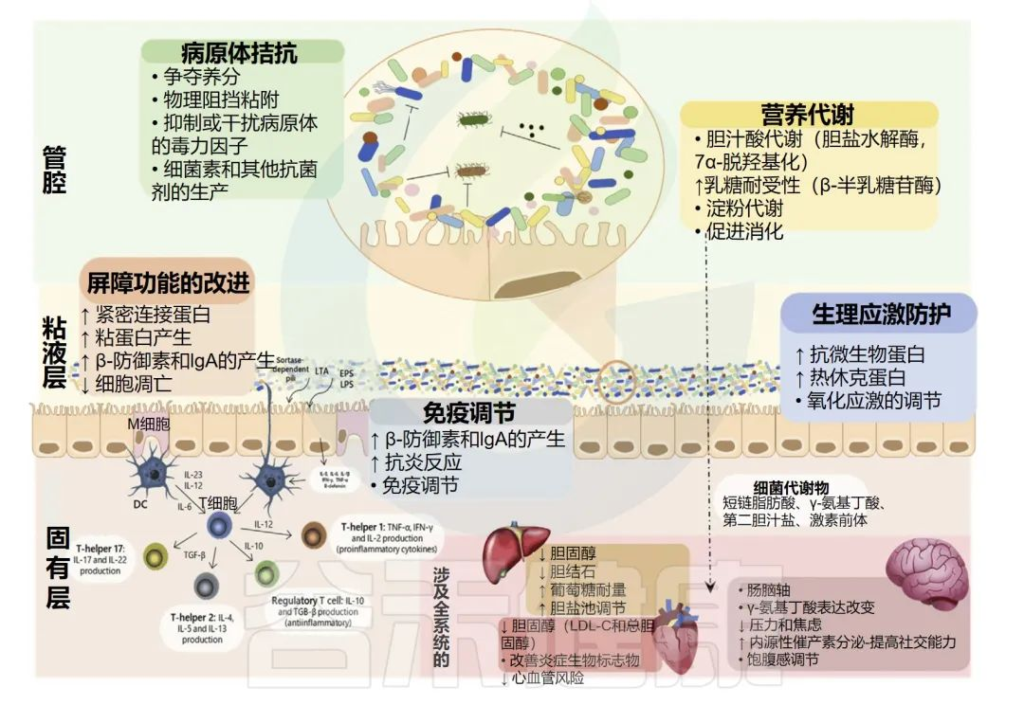
Nma B,et al.Elsevier Inc.2022
这些功能可以是接触依赖的或由表面分子介导的,如脂磷酸和梭酸依赖菌毛,或由分泌分子介导的,如短链脂肪酸和细菌素。
总的来说,突出了免疫调节和病原体拮抗活性,它们对于维持肠道微生物组中细菌群落与宿主之间的平衡至关重要。
一些研究,不仅在体外和动物模型中,而且在人类志愿者中,分析了益生菌对与人类疾病相关的特定细菌病原体或群体的抑制作用,如产气荚膜梭菌 (Clostridium perfringens)、肠炎沙门氏菌(Salmonella enteritidis)和大肠杆菌(Escherichia coli);以及其对生理或炎症标志物和免疫特征的影响。
益生菌也被认为能够调节整体肠道微生物群。事实上,有人提出所需的益生菌效果与肠道中的微生态变化有关。
✦益生菌治疗改变了肠道微生物组成
对动物模型的不同研究表明,用不同菌株的乳酸菌和双歧杆菌以及其他微生物(如布拉氏酵母菌)进行治疗,明显影响小鼠肠道的微生物群组成,促进微生物群落的结构和功能的变化。
注:这些研究大多使用细菌计数或qPCR分析来研究肠道微生物群分布的变化,只获得了生态系统的一小部分图像。在这方面,随着下一代测序技术和生物信息学工具的发展,我们可以对肠道微生物组进行更深入的研究,不仅可以从结构和组成层面,还可以从功能层面深入研究益生菌对微生物群落的影响。
缓解肥胖相关的肠道微生物失调
在动物模型上报道了鼠李糖乳杆菌、嗜酸乳杆菌、植物乳杆菌、两歧双歧杆菌、长双歧杆菌和粪肠球菌对饮食诱导的肥胖小鼠肠道微生物群的调节作用,缓解了饮食诱导的肥胖和相关的肠道微生物群失调。
此外一些益生菌,如茯砖茶中的冠突散囊菌(Eurotium cristatum),也显示出调节肠道真菌和细菌群落的能力,表现出与健康动物中观察到的情况更接近的特征,并减轻饮食引起的肥胖症状。
下面是益生菌调节肠道菌群的一些证据:
调节抑郁症的肠道生态失调
肠道生态失调
在抑郁症小鼠模型中,使用含有L.plantarum LP3、L.rhamnosus LR5、Bifidobacterium lactis BL3、Bifidobacterium breve BR3和Pediococcus pentosaceus PP1的益生菌配方进行干预,可对微生物组产生调节作用,使其与健康动物中观察到的情况相似,从而减少与疾病相关的肠道生态失调。
结肠炎生态失调得到改善
同样,在诱导结肠炎小鼠模型中,摄入布拉迪酵母菌(Saccharomyces boulardii)或两株发酵乳杆菌(Lactobacillus fermentum )已显示出能够调节肠道微生物组,增加生物多样性,并显著改善结肠炎动物的肠道生态失调特征。
结直肠癌的致病菌得到抑制,有益细菌增加
另外,在结直肠癌小鼠模型中,丁酸梭菌(Clostridium butyricum)的干预减弱了致病菌的增加,促进了有益菌的生长,并改变了次生胆盐和短链脂肪酸等微生物源性代谢产物。
在小鼠哮喘模型、抗生素诱导的生态失调小鼠和健康动物中也报道了这些调节作用,显示生物多样性和有益细菌丰度的增加。
益生菌对微生物群的调节在人类中并不明显,因为有的研究显示了相互矛盾的结果。可能还需一系列深入研究来了解益生菌对人体肠道微生物群的具体影响。
下面是存在争议的一些数据:
在最近的一项工作中,研究表明,以嗜酸乳杆菌(L. acidophilus)、乳酸乳杆菌(B. lactis)、长双歧杆菌(B. longum)、双歧双歧杆菌(B. bifidum)、和半乳糖低聚糖混合物组成的共生生物作为益生元进行干预,显示出在一组肥胖患者中调节肠道微生物群落的能力,观察到不同健康相关菌群(如双歧杆菌、毛螺菌和乳杆菌)的比例有所增加;以及与肥胖正相关的慢性炎症相关群体的减少,如瘤胃球菌科(Ruminococcaceae),普雷沃氏菌(Prevotella)和巨球型菌属(Megasphaera) 。
然而,先前对肥胖患者的研究显示了相互矛盾的结果,因此其中一些研究报告称,在益生菌干预后,肠道微生物群落没有变化,质疑这种对肠道微生物组的调节作用的存在。
与此同时,对影响全球数百万人的炎症性肠病患者的研究也显示出有争议的结果。尽管在动物模型中已经报道了对肠道微生物群落的调节作用,但在一组溃疡性结肠炎患者中,用含有短双歧杆菌(B. breve)的某品牌益生菌治疗,在研究过程中,两个治疗组之间没有显著差异。
另一方面,摄入含有L.acidophilus La-5和B. animalis亚种的益生菌酸奶。促进了一组炎症性肠病患者中乳酸菌、双歧杆菌和拟杆菌水平的增加,尽管这项工作没有从宏基因组的角度分析微生物组。
✦特定菌属水平增加,整体菌群组成变化不大
一些研究报道,益生菌不影响整体菌群组成,但可以改变特定属的比例。
例如,在高危特应性疾病婴儿中,在产前最后阶段和出生后第一年摄入益生菌混合物促进了双歧杆菌和乳酸杆菌水平的增加,但在肠道微生物组中没有检测到显著变化。
同样,在一组有功能性便秘症状的个体中,混合使用益生菌的干预促进了瘤胃球菌(Ruminococcus)水平的增加和丹毒丝菌科(Erysipelotrichaceae) 成员的减少,但α多样性指数和整体微生物组组成显示出与安慰剂组检测到的水平相似。
在一组HIV感染个体中,在使用鼠李糖酵母菌GG干预8周后,也描述了相同的趋势。
✦增加与健康相关共生细菌,减少病原体相关细菌
其他研究表明,益生菌可以引起肠道微生物种群的变化,增加与健康相关的共生细菌的水平,减少与病原体相关的群体,甚至恢复被改变的微生物群落的平衡。
在这方面,干酪乳杆菌(Lactobacillus casei Lcr35)在一组患有急性腹泻的儿童中,促进了拟杆菌、粪杆菌和瘤胃球菌属的增加,并降低了大肠杆菌和梭状芽孢杆菌的水平。
在一项针对乳糖不耐受患者的研究中,使用B.longum BB536和L.rhamnosus HN001进行30天的干预也导致肠道微生物组成发生积极变化。
在感染幽门螺旋杆菌的患者中也观察到了这种趋势。他们报告了多重耐抗生素的屎肠球菌LAB制剂与抗生素联合使用时,在正常肠道微生物群方面的有效效果,防止α多样性值的下降,并恢复受抗生素治疗影响的微生物群的水平。
同样,在一组患有早期败血症的成年人中,益生菌的干预导致α多样性的增加以及益生菌的增加,如不同种类的乳酸菌(Lactobacillus)和屎肠杆菌(E.faecium)。
此外,一些研究表明,益生菌不仅可以引起特定细菌种群的变化,还可以引起相关微生物群代谢物的变化。
✦肝硬化患者肠道失调得到改善
在肝硬化患者中,摄入8周鼠李糖乳杆菌GG不仅能调节肠道微生物组,降低肠杆菌科(Enterobacteriaceae)和紫单胞菌科(Porphyromonadaceae)的相对丰度,增加共生菌毛螺菌科(Lachnospiraceae)和瘤胃球菌科(Ruminococcaceae)的丰度,还能调节代谢物的分布,从而改善肠道生态失调和微生物-代谢组的联系。
在摄入增强结肠短链脂肪酸生成的发酵牛奶后,也有类似的趋势被报道;在乙型肝炎诱导的肝硬化患者中摄入含有丁酸梭菌(C.butyricum)和婴儿双歧杆菌(Bifidobacterium infantis)的益生菌混合物3个月后,促进了不同肠粘膜屏障完整性生物标志物的改善。
考虑到来自动物模型和人体试验的这一证据,最近有人提出将益生菌作为预防和治疗结直肠癌的新策略。
注:迄今为止,缺乏针对人类患者的研究,专注于分析益生菌对与CRC相关的肠道菌群紊乱的可能调节作用。
✦健康人群病原相关菌丰度下降
就健康人群而言,只有少数干预研究分析了益生菌摄入量对肠道微生物群的影响,而肠道微生物群可以根据参与者的年龄进行分类。
在一项对3至12个月大的婴儿的研究中,使用三种益生菌菌株进行干预(B.longum subsp. infantis R0033, Lactobacillus helveticus R0052和B. bifidum R0071)。8周后,一些病原相关类群如柯林斯菌属(Collinsella),肠球菌(Enterococcus)和克雷伯氏菌(Klebsiella),粪杆菌(Faecalibacterium)和瘤胃球菌(Ruminococcus)水平下降。
此外,在干预期间,一些细菌群保持稳定,而在安慰剂组中,它们增加了,如大肠杆菌志贺氏菌(Escherichia-shigella)和韦荣氏球菌属(Veillonella)的情况下
增加有益细菌,减少病原体相关菌的一些证据:
在另一项对健康婴儿的研究中,在生命的第一年摄入补充了4种双歧杆菌菌株的配方,促进了脆弱拟杆菌(Bacteroides fragilis)和经黏液真杆菌属(Blautia)水平的下降,以及代谢物谱的差异,尽管没有检测到整体微生物组的变化。
在成人人群中,有研究表明,益生菌摄入量对肠道微生物群落的影响与肠道菌群的基础组成密切相关。每天食用干酪乳杆菌(L.casei)14天,增加了某些有益细菌的相对丰度,如乳酸杆菌(Lactobacillus)、罗氏菌属(Roseburia)、粪球菌属(Coprococcus),而一些有害细菌的水平则降低了。
在用Lactobacillus kefiri LKF01治疗后,以及B.longum BB536和L.rhamnosus HN001进行干预后,也报告了类似的结果,显示出强大的调节肠道微生物群组成的能力,导致有益细菌的增加,并显著减少直接参与促炎反应和胃肠道疾病发作的潜在有害菌群。
✦产丁酸盐细菌增加
此外,每天食用两歧歧杆菌(B.bifidum strain Bb)4周会影响粪便微生物群中优势类群的相对丰度,并调节粪便丁酸盐水平,因此作者观察到普雷沃氏菌(Prevotellaceae)的数量减少,而瘤胃球菌(Ruminococcaceae)和Rikenellaceae的比例更高。
这些研究支持了之前的研究,即益生菌对肠道菌群的调节作用。有趣的是,最近有人提出了益生菌“ salami”的潜力,尽管微生物组的整体结构没有发现显著变化,但摄入益生菌萨拉米会促进产生丁酸盐的细菌的增加。
在益生菌摄入过程中检测到粘附相关基因的差异表达和大量参与细菌运动性的基因,提示益生菌干预可以通过调节其表达模式来调节肠道微生物组。
小结
总之,在过去的几十年里,大量的科学报告已经证实了益生菌对宿主健康的积极作用。然而,尽管存在不同的动物模型和人体研究,显示益生菌可能对肠道微生物群落及其功能具有调节作用,但在人体试验中报道的还存在一定的争议。
现有的研究使用了不同的益生菌菌株、广泛的年龄范围、疾病和疾病状态,以及摄入不同饮食和营养素的志愿者;因此,研究组表现出较高的个体内部和个体间的变异性,使得阅读结果变得困难。
另一方面,正如前面提到的,我们必须考虑到益生菌的作用不仅取决于菌株,还取决于基础微生物群,而最常用的益生菌(主要属于乳酸杆菌和双歧杆菌)不一定在肠道定植,并且可以在益生菌干预后释放,从而使确定益生菌调节宿主肠道微生物群的具体机制变得更加复杂。
为了阐明益生菌对肠道环境和微生物群影响的分子机制,我们应该进一步研究更大、更均匀的群体,也许还应该研究从人类肠道环境中分离出来的下一代益生菌,它们能够在肠道粘膜上定植,产生长期效应。
益生元概念首次定义为“不易消化的食物成分,通过选择性地刺激一种或少数已经存在于结肠中的细菌的生长或活性而有益地影响宿主”。
然而,这一概念多年来一直在发展。当前的国际益生菌和益生元协会共识小组提出了以下益生元的定义:“一种被宿主微生物选择性利用的对健康有益的底物”。
各种益生元
应该指出的是,目前确立的益生元是基于碳水化合物的,尽管其他物质如酚类化合物和多不饱和脂肪酸转化为各自的共轭脂肪酸可能符合这一定义。
几十年来,只有少数几类碳水化合物被认为是益生元,包括人乳寡糖(HMOs)、菊粉和低聚果糖(FOS)、低聚半乳糖(GOS)和乳果糖。
一些益生元及相关研究
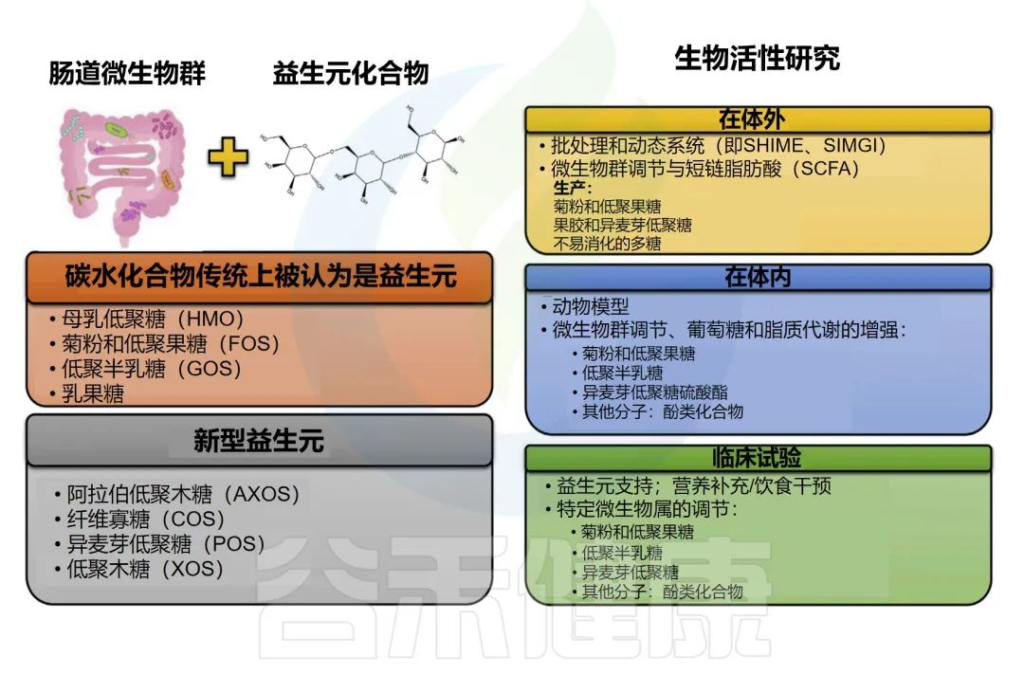
Nma B,et al.Elsevier Inc.2022
•人乳寡糖
人乳寡糖包括由5个单体(葡萄糖、半乳糖、n-乙酰葡萄糖胺、焦糖和唾液酸)通过多达12个不同的a-和b-糖苷键结合形成的广泛的低聚糖结构。
人乳寡糖对新生儿肠道菌群发育有很大影响,促进双歧杆菌(Bifidobacteriaceae)和拟杆菌(Bacteroidaceae)生长。
•低聚果糖
另一方面,低聚果糖是通过蔗糖的果糖转基化或菊粉的部分水解获得的,是唯一主要由果糖单位组成的碳水化合物。
•低聚半乳糖
相比之下,低聚半乳糖包括含有b键的b-GOS和来自乳糖延伸的b-GOS,以及存在于各种蔬菜来源中的a-GOS,如棉子糖和水苏糖。
•其他益生元
其他潜在的益生元低聚糖包括异麦芽糖-低聚糖(IMOS),由糖化植物淀粉中获得的由a键连接的葡萄糖单位形成的分支结构;
低聚木糖(XOS)和阿拉伯低聚糖(AXOS),由木聚糖和阿拉伯低聚木聚糖水解产生的含有阿拉伯糖酰和葡萄糖醛基分支的低聚木糖物组成;
纤维寡糖(COS),由木质纤维素生物质酶解获得,由b键连接的葡萄糖单位形成;
果胶寡糖(POS),由果胶部分水解产生,由半乳糖醛酸线性链形成,可能显示含有鼠李糖、阿拉伯糖、木糖和半乳糖等中性糖的分支结构域。
许多研究通过动物模型来评估体内益生元化合物的潜在生物活性。
✦增强代谢和生物利用度
通过16S rRNA基因测序和鸟枪法测序,大鼠给予低聚果糖和低聚半乳糖可导致普雷沃氏菌(Prevotella)水平升高,并由于普雷沃氏菌的代谢功能而增强了人参总苷的生物转化和生物利用度。
深度宏基因组测序实验已经报道了低聚果糖在高脂饮食喂养小鼠中促进了以下菌群的生长:
Allobaculum ↑↑↑
Oribacterium ↑↑↑
普雷沃氏菌(Prevotella) ↑↑↑
并降低了以下菌群:
嗜胆菌属(Bilophila) ↓↓↓
Butyrivibrio LE30 ↓↓↓
鸟枪测序分析还显示,菊粉处理的高脂饲料饲养的小鼠后代的葡萄糖和脂质代谢途径增强。
以下菌群水平增加:
B. breve ↑↑↑
B. acidifaciens ↑↑↑
Clostridium sp. CAG 343 ↑↑↑
Eubacterium sp. CAG 786 ↑↑↑
下列菌群水平下降:
B. massiliensis ↓↓↓
Oscillibacter sp. 1-3 ↓↓↓
Ruminococcus gnavus CAG 126 ↓↓↓
✦降低致病菌丰度
在肉鸡等其他动物模型中测试了低聚果糖补充剂作为抗生素的替代品,导致致病性脱硫弧菌属(Desulfovibrio)和幽门螺杆菌(Helicobacter)的丰度降低。
研究集中在其他寡糖家族,如异麦芽低聚糖,揭示了绿茶与异麦芽低聚糖联合使用,通过预防小鼠肠道生态失调来对抗高脂肪饮食诱导的代谢改变,显示了有益细菌(如Akkermansia muciniphilia,双歧杆菌,乳酸菌和罗氏菌属)丰度的增加。
✦非碳水化合物类益生元可以改善肥胖和肠道炎症
非碳水化合物的调节作用也在体内进行了研究,并通过元分类学评估了微生物种群的变化。已有研究证明,富含多酚的蔓越莓提取物可预防饮食诱导的肥胖、胰岛素抵抗和肠道炎症,并增加小鼠肠道微生物群中嗜黏蛋白阿克曼菌(Akkermansia muciniphila)丰度。
同样,槲皮素对高脂饮食诱导的非酒精性脂肪肝疾病的保护作用,以及类黄酮通过调节厚壁菌门/拟杆菌门比率来改善肥胖诱导的小鼠肠道生态失调的潜力已被描述。
临床试验
已经进行了几项临床试验,以测试不同益生元化合物的潜在作用。
✦增加新生儿双歧杆菌丰度
经荧光原位杂交测定,含有低聚半乳糖和低聚果糖混合物(比例为9:1)的配方可促进婴儿体内双歧杆菌的生长,而补充相同比例的短链低聚半乳糖和长链低聚果糖可导致丙型肝炎病毒感染母亲的新生儿粪便双歧杆菌和乳酸菌数量增加。
此外,肠内补充80%短链低聚半乳糖和长链低聚果糖(比例9:1)和20%酸性果胶寡糖的益生元混合物可导致早产儿双歧杆菌和出生后肠道定植增加。
✦抑制了艾滋病患者中一些有害菌的生长
在人类免疫缺陷病毒1型(HIV-1)感染的成人中,类似的益生元混合物促进了双歧杆菌的生长,并抑制了以下细菌的生长:
Clostridium coccoides ↓↓↓
直肠真杆菌(Eubacterium rectale) ↓↓↓
Clostridium lituseburense ↓↓↓
Clostridium histolyticum ↓↓↓
✦改善肥胖人群肠道菌群构成
同样,服用低聚半乳糖导致50岁以上男性和女性双歧杆菌丰度增加,并增加双歧杆菌数量,减少超重成人粪便样本中拟杆菌和Clostridium histolyticum的数量。
临床试验还报告了低聚果糖和果聚糖的益生元效应,结果表明菊粉型果聚糖导致双歧杆菌和普拉梭菌(F. prausnitzii)的增加;根据16S rRNA基因测序数据,这两种细菌都与血清脂多糖水平呈负相关,并且还降低了肥胖女性的肠道拟杆菌、Bacteroides vulgatus和丙酸杆菌(Propionibacterium)。
✦增加健康成年人体内双歧杆菌含量
此外,根据对健康成年人粪便微生物群的分析,菊粉给药增加了铁含量低的女性的粪便双歧杆菌,大豆寡糖、棉子糖和水苏糖可以被双歧杆菌选择性代谢。
✦其他益生元的作用
研究还评估其他益生元化合物的潜在效应。
食用聚葡萄糖可增加健康男性体内戴阿利斯特杆菌属 (Dialister)、普拉梭菌(Faecalibacterium)和考拉杆菌属(Phascolarctobacterium)的丰度。
这几类细菌的作用在谷禾之前的文章中有具体讲述
根据两项研究中获得的16S rRNA基因测序数据,B. animalis subsp. lactis 420促进了阿克曼菌、Christensenellaceae和Methanobrevibacter的生长,并降低了健康超重或肥胖个体中Paraprevotella的生长。
在健康个体中,部分水解瓜尔胶的饮食干预促进了拟杆菌、普拉梭菌、Fusicatenibacter和瘤胃球菌的生长,并减少了经黏液真杆菌属、毛螺菌科和罗氏菌属的生长。
值得注意的是,这些影响在男性参与者中更为明显。
同样,根据16S rRNA基因测序数据,女性不孕症患者口服部分水解瓜尔胶可导致双歧杆菌丰度增加,拟杆菌丰度降低,改善肠道生态失调和妊娠成功。
每日饮用红酒多酚可增加健康男性中的菌群数量,包括以下菌属:
拟杆菌属(Bacteroides) ↑↑↑
双歧杆菌属(Bifidobacterium) ↑↑↑
肠球菌属(Enterococcus) ↑↑↑
普雷沃氏菌属(Prevotella) ↑↑↑
Blautia coccoides ↑↑↑
Eggerthella lenta ↑↑↑
直肠真杆菌(E.rectale) ↑↑↑
益生菌定义中的关键词是“活的”,这意味着微生物必须是活的(在施用时)才能发挥有益的作用。
但是,在过去的几年里,越来越多的证据表明,不可活的细菌,它们的成分或它们分泌的代谢物,也可能在对健康的积极影响中发挥关键作用。
“后生元”一词于2011年首次提出,指的是摄入后对健康有益的灭活、不可存活的微生物细胞。
这一概念是基于观察到非活细菌或其提取物(破碎的细菌)具有调节宿主免疫反应的能力。
✦后生元相较益生菌的优势
将后生元与益生菌进行比较发现,后生元生产的复杂性较低,具有更好的长期稳定性。
对于某些高炎症状态的患者,使用后生元可能是更安全的替代方案,避免了使用活微生物时反应加剧。
在对术语进行分类的尝试中,提出了“true probiotics”(活的和活跃的细胞)、“pseudo-probiotic”(活的和不活跃的细胞,无论是营养细胞还是孢子细胞)和“ghost probiotics”(非活的或死亡的细胞,完整的或破裂的。
此外,活微生物对食物基质成分的活性也可能释放对健康有积极影响的副产物。例如,牛奶蛋白中加密的生物活性肽。
✦后生元的组成
后生元的组成可以是可变的。除了灭活的(无生命的)微生物生物量,细胞壁的成分(肽聚糖、蛋白质、糖蛋白、磷壁酸)、细胞质膜(磷脂、蛋白质)或细胞外成分,如胞外聚合物(EPS)。此外,分泌到培养基上清液或特定发酵食品的代谢物也可能存在。
后生元的成分示例及显微镜下图像
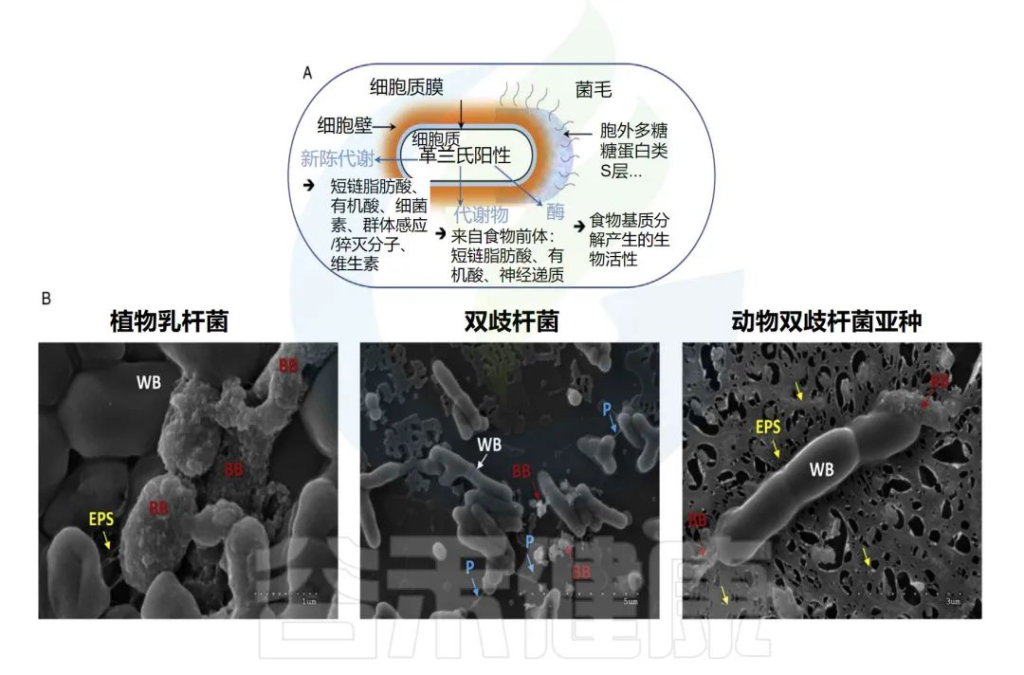
Nma B,et al.Elsevier Inc.2022
✦后生元的作用
后生元作用机制与益生菌的类似,即调节有益菌群的能力、对病原体的拮抗作用、增强肠上皮屏障功能的能力以及调节免疫反应或宿主的其他生理功能。
•刺激免疫反应
双歧杆菌产生的EPS,作为报道的对产生细菌有益作用的关键成分之一。无论是被聚合物包围的紫外线灭活双歧杆菌还是纯化的EPS本身,都能够以不同的方式刺激免疫反应,这取决于聚合物的物理化学特性。这些细菌EPS能够与肠上皮细胞的TLR4相互作用。
•促进脂质代谢和葡萄糖稳定
此外,双歧杆菌EPS是能够积极调节菌群组成的碳水化合物,因为它们被用作肠道某些细菌的选择性发酵底物。特异性EPS还促进饮食诱导的肥胖小鼠模型的脂质代谢和葡萄糖稳态的变化。
含有能够释放抗菌化合物的活益生菌,也可以被视为后生物制剂,可能有助于提高发酵产品的安全性。
将后生元应用于食品,虽然是改善我们健康的一种新方法,但仍然具有挑战性。
精神生物制剂
关于肠道菌群在维持我们生理系统(被称为微生物-肠道轴)方面的知识越来越多,这促使了与调节这种双向对话的饮食策略相关的新概念的发展。这些轴之一是研究的热点,即微生物-肠道-大脑轴。
这是基于对肠道微生物群活动可能参与神经疾病和随着年龄增长而维持认知功能。
▸ 精神生物制剂
针对这目标,提出了与益生菌相关的新定义:精神生物制剂是一种“活的有机体,当摄入足量时,会对患有精神疾病的患者产生健康益处”。
最近,这一定义扩大到包括能够调节肠-脑轴影响肠道微生物群的底物;因此,精神生物制剂是“有益的细菌(益生菌)或对影响细菌-大脑关系的细菌(益生元)的支持”。
研究强调,某些微生物代谢物可以作为某些代谢紊乱的调节因子。同样,在肠道-大脑相互作用的机制中,微生物代谢物也是关键角色。
✦改变行为
已经证明一些微生物来源的精神生物制剂能够有效地改变行为。在一种优雅的实验中,产生酪胺的Providencia能够以一种对共生和宿主都有益的方式操纵秀丽隐杆线虫(Caenorhabditis elegans)的“感官”决定;在食物选择测定中,被Providencia定植的线虫选择了含有这种细菌的食物。
然而,肠道微生物群代谢组的操纵非常复杂,因为肠道微生物的大量多样性之间存在着一种微生物代谢物生产者和消费者之间持续反馈的串扰。
例如,γ-氨基丁酸GABA——(中枢神经系统的主要抑制递质)可以由肠道微生物从饮食前体(如谷氨酸)合成,但它被用作支持该生态位中其他生物生长的营养素。
✦减轻疼痛
神经调节的精神生物学已经被提出,通过体外实验和在大鼠模型上的进一步验证,表明嗜酸乳杆菌菌株(Lactobacillusacidophilus NCFM)能够在肠道水平诱导鸦片样物质和大麻素受体的合成,从而作为一种镇痛药减轻疼痛。
✦调节焦虑
对无菌小鼠口服乳酸菌(Lactiplantibacillus plantarum PS128) 可增加小鼠大脑特定区域的多巴胺和血清素水平,并调节动物的焦虑样行为。
关于能够产生γ-氨基丁酸(中枢神经系统的主要抑制递质)的益生菌,已经在一些乳酸菌和双歧杆菌中描述了不同的代谢途径,其中Levilactobacillus是迄今为止量化产量最高的物种。
不同的策略,如优化培养基组成,可用于提高其他物种的γ-氨基丁酸产量。此外,其中一些乳酸菌也被用于发酵乳制品,以改善这种神经递质在食物中的合成。
//建议
通过γ-氨基丁酸前体的生物转化将食物与γ-氨基丁酸进行天然生物强化,以及使用食物或补充剂作为在肠道中输送能够合成γ-氨基丁酸的益生菌的载体,可能是调节大脑活动的一种策略。
上述的一些证据已经证明,发酵食品、健康的饮食模式、益生菌、益生元以及后生元等都是促进肠道健康的重要因素。通过食用这些食物和补充这些营养素,可以增加肠道内有益菌的数量和种类,并在一定程度上抑制病原相关菌群的丰度,维持肠道菌群的平衡,从而促进肠道健康。
饮食在塑造肠道微生物群的组成和活性方面起着重要作用,对营养饮食与健康肠道微生物组之间相互作用的深刻理解,将为我们理解其在疾病预防和治疗中的作用奠定基础。
任何饮食的好处都将在很大程度上取决于此人的微生物组,每个人根据自己独特的肠道微生物群摄入不同的饮食的方式,将会使我们迎来新时代的饮食模式。
除了肠道健康之外,针对多个健康维度例如提升免疫力,改善情绪,体重管理,皮肤管理等领域产品和食品的需求都在日益提升,这些与肠道微生物组之间又都存在密不可分的关联。因此基于肠道微生物组重要性的开创性研究,可能会从根本上改变消费者偏好及健康食品行业的走向。
主要参考文献
Nma B , Csa C , Ica C , et al. Mechanisms of Gut Microbiota Modulation by Food, Probiotics, Prebiotics and More – ScienceDirect.
Agus, A., Clément, K., Sokol, H., 2021. Gut microbiota-derived metabolites as central regulators in metabolic disorders.
Ali, A., Kamal, M., Rahman, H., Siddiqui, N., Haque, A., Saha, K.K., Rahman, A., 2021. Functional dairy products as a source of bioactive peptides and probiotics: current trends and future prospectives. J. Food Sci. Technol.
Barros, C.P., et al., 2020. Paraprobiotics and postbiotics: concepts and potential applications in dairy products. Curr. Opin. Food Sci. 32, 1–8.
Bellini, M., Tonarelli, S., Nagy, A.G., Pancetti, A., Costa, F., Ricchiuti, A., de Bortoli, N., Mosca, M., Marchi, S., Rossi, A., 2020. Low FODMAP diet: evidence, doubts, and hopes.Nutrients 12 (1), 148.
Cunningham, M., Acarate-Peril, M.A., Barnard, A., et al., 2021. Shaping the future of probiotics and prebiotics. Trends Microbiol.
Fiore, W., Arioli, S., Guglielmetti, S., 2020. The neglected microbial components of commercial probiotic formulations. Microorganisms 8, 1177.