 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室
二级病原微生物安全实验室- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室


短链脂肪酸这一词经常出现在谷禾的文章和报告中,那你真的了解短链脂肪酸吗?短链脂肪酸(SCFA)主要是肠道微生物群在结肠内通过发酵碳水化合物(包括膳食和内源性碳水化合物,主要是抗性淀粉和膳食纤维)和一些微生物可利用的蛋白质而产生的。
短链脂肪酸主要是乙酸、丙酸和丁酸,在结肠中的浓度比大致为60-70%:20-30%:10-20%。这些代谢产物能够被宿主利用,尤其是丙酸和丁酸,它们发挥一系列促进健康的功能。它们能被肠粘膜有效吸收,作为能量来源,还能作为基因表达调节剂以及特定受体识别的信号分子,对宿主生理产生重要影响。近几年的研究还发现其能够调节免疫细胞发育并抑制炎症。
然而,三种主要短链脂肪酸——乙酸盐、丙酸盐和丁酸盐——它们在体内的功能和组织分布不同,对宿主生理的影响存在显著差异。丁酸盐优先被肠粘膜用作能量来源,还具有抗炎特性,可增强肠道屏障功能和粘膜免疫。丙酸盐有助于抵抗肝脏中的脂肪形成、降低胆固醇、也有一定的抗炎和抗致癌作用。而乙酸在血液中浓度最高,可用作肝脏胆固醇和脂肪酸合成的底物,增加结肠血流量和氧气吸收,并通过影响回肠收缩来增强回肠运动。
其次,它们与宿主蛋白(如丁酸和丙酸抑制组蛋白去乙酰化酶)和受体的相互作用也不同。因此,特别需要考虑这些短链脂肪酸的微生物来源。与此同时,饮食结构和肠道生理变化如何影响这些脂肪酸的相对产量和在结肠中的浓度也是至关重要的。
本文将重点介绍丙酸盐和丁酸盐,因为这两种短链脂肪酸被认为对健康有重要影响,包括丁酸预防结直肠癌,丙酸促进饱腹感并降低胆固醇。而乙酸是大多数肠道厌氧菌的主要发酵产物,也可由还原性乙酸生成,而丙酸和丁酸则由不同的肠道细菌群产生。
我们将探讨在人类结肠微生物群中已知的形成这两种短链脂肪酸的途径,以及各种饮食和环境因素对其产生的调节可能性。详细了解肠道微生物群的短链脂肪酸代谢及其生理功能对于制定个性化的健康营养方案是必不可少的。
丙酸盐和丁酸盐在结构、来源、生理功能以及对健康的影响方面存在一些差异。
▸ 结构上的差异:
丙酸盐(Propionate)含有三个碳原子,羟基(-OH)位于第二个碳原子上。
丁酸盐(Butyrate)含有四个碳原子,羟基(-OH)位于第四个碳原子上。
▸ 来源上的差异:
丙酸盐通常由肠道细菌通过发酵L-鼠李糖、聚葡萄糖、阿拉伯木聚糖、D-塔格糖、甘露寡糖、昆布多糖等糖类物质产生。
丁酸盐可以通过肠道微生物群发酵富含抗性淀粉和果聚糖的食物来增加,如菊粉、马铃薯、洋葱等。
▸ 功能上的差异:
丙酸盐在体内的主要功能是作为肝脏中糖原合成的前体物质,有助于调节血糖水平,影响食欲。
丁酸盐是结肠上皮细胞的首选能源,有助于维持肠道屏障,发挥免疫调节和抗炎作用。丁酸盐还可以通过减少NF-kB信号传导和诱导凋亡来促进神经保护。
▸ 对健康的影响:
丙酸盐的健康益处主要是调节能量代谢和改善代谢综合征,可能对调节肠道菌群平衡也有积极作用,但其具体影响取决于个体的肠道菌群组成。
丁酸盐对健康的影响更为广泛,包括维持肠道稳态、促进肠道屏障的完整性、刺激绒毛的生长、促进粘蛋白的产生。以及改善认知功能、促进睡眠、调节社交行为和在糖尿病中的潜在益处。
丙酸盐是许多生物(从细菌到人类)的代谢副产物,产生丙酸的代谢途径可分为三类。主要发酵途径将不同的碳源分解代谢为丙酸盐(图A);分解代谢途径则能将多种氨基酸降解为丙酸盐(图B);此外,通过与从丙酮酸或二氧化碳生成生物质前体相关的合成代谢途径也可以生产丙酸盐(图C)。
产生丙酸盐的代谢途径
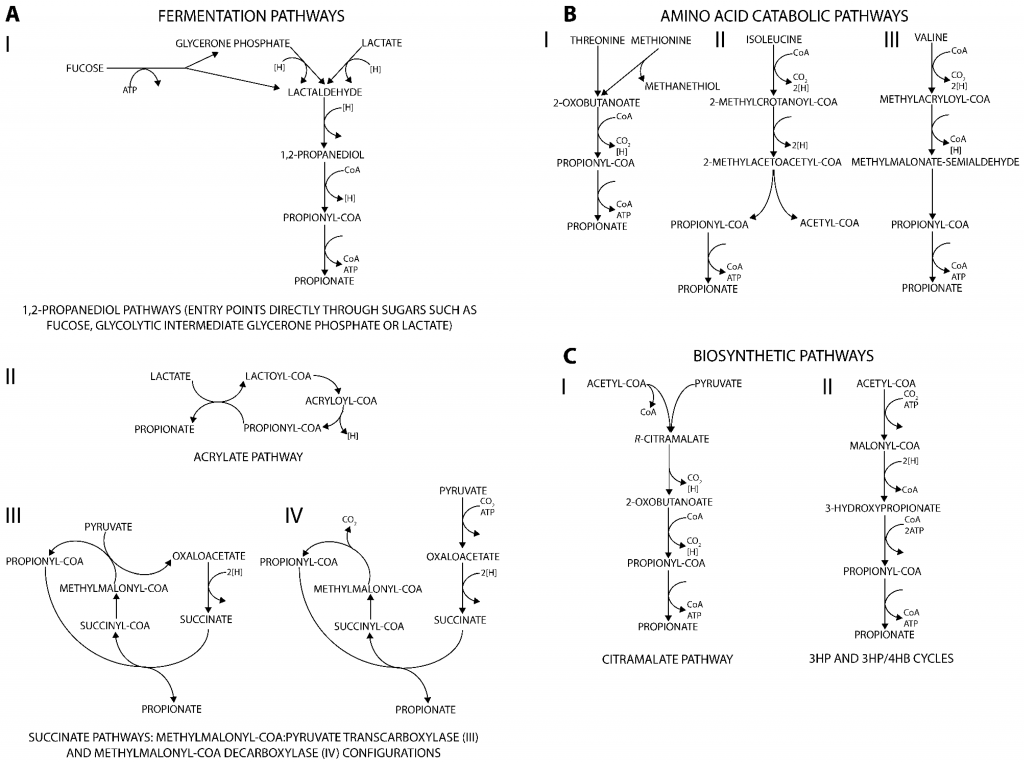
doi.org/10.3390/fermentation3020021
肠道微生物通过发酵不可消化的碳水化合物产生的丙酸盐是体内丙酸盐的主要来源。下面将讲述肠道中丙酸盐通过发酵碳水化合物的三种主要产生途径以及参与这些途径的微生物。
与氨基酸降解和生物合成途径相比,发酵途径不仅提供能量,还帮助消耗由糖分解代谢产生的还原辅因子。它们在能量产生和维持氧化还原平衡中的作用,使这些途径能够与细胞生长相耦合。
能够产生丙酸盐的微生物种类

doi.org/10.3390/fermentation3020021
▸
丙二醇相关途径
• 鼠伤寒沙门氏菌和Roseburia inulinivorans在这一过程可以产生丙酸
丙酸盐在肠胃中的生成是由1,2-丙二醇(PDO)发酵菌和PDO消耗菌组成的微生物联合作用的结果。已知一些生物体(如鼠伤寒沙门氏菌和Roseburia inulinivorans)能同时进行这两种过程。
在二醇脱水酶和两种常与乙酸代谢相关的混杂酶(辅酶A依赖性醛脱氢酶磷酸转酰基酶和乙酸激酶)的共同作用下,PDO分解代谢为丙酸,同时生成一个ATP和一个还原辅因子。
然而,通过该途径产生丙酸取决于可用于其他细菌生长的碳水化合物,据报道岩藻糖和鼠李糖是丙酸的前体。
通过1,2-丙二醇形成丙酸的微生物途径
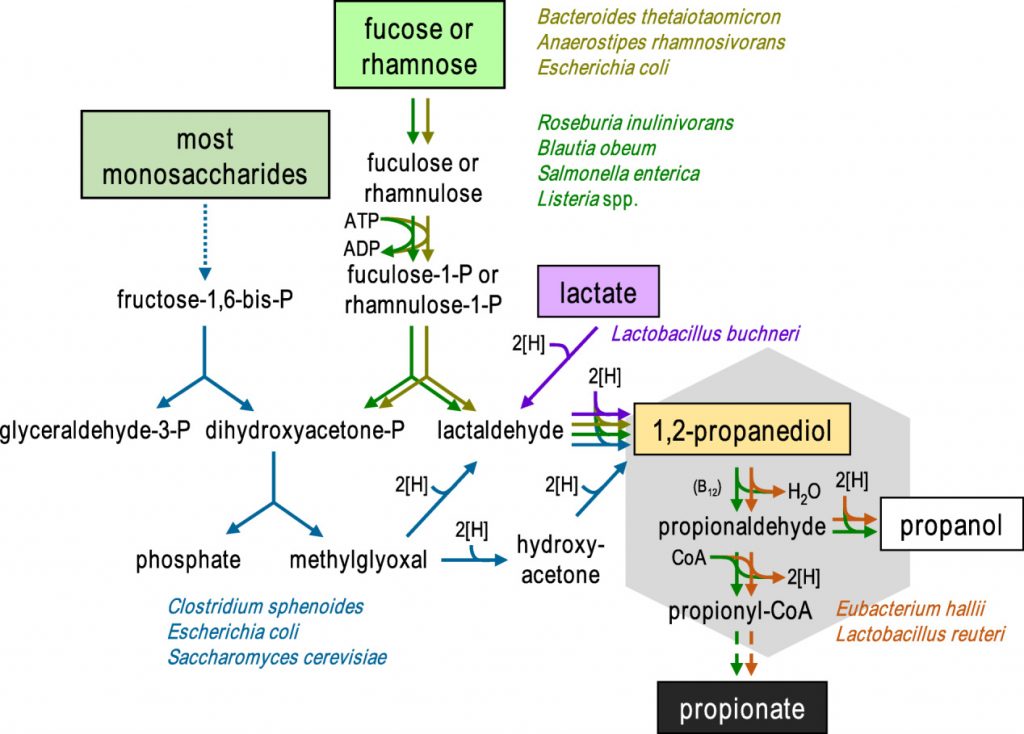
doi: 10.1111/1462-2920.13589.
霍氏大肠杆菌(E.hallii)和罗伊氏乳杆菌(Lactobacillus reuteri)虽然无法在岩藻糖或鼠李糖上生长,但仍然能够利用1,2-丙二醇产生丙酸和丙醇。此外,宏基因组研究表明,其他肠道厌氧菌,包括Flavonifractor plautii、Intestinimonas butyriproducens和Veillonella spp. 也可能能够从这种底物中产生丙酸。因此,不同细菌之间中间体 1,2-丙二醇可能在脱氧糖生产丙酸中发挥重要作用。
▸ 丙烯酸酯途径
丙烯酸酯途径在消耗NADH的情况下,使乳酸在ATP中性条件下转化为丙酸。该途径存在于几种细菌中,包括丙酸梭菌(Clostridium propionicum)、埃氏巨球菌(Megasphaera elsdenii)和瘤胃普氏菌(Prevotella ruminicola)。
虽然多种底物可以分解为丙酸和乙酸,包括乳酸、丝氨酸、丙氨酸和乙醇,但葡萄糖发酵在这过程中似乎不会导致任何天然生产者产生丙酸,这可能是因为葡萄糖发酵不会触发启动循环所需的乳酸消旋酶的表达。
▸ 琥珀酸途径
琥珀酸途径主要存在于拟杆菌门和厚壁菌门中,拟杆菌门的一些细菌从膳食碳水化合物生成丙酸,并且拟杆菌门的相对丰度与人类粪便中丙酸盐的相对水平相关。
• 琥珀酸转化为丙酸盐还需要维生素B12
琥珀酸是丙酸的前体,但在高pCO2(二氧化碳分压)和高稀释率等条件下,它可在磷酸烯醇式丙酮酸羧激酶受到抑制的拟杆菌属培养物中积累。琥珀酸转化为丙酸还需要维生素B12,如果缺乏B12,琥珀酸可能无法转化成丙酸盐。
人类结肠中的一些厚壁菌门细菌(例如Phascolarctobacterium succinatutens)能将琥珀酸转化为丙酸;其他革兰氏阴性菌通过琥珀酸途径(如韦荣氏球菌属)或丙烯酸酯途径(Megasphaera elsdenii)将乳酸转化为丙酸盐。
▸ 氨基酸降解产生丙酸
缬氨酸、苏氨酸、异亮氨酸和蛋氨酸的降解可导致通过丙酰辅酶A产生丙酸和ATP。
由于氨基酸的合成和随后的分解代谢途径存在于多种微生物中,因此可以使用氨基酸合成代谢和分解代谢途径的组合从葡萄糖生产丙酸。
几种拟杆菌在蛋白水解和肽形成丙酸方面发挥重要作用。
苏氨酸的产生和分解代谢已在大肠杆菌中得到广泛研究,作为丙酰辅酶A的可能来源,可用于生产各种化学物质,包括丙醇、红霉素和3-羟基戊酸。此外,考虑到氧化还原和能量平衡,缬氨酸/异亮氨酸和苏氨酸途径的组合具有协同作用,可以略微提高丙酸产量。
除此之外,可以利用丙酮酸经过柠檬酸途径产生丙酸,由于和菌群关联较小,在这就不展开描述了。
▸ 增加丙酸盐生成的底物
一些益生元等化合物可以影响丙酸盐的生产,不过由于浓度以及肠道微生物群落构成的不同,这些化合物的调节丙酸盐产生时具有一定差异。
• L-鼠李糖能够明显增加丙酸盐产量
L-鼠李糖或6-脱氧-L-甘露糖是一种天然脱氧糖。它存在于多种动物、植物和细菌多糖中。在短期体外实验中,L-鼠李糖已被证明能使丙酸产量增加四倍于乳果糖。
在一项人体体内研究中也获得了类似的结果,其中受试者在三个不同的时间被给予25克L-鼠李糖、乳果糖或D-葡萄糖。摄入后24小时测量血清丙酸,L-鼠李糖的血清丙酸明显高于乳果糖或D-葡萄糖的血清丙酸。
一项长期研究也证实了L-鼠李糖诱导丙酸的作用,该研究结果表明,与摄入D-葡萄糖作为对照相比,摄入25克L-鼠李糖可显著提高人体28天内的血清丙酸水平。
• 抗性淀粉有助于增加丙酸
抗性淀粉对淀粉酶降解具有抗性,但它会发酵成丁酸或丙酸。特别是,来自大米的抗性淀粉与丙酸产量增加有关。
研究了抗性淀粉在大鼠体内的发酵情况,饲喂抗性淀粉(630g/kg饲料)的大鼠的肝脏甘油三酯和总胆固醇浓度显著低于对照组。与此同时,血清丙酸浓度也显著增加。
• 菊粉对于增加丙酸和丁酸非常有效
菊粉属于果聚糖家族,主要由β-(2,1)连接的果糖基组成。它天然存在于菊苣和菊芋等开花植物中。作为益生元,菊粉已被证明对增加丁酸和丙酸的产量非常有效。使用人体肠道微生物体外模拟研究了菊粉增加丙酸的效果。
补充菊粉1周(5g/d)后观察到短链脂肪酸产生的代谢变化。较高浓度的短链脂肪酸源于丙酸和丁酸产量的增加。
此外一项针对喂食菊粉(10%)的大鼠的体内研究也导致丙酸产量大幅增加,高达 58.4mmol/。
• 聚葡萄糖
聚葡萄糖是一种支链、随机聚合的多糖,主要由葡萄糖合成,在胃肠道上部不会被消化。使用结肠模拟器研究了这种底物对结肠微生物组成和代谢活性的调节作用。与对照糖木糖醇(8.3mmol/L)相比,短链脂肪酸产量显著增加,尤其是丙酸盐(22.9mmol/L)。
• 阿拉伯木聚糖
阿拉伯木聚糖是许多谷物中发现的主要非淀粉多糖,是膳食纤维的一部分。
在体内研究中,比较了54只大鼠,这些大鼠分别喂食对照饮食(含710g/kg小麦)、阿拉伯木聚糖补充饮食(610g/kg小麦淀粉加100g/kg玉米阿拉伯木聚糖)和胆固醇补充饮食(不含或含2g/kg 胆固醇)。由于短链脂肪酸的积累,尤其是丙酸(摩尔百分比>45%),盲肠pH值从7降至6。然而,丁酸的产生不受影响。
车前草是一种可溶性纤维来源,可提供与麦麸阿拉伯木聚糖相当的多糖。在一项大鼠体内研究中,比较了车前草(5%)对盲肠和结肠发酵的影响与麦麸 (10%) 的影响。研究发现,车前草发酵可产生更高的短链脂肪酸,尤其是盲肠和所有结肠中的丙酸更多。
除此之外,D-塔格糖、甘露寡糖、昆布多糖等物质也可以增加人体丙酸盐的产生。
丙酸盐已被证明具有抗脂肪形成和降低胆固醇的作用。它还对体重控制和进食行为有很强的影响。此外,有研究表明,丙酸和丁酸一样,对结肠癌细胞具有抗增殖作用。
丙酸盐对健康的影响
doi: 10.1111/j.1753-4887.
然而,与结肠细胞用作能量来源的丁酸不同,丙酸在血液循环中的浓度较高。因此,丙酸的生物活性可能不仅限于结肠本身,还扩展到人体的其他部位。需要强调的是,了解体内结肠丙酸浓度或短链脂肪酸浓度不足以推断健康状况。
▸ 影响肝细胞的脂质合成
肝脏的脂质合成包括将饮食来源的脂肪酸和甘油转化为具有不同脂肪酸组成的胆固醇和甘油三酯。然后,这些肝脏脂质分子被结合到脂蛋白中,从而通过循环分布到各种组织中。
• 脂质合成受到短链脂肪酸的强烈影响
有趣的是,肝细胞中的脂质合成受到肠道纤维发酵产生的短链脂肪酸的数量和类型的强烈影响。丙酸盐已被确定在其中一些过程中发挥重要作用。
对肝脏脂质合成的饮食调节早期观察显示,膳食纤维摄入与肝脏脂质合成密切相关。这种影响的部分原因如下:1)粪便中胆固醇和胆汁酸从肠道排出的增加;2)胆固醇向胆汁酸的肝脏转化率较高;3)通过减少乳糜微粒的大小和降低胆固醇在乳糜微粒中的掺入来优化脂蛋白的外周代谢。
• 降低血清胆固醇水平、抑制脂肪酸合成
短链脂肪酸作为微生物碳水化合物发酵的产物,在肝脏脂质合成中发挥着特定作用,已被证明在喂食大鼠时可降低血清胆固醇水平。对分离的大鼠肝细胞的体外研究表明,丙酸对脂肪酸合成有抑制作用,但对胆固醇合成没有抑制作用。
其他大鼠实验表明,加入纤维的饮食可降低肝和血浆胆固醇水平及血浆甘油三酯中的胆固醇,而肝甘油三酯没有受到影响。
尽管这些研究的结果令人信服,但其他研究并不总是能够证实丙酸盐对脂质代谢的抑制作用。例如,每天在面包中补充9.9克丙酸盐不会改变6名健康志愿者的脂质代谢,甚至会导致5名受试者的甘油三酯浓度升高。
在另一项研究中,比较了丙酸盐对人和大鼠肝细胞脂质代谢的影响。发现浓度为0.1mmol/L的丙酸盐对大鼠乙酸盐合成脂质有抑制作用。然而,在人类肝细胞中,需要更高浓度的丙酸盐(约10-20 mmol/L)才能获得同样的抑制作用。该值比门静脉血中丙酸盐的浓度高100-200倍,表明大鼠模型不能完全外推到人类的情况。
▸ 作为影响饱腹感的分子
丙酸盐不仅具有降低胆固醇和抗脂肪生成的作用,还可能通过刺激饱腹感来控制体重。已有研究表明,短链脂肪酸(如乙酸、丙酸、丁酸)具有诱发饱腹感的作用。
• 丙酸盐影响肠道激素的形成进而影响饱腹感
现有证据表明,细菌调节肠道激素(如胰高血糖素样肽1(GLP-1)和肽YY(PYY))的形成,是通过短链脂肪酸介导的。乙酸、丙酸和丁酸的生理浓度,以及pH值从7.5降至6.0,会显著增加肠内分泌结肠细胞系STC-1中的胰高血糖素原和PYY。
GLP-1和PYY是刺激饱腹感的激素,由L细胞(主要位于回肠和结肠)响应营养摄入而释放。GLP-1促进胰岛素分泌和胰腺β细胞增殖,同时控制肌肉细胞中的糖原合成;而PYY则减缓胃排空。相反,生长素释放肽刺激食欲,主要由胃中的P/D1细胞产生。
不易消化的碳水化合物,如低聚果糖、乳糖醇和抗性淀粉,通过调节肠道肽GLP-1、PYY和生长素释放肽的产生,有效地诱导饱腹感,这一机制还涉及肠道微生物群落。
• 丙酸盐对摄食行为有显著影响,诱导瘦素产生
在短链脂肪酸中,丙酸盐被重点研究作为一种饱腹感诱导剂,对能量摄入和摄食行为有显著影响。人体和动物试验表明,丙酸盐给药(体内范围为130-930mmol/L,体外范围为0.01-10mmol/L)显著增强饱腹感并降低进食欲望。
丙酸盐触发的饱腹感信号之一是瘦素,这是一种强效的厌食激素,通过中枢神经系统中表达的受体抑制食物摄入。研究显示,每天服用500µmol的丙酸盐几乎使小鼠血浆中的瘦素浓度翻了一倍。
在另一项研究中,浓度为3mmol/L的丙酸在mRNA和蛋白质水平上诱导了人内脏脂肪组织中瘦素的产生。这些数据表明,肠道微生物群对肥胖的调节作用可能部分由短链脂肪酸(特别是来自微生物碳水化合物发酵的丙酸)介导。
▸ 影响心脑血管健康
丙酸盐通过与肠道受体GPR 41和GPR 43(也称为脂肪酸受体FFAR2和FFAR3)相互作用,对心脑血管健康具有一些潜在的影响。
• 脑血管患者体内的丙酸盐含量较低
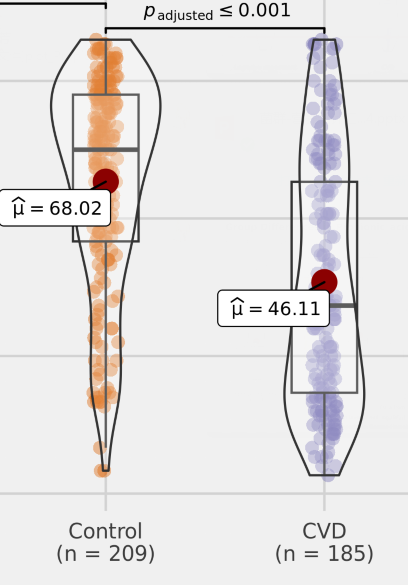
编辑
谷禾的数据中发现,与健康对照相比,脑血管疾病患者中的丙酸盐含量较低,并且具有明显的统计学差异。因此丙酸盐的含量可能是影响心脑血管健康的一个重要指标。
• 较高的丙酸盐浓度患冠状动脉硬化风险较低
最近一项大型横断面研究表明血浆丙酸浓度与冠状动脉疾病存在关联,较高浓度的丙酸盐与较低的冠状动脉粥样硬(CAD)风险相关,且与已知的心血管风险因素无关。
如上所述,越来越多的实验数据表明,丙酸盐可能对高血压、内皮功能障碍和高胆固醇血症等心血管风险因素产生有益影响。
▸ 丙酸盐在癌症中的潜在作用
短链脂肪酸对癌症(尤其是结肠癌)的影响已被广泛研究。丁酸能够调节基因表达,并对细胞凋亡和细胞周期的关键调节因子产生影响。
几种机制促成了丁酸对基因表达的调节作用。这些机制包括组蛋白和非组蛋白的过度乙酰化以及DNA甲基化的改变,从而增强了转录因子对核小体DNA的可及性。
• 丙酸盐诱导结直肠癌细胞凋亡
在一项研究中,丙酸盐和乙酸盐(浓度分别为26-40和9-16mmol/L)在人类结直肠癌细胞系中诱导了典型的细胞凋亡迹象。这些迹象包括线粒体跨膜电位的丧失、活性氧的产生、胱天蛋白酶3加工和核染色质凝聚。
• 抑制结肠癌细胞系的生长
短链脂肪酸对结肠上皮细胞增殖具有矛盾作用。虽然这些阴离子刺激正常隐窝细胞,但它们抑制结肠癌细胞系的生长。
丁酸盐和丙酸盐也是诱导分化和细胞凋亡的最有效脂肪酸。因此,它们通常可以预防癌症的发展,尤其是预防结直肠癌。虽然丁酸比丙酸更有效,但它主要被结肠细胞吸收作为能量来源。相比之下,丙酸和乙酸盐进入血液循环的浓度比丁酸高得多,并且它们被肝脏大量吸收(约60%)。由于这些阴离子在肝脏中的浓度很高,它们很可能会影响肝癌细胞以及已知会导致肝脏转移的其他典型癌细胞,例如乳腺癌和结肠癌。
一项针对猝死患者的研究表明,外周血中的短链脂肪酸数量可以量化。因此,这种循环中的丙酸盐、乙酸盐和丁酸盐的抗癌作用非常值得研究;例如,这种影响会在多大程度上延伸到小肠、大肠和肝脏之外,从而影响不同的组织?
!
• 丙酸盐过量可能与阿尔茨海默病相关
最近的几项研究提供了丙酸盐与阿尔茨海默病(AD)之间联系的证据。例如,分析了阿尔茨海默病患者和健康对照者的唾液样本,发现阿尔茨海默病患者的丙酸水平比健康对照者高出1.35倍。分析了轻度认知障碍者、阿尔茨海默病患者和健康对照者的唾液样本,也发现AD患者的丙酸水平显著升高。
几项啮齿动物研究也将粪便和循环中丙酸盐水平与阿尔茨海默病联系起来。接受AD患者粪便微生物群移植的小鼠丙酸水平高于对照组。
此外,发现阿尔茨海默病小鼠海马体中的丙酸增加了1.23倍。也发现阿尔茨海默病小鼠前额叶皮层中的丙酸浓度显著高于野生型小鼠,6个月大的阿尔茨海默病小鼠粪便中的丙酸浓度也显著更高。
• 丙酸血症的丙酸盐代谢异常
丙酸血症是人类最常见的有机酸代谢紊乱,是一种由丙酰辅酶A羧化酶基因缺陷引起的先天性代谢错误,丙酸过量且无法通过丙酰辅酶A转化。
丙酸血症患者的体内丙酸及其代谢物水平显著升高,会引起代谢性酸中毒、血氨升高,可能还会诱导一些严重的并发症如脑损伤、心肌病发生。
•有研究认为过多的丙酸盐可能导致肥胖和糖尿病
2019年哈佛大学公共卫生学院发表的一篇文献称:小鼠长期接触丙酸将导致肝细胞产生更多葡萄糖,形成高血糖症,这是糖尿病的一个典型特征。此外,研究人员还发现,长期服用丙酸盐剂量的小鼠,其体重会显著增加,胰岛素抗性也会增大。
还对14名健康人开展对照试验,受试者被随机分为两组,其中一组的餐食中添加1克丙酸盐,另一组添加安慰剂。结果显示,进餐后不久,丙酸盐组受试者血液中的去甲肾上腺素水平显著升高,胰高血糖素等激素水平也有所增长。这表明丙酸盐可能会作为“代谢破坏者”,增加人类患糖尿病和肥胖症的风险。
而这与前文丙酸盐会影响脂质合成与摄食行为相矛盾,因此,丙酸盐对人体的影响可能取决于其含量以及人体独特的肠道菌群结构与代谢能力。
内源性丙酸盐与外源性丙酸盐存在区别
内源性丙酸:指的是人体内部产生的丙酸,通常情况下,人体中的微生物能够在结肠中通过发酵未完全消化的碳水化合物来产生丙酸,这些丙酸对人体是有益的。
外源性丙酸:指的是从外部来源摄入的丙酸,比如通过食物摄入或作为补充剂。以及在一些药物中也可能含有丙酸盐。
★ 外源性丙酸盐摄入过多对代谢健康有害
但已经有各种研究表明,外源性地摄入和自体产生的丙酸作用并不一样,外来的丙酸对个体的许多代谢过程有不利影响。
哈佛大学公共卫生学院的研究人员完成了一项实验,发现从食品中摄入的丙酸在短期内会导致高血糖、胰岛素偏高,而且长期摄入将会造成肥胖和胰岛素抵抗等更严重的症状。
我们通过口腔摄入含丙酸添加剂的食物后,会激活我们的交感神经系统,并促使体内胰高血糖素和脂肪酸结合蛋白4等激素的升高。受到激素影响,肝糖原持续分解,血糖升高。身体开始分泌更多胰岛素进行代偿降低血糖浓度,最终导致胰岛素抵抗和肥胖产生。
这项研究表明,自身产生的丙酸和外界摄入的丙酸具有不同的功能,我们推测,这是因为从食物中摄入会极大地增加细胞和丙酸的接触,而细菌产生的仅仅只会局限在结肠部分。
在人体肠道中,丁酸由膳食纤维经细菌发酵产生,通过两种代谢途径。在第一条途径中,丁酰辅酶A被磷酸化形成丁酰磷酸,并通过丁酸激酶转化为丁酸。在第二条途径中,丁酰辅酶A的辅酶A部分通过丁酰辅酶A:乙酸辅酶A转移酶转移到乙酸盐,从而形成丁酸和乙酰辅酶A。其中丁酰辅酶A:乙酸辅酶A转移酶途径占主导地位。
对人类粪便微生物群的放射性同位素分析也表明,肠道中的大多数丁酸盐是由碳水化合物通过Embden-Meyerhof-Parnas(糖酵解途径)由乙酰辅酶A产生的。
• 少量的丁酸盐也可由蛋白质合成
除了碳水化合物,少量的丁酸也可以通过谷氨酸、赖氨酸、戊二酸和γ-氨基丁酸途径由蛋白质合成。厚壁菌种也对氨基酸表现出高活性,特别是肠单胞菌(Intestinimonas AF211),它通过不同的途径将葡萄糖和赖氨酸发酵成丁酸。
形成丁酸盐的微生物途径
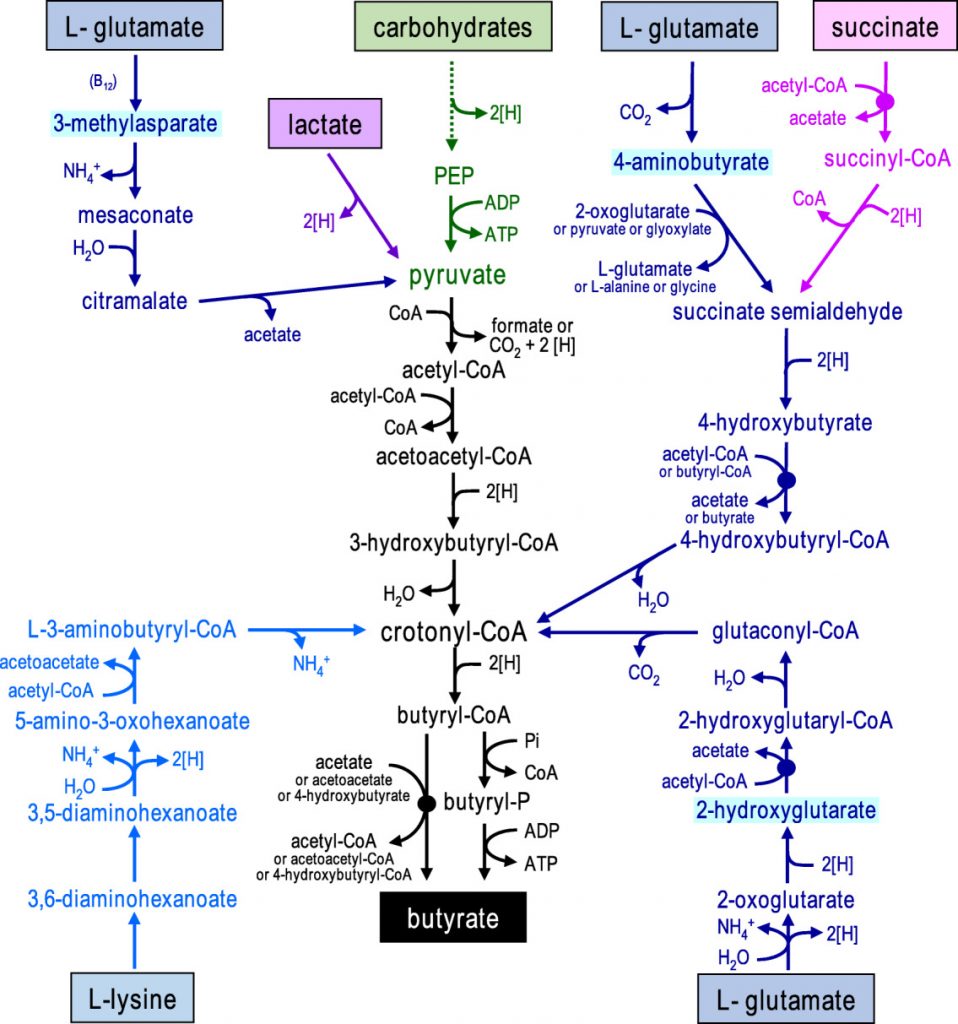
doi: 10.1111/1462-2920.13589.
碳水化合物通过糖酵解发酵为丙酮酸显示为绿色,从乙酰辅酶A形成丁酸盐显示为黑色,氨基酸发酵途径显示为蓝色(以中间体命名不同的谷氨酸途径),乳酸和琥珀酸发酵分别显示为紫色和粉色。
• 人结肠中的瘤胃球菌和毛螺菌是产丁酸的主力
除食源性丁酸,人体内丁酸主要是由盲肠和结肠的厌氧型细菌发酵产生,而由胃和小肠产生的丁酸含量极低。大肠产丁酸的菌种主要是梭菌属XIVa和IV族,以及真杆菌属和梭杆菌属
盲肠和结肠中丁酸的产生速度和数量主要取决于肠道微生物组成、日常膳食中可利用发酵成分组成等。在人的结肠中,厚壁菌门中的瘤胃球菌科(Ruminococcaceae)和毛螺菌科(Lachnospiraceae)这两个主要科,以及包括丹毒丝菌科(Erysipelotrichaceae)和梭菌科(Clostridiaceae)在内的其他厚壁菌门物种,都发现了丁酸生产菌种与非生产菌种的交替存在。
但应注意,许多人类结肠的主要厚壁菌门(例如Blautia spp.、Eubacterium eligens、Ruminococcus spp.)缺乏从碳水化合物中生成丁酸的能力。
产生丁酸盐的细菌被认为在生命的第一年内定植于宿主,并且在成年时占总细菌群落的20%以上。在消化道中发现的已知丁酸盐生产者中,大多数似乎属于毛螺菌科和瘤胃球菌科。我们将简要介绍两个丁酸生产菌。
▸ 普拉梭菌
普拉梭菌(Faecalibacterium prausnitzii)是健康人类微生物群中最丰富的菌种之一,也是主要的丁酸盐生产者之一。它通过丁酰辅酶A:乙酸辅酶A转移酶产生丁酸,并消耗乙酸。
虽然F.prausnitzii菌株是专性厌氧菌,但在核黄素(维生素B2)和还原性化合物(如半胱氨酸或谷胱甘肽)存在的情况下,低浓度氧气也能促进其生长。
氧气消耗会伴随丁酸形成的减少。F. prausnitzii分离株在利用食物多糖(如淀粉和半纤维素)生长方面能力有限,但一些菌株可以利用菊粉和果胶衍生物,并且普遍利用糖醛酸。
• 肠道炎症患者产生丁酸会减少
F. prausnitzii在炎症性肠病患者中很少见,尤其是克罗恩病,有证据表明它具有抗炎作用,因此引起了人们对其作为潜在治疗药物的兴趣。同样,据报道,Butyricicoccus pullicaecorum在炎症性肠病患者中较少见,并且也可能具有治疗潜力。
注:其他瘤胃球菌科细菌也能产生丁酸,但人们对大多数此类生物知之甚少。关于Faecalibacterium prausnitzii,详见:肠道核心菌属——普拉梭菌(F. Prausnitzii),预防炎症的下一代益生菌
▸ 毛螺菌科
直肠真杆菌(Eubacterium rectale)和密切相关的罗氏菌属(Roseburia)构成了产丁酸毛螺菌科的主要群体,它们具有相同的丁酰辅酶 A:乙酸辅酶 A 转移酶途径来生产丁酸,并且其丁酸合成基因的基因组组织也相同,从乙酰辅酶A到丁酰辅酶A。
• 罗氏菌属通过代谢饮食多糖产生丁酸
在某些Roseburia菌株中,特别是在弱酸性pH值下,丁酸几乎是唯一产生的发酵酸,乙酸的净消耗通常伴随着丁酸的形成。一些其他菌株和物种除了产生丁酸外,还产生甲酸和乳酸。基因组分析表明,该群体具有相当大的利用饮食来源的多糖的能力,包括淀粉、阿拉伯木聚糖和菊粉,不同菌株和物种之间的差异很大。
其他拥有丁酰辅酶A:乙酸辅酶A转移酶基因的毛螺菌科包括Eubacterium hallii、Anaerostipes hadrus、Coprococcus catus、与分离株SS3/4和M62/1有关的未鉴定物种。
• 一些毛螺菌科细菌利用乳酸和乙酸来产生丁酸
某些毛螺菌科能够在乳酸和乙酸盐存在下生长并产生丁酸,其总净化学计量为4mol乳酸和2mol乙酸盐产生3mol丁酸 。
这包括仅使用D-乳酸的物种Anaerostipes hadrus和能够利用两种乳酸异构体的E. hallii。
此外,双歧杆菌等常见益生菌配方中的微生物与丁酸盐生产者之间的交叉喂养相互作用已被证明是可以产丁酸盐的。
人体肠道中的主要丁酸生产者
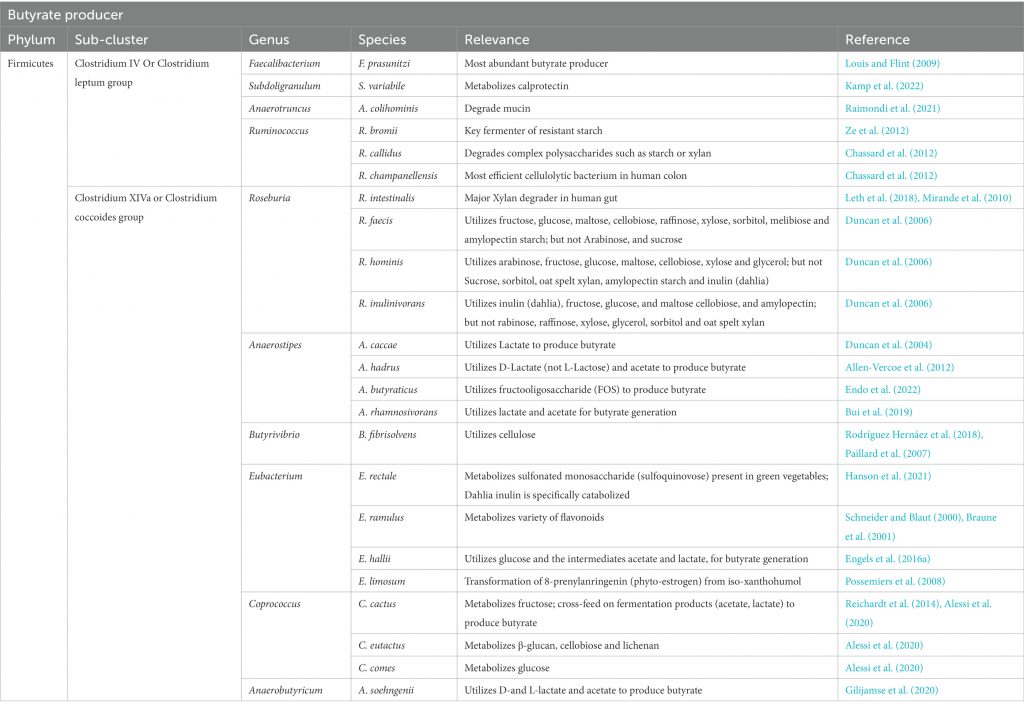
doi: 10.3389/fmicb.2022.1103836.
▸ 增加丁酸盐生成的食物
饮食对人体微生物群有显著影响,通过饮食干预可显著改变细菌数量并增加微生物多样性。富含纤维的饮食对丁酸盐的产生特别有益,因为它可以滋养产生丁酸盐的细菌。
• 高纤维饮食能够有效增加丁酸盐的生成
蔬菜、水果、豆类和全谷物,它们含有膳食纤维,可被肠道细菌发酵成丁酸盐等有机化合物。通过食物增强微生物群是促进消化系统健康和丁酸盐生成的有效且安全的方法。因此,增加丁酸盐产量的最佳方法是通过高纤维饮食。
• 高脂、低纤维饮食不利于丁酸盐的生成
高蛋白、高脂肪、低碳水化合物的饮食已被证明会破坏微生物组中丁酸盐的产生。在一项研究中,研究人员分析了短期饮食限制碳水化合物摄入量的肥胖参与者的微生物组,从而限制了他们对植物性膳食纤维的消耗。
在低碳水化合物饮食(每天24克)和中等碳水化合物饮食(每天164克)4 周后,短链脂肪酸的浓度低于高碳水化合物饮食(每天399克)。具体来说,当碳水化合物摄入量减少时,丁酸盐浓度会降低。
同时还发现,厚壁菌门细菌Roseburia和E.Rectale的密度与丁酸盐浓度之间存在联系,两者都随着碳水化合物摄入量的减少而降低。
一些食源性物质产生短链脂肪酸的量
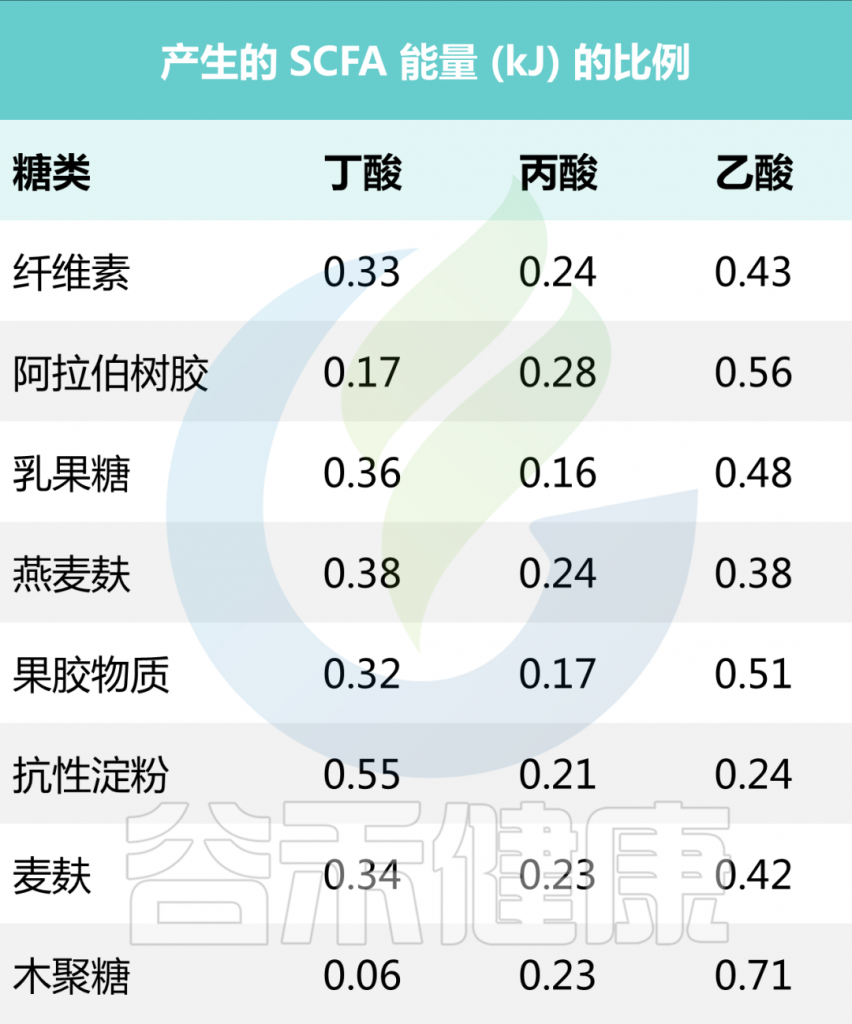
• 抗性淀粉
研究表明,从生马铃薯、高直链淀粉玉米和全谷物中提取的阿拉伯木聚糖中,含有高含量2型抗性淀粉的食物显著增加了丁酸盐的产量。
在174名健康年轻人的饮食中添加马铃薯抗性淀粉后,丁酸盐产量增加。玉米、菊苣和玉米中的抗性淀粉也进行了测试,但只有当食用土豆中的抗性淀粉时,粪便中的丁酸总量才会显著增加。
• 果聚糖(菊粉)
许多研究表明,菊粉可以增加短链脂肪酸的产量,包括丁酸盐。这可能解释了香蕉在溃疡性结肠炎和克罗恩病患者的饮食中如此有效的原因之一。然而,链长较短的果聚糖通常比链长的果聚糖(如菊粉)更容易喂养产丁酸菌。
也就说,抗性淀粉和果聚糖(短链低聚果糖和长链菊粉)在肠道中发酵时会产生丁酸盐。
富含抗性淀粉的食物:全麦面包、燕麦、大麦和糙米等全谷物;黑豆、红豆、绿豆等豆类;马铃薯、玉米、菊苣根、牛蒡根、魔芋根、亚麻籽。
富含果聚糖的食物:洋葱、菊苣、香蕉、朝鲜蓟、芦笋、大蒜、韭菜、西兰花、开心果。
需要注意的
由于人与人之间的常驻微生物存在显著差异,并且它们消化某些底物的能力不同,因此比较明智的选择是食用包含多种抗性淀粉和果聚糖的饮食。例如以马铃薯抗性淀粉、燕麦麸皮纤维或车前草种子或短链低聚半乳糖、长链低聚果糖和谷氨酰胺的混合物的形式短期补充,增加丁酸盐水平。
丁酸盐作为一种重要的的调节因子,是宿主-微生物串扰的关键介体。丁酸盐在体内可以通过脂肪酸氧化为机体供应能量,是肠道上皮细胞的主要供能物质。
丁酸盐还与机体健康密切相关,对调节肠道健康、修复肠道屏障、抑制炎症及癌症等病症意义重大。在养殖业中常添加丁酸盐保护动物健康生长,如预防断奶仔猪腹泻、调节鸡肠道菌群并增强其免疫力等。
▸ 为肠道细胞提供能量
丁酸盐是结肠细胞的主要能量来源,结肠细胞是构成肠道内壁的细胞。
与身体中使用糖(葡萄糖)作为主要能量来源的大多数其他细胞不同,肠道内壁细胞(结肠细胞)主要使用丁酸盐。如果没有丁酸盐,这些细胞就无法正确执行其功能。
• 丁酸盐为结肠细胞提供能量,细胞利于产丁酸盐细菌的生长
厚壁菌属的成员以产生丁酸盐而闻名,像Roseburia,Faecalibacterium prausnitzii,直肠真杆菌(E.rectale)等。
这种关系是相互的。丁酸盐为结肠细胞提供燃料,作为回报,这些细胞有助于提供一个无氧环境,有益的肠道微生物在其中茁壮成长。这可以控制炎症,保持肠道细胞健康,并使肠道细菌保持健康。
▸ 促进肠道运动
实验研究表明,丁酸盐通过作为短链脂肪酸受体的配体和激活剂,诱导肠道激素肽YY或介导肠嗜铬细胞释放5-羟色胺来促进肠道运动。
丁酸盐还可以通过上调Na+—H+交换器和诱导ATPase离子交换器基因来增强水和电解质的吸收。并且可能有益于预防某些类型的腹泻。
▸ 抗炎、抗癌特性
丁酸盐对肠道具有抗炎和抗癌功能。
肠道内壁会保持低水平的炎症,以防与微生物群接触的粘膜表面发生任何变化。低水平的炎症受到严格控制,但如果它被破坏,会导致氧化损伤,并可能在很长一段时间内导致癌症。
• 丁酸盐减少促炎细胞因子的产生
丁酸盐会阻止体内的一些促炎物质发挥作用。丁酸盐的抗炎作用可减少氧化应激并控制自由基造成的损害。
丁酸盐的抗炎特性,部分原因是其抑制核转录因子(NF-κB)的激活,通过下调NF-κB信号通路,丁酸盐可以调节促炎细胞因子的产生。
• 丁酸盐阻止结肠癌细胞的生长
丁酸盐也是一种组蛋白脱乙酰酶(HDAC)抑制剂。组蛋白脱乙酰酶是大多数癌症中产生的酶。因为丁酸盐是一种抑制剂,它实际上会改变基因表达,抑制细胞增殖,诱导细胞分化或凋亡。因此,它可以阻止癌细胞的发展。
有证据表明,结直肠癌(CRC)患者的微生物组图显示主要产丁酸菌属减少,包括罗氏菌属(Roseburia)、Clostridiales、Faecalibacterium和Lachnospiraceae科成员,而使用产丁酸的丁酸梭菌可有效减少癌细胞增殖并增强癌细胞凋亡。
▸ 修复肠道屏障,防止肠漏
肠上皮细胞的屏障功能是重要的第一道防线,确保上皮层具有适当的通透性。研究发现丁酸盐能够修复和增强肠上皮细胞的屏障功能。
• 丁酸盐上调粘蛋白的表达,增强粘液屏障
最新研究表明,丁酸对肠屏障功能具有保护作用。例如,丁酸能够上调粘蛋白2(MUC2)的表达。粘蛋白2是肠粘膜表面主要的粘蛋白,可增强粘膜层,从而限制有害物质通过肠道。
• 丁酸盐对肠道屏障的影响可能具有浓度依赖性
此外,丁酸还能增加三叶因子(TFF)的表达,这是一种粘蛋白相关肽,有助于维持和修复肠粘膜。丁酸还调节紧密连接蛋白的表达,以降低细胞旁通透性。其中一种机制是通过激活单分子层中的AMP活化蛋白激酶。
体外模型研究表明,丁酸对肠道屏障功能的影响可能具有浓度依赖性。丁酸在低浓度(≤2 mM)时可促进肠道屏障功能,但在高浓度(5或8 mM)时可能通过诱导细胞凋亡来破坏肠道屏障功能。
根据哺乳动物胃肠道中的生理浓度,目前体外模型中推荐使用的丁酸浓度为0-8mM。然而,考虑到大多数丁酸在结肠上皮中被代谢为能量底物,体内和体外模型中治疗剂量可能存在差异。
▸ 抗氧化能力
说起抗氧化,我们先了解一下自由基。它基本上是体内化学反应产生的废物。另一方面,抗氧化剂是身体抵御它们的防御措施。大量自由基会造成损害并压倒身体的修复系统。我们称之为氧化应激。氧化应激被认为是导致衰老和疾病的一个重要因素。
• 丁酸盐增加谷胱甘肽,可以中和自由基
结肠或大肠是身体产生的废物的储存容器。较高的丁酸盐水平已被证明会增加谷胱甘肽的水平,谷胱甘肽是一种在人体细胞中产生的抗氧化剂,可以中和肠道中的自由基(自由基与炎症和许多疾病有关),从而起到抗氧化作用。
▸ 调节肠道免疫
• 维持厌氧环境,阻止有害菌生长
丁酸通过增强结肠细胞的氧消耗和稳定缺氧诱导因子(HIF)来维持结肠内的厌氧环境,而丁酸的缺失则会促进潜在有害细菌和分子的积聚,例如沙门氏菌、大肠杆菌和一氧化氮(NO)。
肠道中丁酸水平降低,这促进了肠道上皮氧合和鼠伤寒沙门氏菌(S.Typhimurium)的生长,鼠伤寒沙门氏菌是食源性肠道炎症和腹泻的已知病因。
• 影响免疫细胞迁移、粘附
除了营造无氧环境外,短链脂肪酸,尤其是丁酸盐,还可以作为免疫细胞趋化和粘附的调节剂。丁酸可以调节肠上皮细胞介导的中性粒细胞向炎症部位的迁移,并且这种作用是浓度依赖性的。
此外,丁酸在细胞增殖和凋亡中发挥作用。丁酸刺激细胞生长和DNA合成,并诱导细胞周期G1期的生长停滞。虽然低浓度的丁酸会增强细胞增殖,但高浓度的丁酸会诱导细胞凋亡。
丁酸盐等短链脂肪酸可以在先天反应过程中通过影响巨噬细胞和粒细胞以及树突状细胞的抗原呈递发生,也可能在适应性免疫反应过程中通过影响T细胞和B细胞功能发生。
短链脂肪酸的免疫调节作用
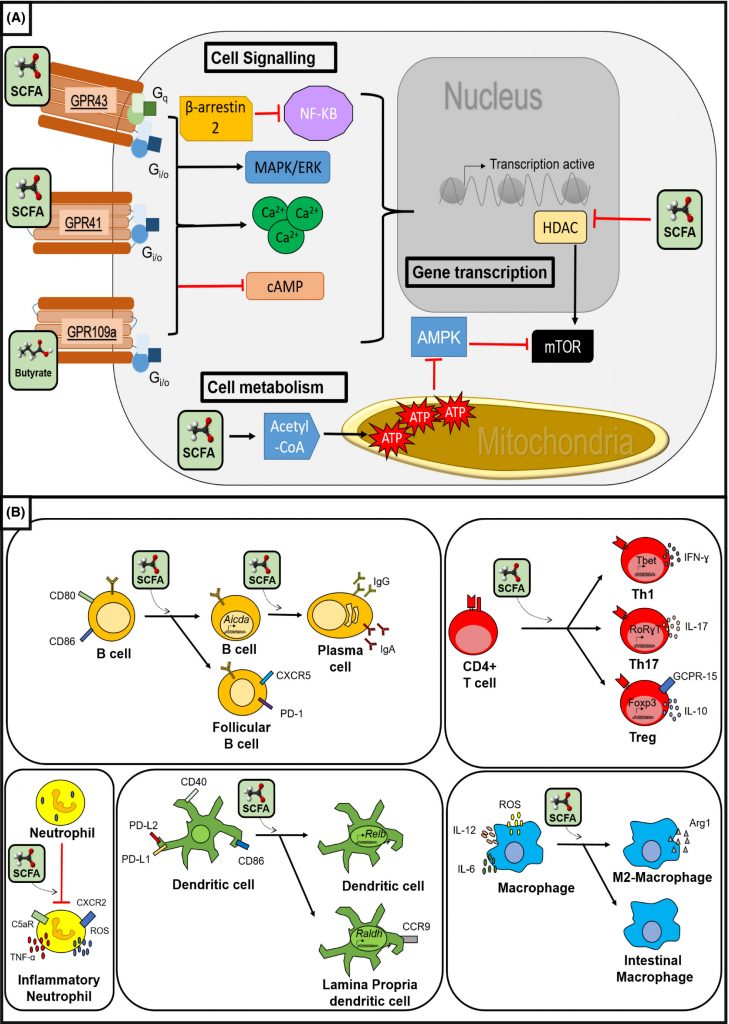
doi: 10.1111/apt.14689.
▸ 影响代谢健康
• 对肥胖存在一定影响,但还不能完全确定
包括丁酸盐在内的短链脂肪酸可通过激活肠细胞内的FFAR来降低食欲和体重。这促进胰高血糖素样肽1(GLP-1)和肽YY的释放,前者促进胰岛素分泌并抑制胰高血糖素分泌,后者降低食欲并减缓胃排空。
短链脂肪酸还能减少所谓的“饥饿激素”——胃促生长素(ghrelin)的分泌;FFAR2存在于ghrelin分泌细胞上,包括丁酸盐和丙酸盐在内的FFAR2激动剂可减少ghrelin分泌。
丁酸盐影响脂质代谢的机制
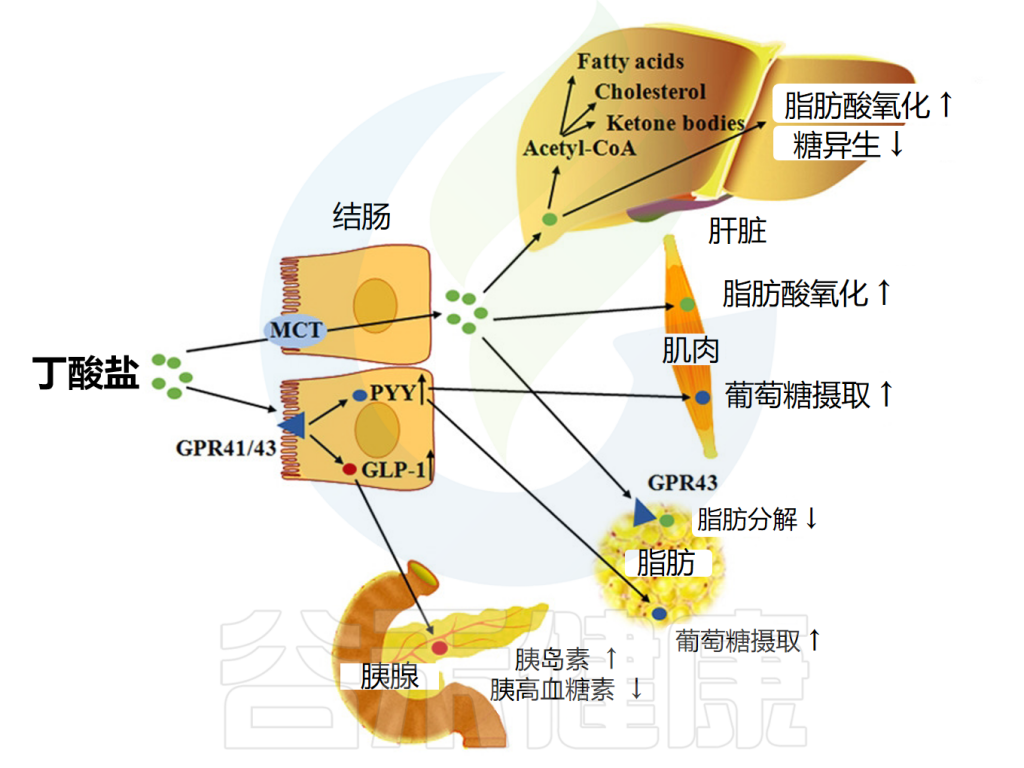
doi: 10.1093/advances/nmx009.
也有证据表明短链脂肪酸作用于交感神经系统,交感神经节中FFAR3的激活导致能量消耗增加。
然而,与丙酸盐一样,关于丁酸盐对食物摄入的影响,有相互矛盾的结果报道。
• 丁酸盐对控制血糖水平可能有潜在好处
研究发现,糖尿病患者和糖尿病前期受试者中丁酸盐水平降低。
丁酸在肠内分泌细胞(EEC)中与游离脂肪酸受体(FFAR)FFAR2和FFAR3结合,调节肠道激素释放,如胰高血糖素样肽1(GLP-1)和肽YY(PYY)。
丁酸盐通过多种途径影响糖代谢的调节
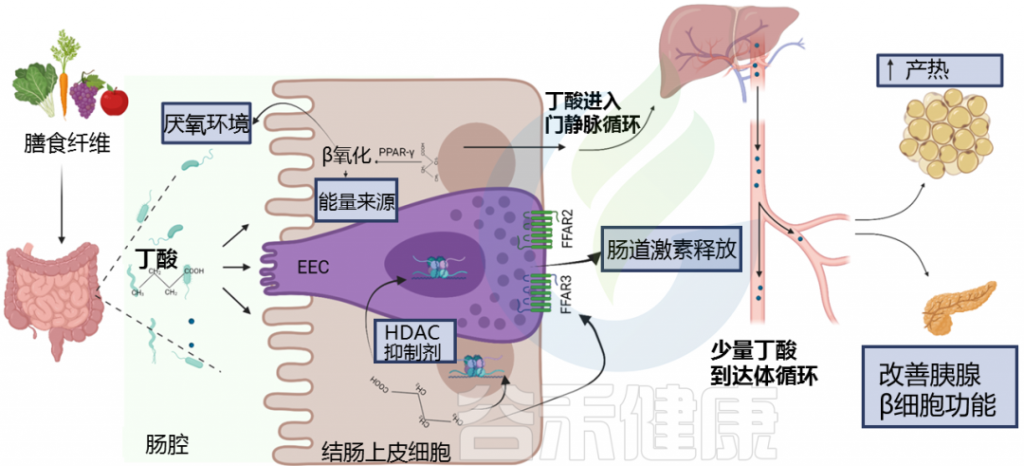
doi: 10.3389/fendo.2021.761834.
这些激素共同作用以保持血糖水平稳定。当血糖过高时,胰岛素会告诉身体的肌肉和脂肪细胞吸收多余的葡萄糖,所以说这些激素对肥胖和糖尿病很重要。丁酸盐增加这些肠道激素的释放,表明对控制血糖水平和防止体重增加有潜在的好处。
!
丁酸盐过低可能会导致以下危害:
•肠道屏障功能受损:丁酸盐有助于维持结肠的“生理性缺氧”,并提供能量给结肠细胞,有助于维持肠道稳态。丁酸盐的减少可能导致肠道屏障功能受损。
•增加慢性疾病风险:变形菌门的兼性厌氧细菌的扩张几乎总是伴随着产丁酸盐细菌丰度的减少,并与许多慢性疾病有关,包括炎症性肠病、肠易激综合症、结直肠癌、2型糖尿病、肥胖等。
•神经炎症:丁酸盐具有抗炎特性,对大脑健康具有重要意义。丁酸盐过低可能导致炎症增加,影响大脑健康。
•睡眠问题:丁酸盐可能作为细菌源性促睡眠信号,丁酸盐过低可能影响睡眠,导致睡眠问题。
•肠道菌群失衡:丁酸盐过低可能导致肠道菌群失衡,因为变形菌门的扩张伴随着产丁酸盐细菌的减少。
•不利于肠道损伤修复:在轻微炎症或轻度溃疡部位添加丁酸盐可以促进肠道损伤的修复,而丁酸盐过少可能不利于溃疡的恢复。
!
丁酸盐并不总是越多越好,低浓度丁酸促进细胞增殖和生长,高浓度丁酸反而抑制细胞增殖和生长,增加肠道的通透性。
• 不同部位对丁酸盐的耐受阈值存在差异
胃肠道不同部位对丁酸盐的耐受阈值也存在一定差异,胃和小肠对丁酸的耐受阈值低,结肠和盲肠耐受阈值高。
添加普通丁酸钠制剂(主要在肠道前端被吸收利用),反而造成肠道炎症、菌群失调。
• 肠道严重溃疡时丁酸盐可能加剧症状
更有意思的是,轻微炎症或者轻度溃疡部位添加丁酸盐可以促进肠道损伤的修复,在严重溃疡肠道部位添加,不利于溃疡的恢复,甚至加剧整个溃疡。
有学者发现溃疡部位的粘膜组织对丁酸的代谢降低,甚至只有正常粘膜组织的一半,主要是由于其转运载体和氧化相关的酶活降低。
主要参考文献
Louis P, Flint HJ. Formation of propionate and butyrate by the human colonic microbiota. Environ Microbiol. 2017 Jan;19(1):29-41.
Gill PA, van Zelm MC, Muir JG, Gibson PR. Review article: short chain fatty acids as potential therapeutic agents in human gastrointestinal and inflammatory disorders. Aliment Pharmacol Ther. 2018 Jul;48(1):15-34.
Hosseini E, Grootaert C, Verstraete W, Van de Wiele T. Propionate as a health-promoting microbial metabolite in the human gut. Nutr Rev. 2011 May;69(5):245-58.
Arora T, Tremaroli V. Therapeutic Potential of Butyrate for Treatment of Type 2 Diabetes. Front Endocrinol (Lausanne). 2021 Oct 19;12:761834.
Singh V, Lee G, Son H, Koh H, Kim ES, Unno T, Shin JH. Butyrate producers, “The Sentinel of Gut”: Their intestinal significance with and beyond butyrate, and prospective use as microbial therapeutics. Front Microbiol. 2023 Jan 12;13:1103836.
Gonzalez-Garcia, R.A.; McCubbin, T.; Navone, L.; Stowers, C.; Nielsen, L.K.; Marcellin, E. Microbial Propionic Acid Production. Fermentation 2017, 3, 21.
Reichardt N, Duncan SH, Young P, Belenguer A, McWilliam Leitch C, Scott KP, Flint HJ, Louis P. Phylogenetic distribution of three pathways for propionate production within the human gut microbiota. ISME J. 2014 Jun;8(6):1323-35.
Liu H, Wang J, He T, Becker S, Zhang G, Li D, Ma X. Butyrate: A Double-Edged Sword for Health? Adv Nutr. 2018 Jan 1;9(1):21-29.

谷禾健康

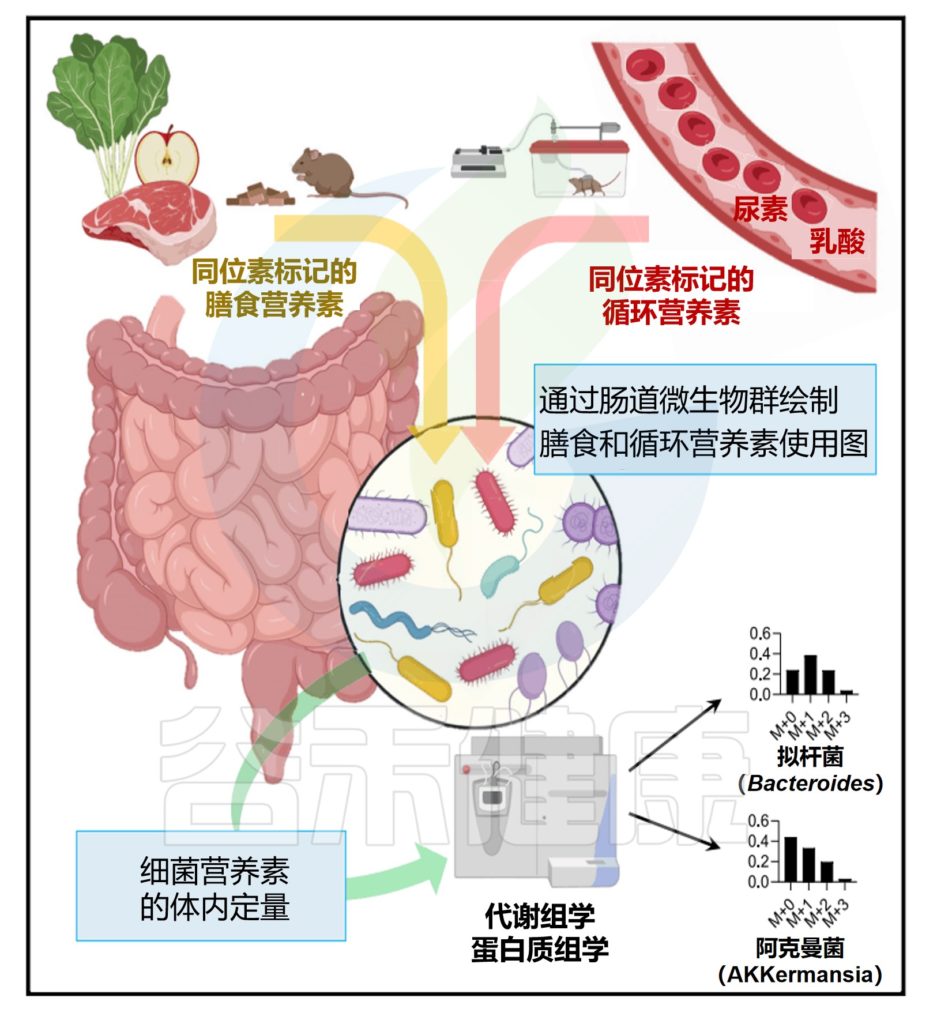
随着一日三餐米面肉蛋菜等一些列食物的食用,数百种化学成分会进入我们的消化道。在那里,它们被肠道微生物组进一步代谢,这是数千种微生物物种的独特集合。
因此,肠道微生物组在决定营养如何影响健康方面发挥着重要作用。然而到目前为止,微生物组中的许多微生物的代谢能力仍然是未知的。这意味着我们不知道它们以什么物质为食,以及它们是如何处理这些物质的。
近期,来自普林斯顿大学的研究人员在《CELL》期刊上发表了最新的文章:
“Gut bacterial nutrient preferences quantified in vivo”,研究人员使用同位素追踪定量研究了小鼠肠道微生物群的输入和输出。

微生物碳水化合物发酵的主要输入是膳食纤维,支链脂肪酸和芳香代谢物的主要输入为膳食蛋白质。此外,循环宿主乳酸、3-羟基丁酸和尿素(但不是葡萄糖或氨基酸)为肠道微生物群提供食物。
肠道菌群拥有巨大的酶多样性,超过哺乳动物基因组的数量100多倍。这些酶的能力能使摄入的膳食营养物质加工成一些列微生物代谢物。
为了复制自身和释放代谢产物,肠道细菌需要营养输入。这些形式包括摄入的食物、宿主合成的肠道粘液和宿主循环代谢物。
//
在本文中,研究人员通过对肠道菌群及其进入宿主循环系统的代谢物进行了大规模的定量评估。
研究了膳食淀粉、纤维和蛋白质的贡献以及宿主粘液的贡献,也研究了大多数主要的循环宿主营养素,发现乳酸、3-羟基丁酸和尿素在从宿主传递到肠道微生物群中表现突出。基于对细菌特异性肽序列的测量,评估了不同细菌属的营养偏好,并表明这些偏好与响应改变饮食的微生物组分变化一致。
同位素追踪能够定量测量代谢物和生物量的输入。与质谱检测相结合的稳定同位素示踪剂,使得能够测量特定下游产物的标记。通过注入氮标记的苏氨酸来标记宿主粘液,研究人员能够比较饮食和粘液蛋白对肠道微生物群的贡献,并观察到喂食低蛋白饮食的小鼠中粘液贡献的变化。
从小鼠尾部静脉抽取血样;
使用注射器从小鼠膀胱采集尿液;
所有血清样品在没有抗凝剂的情况下置于冰上 15 分钟,并在 4°C 下以 16,000 x g 离心 15 分钟。
用预冷的Wollenberger钳在液氮中快速分离并快速冷冻(< 5秒)获得组织;夹紧前取出肠内容物;盲肠内容物取样时,先将小鼠盲肠取出并在表面切开,然后用镊子将盲肠内容物挤出。
取新鲜粪便,轻揉小鼠腹部诱导排便。将血清、组织和粪便样本保存在 -80 ºC 直至进一步分析。
为了测定血清和组织样本中的代谢物浓度,进行了同位素标配(isotope spike-in)或标准标配(standard spike-in )。
对于前者将已知浓度的同位素标记标准品加入血清或组织提取液中,通过标记与未标记代谢物的比值计算浓度。
当没有同位素标准品时,加入连续稀释的非标记标准品,测量的总离子计数与加入的标准品浓度之间产生线性拟合。然后通过拟合线的x截距确定内源代谢物的浓度;蛋白质氨基酸组成采用酸水解法测定。
首先,使用13C同位素标记的不同营养物质,通过口服管饲法对小鼠进行灌胃采集小鼠的血清、组织和粪便样本。对粪便和肠内容物进行16S rRNA测序获得细菌分类。
首先使用代谢组学方法测定盲肠内容物中游离氨基酸13C-或15N标记。
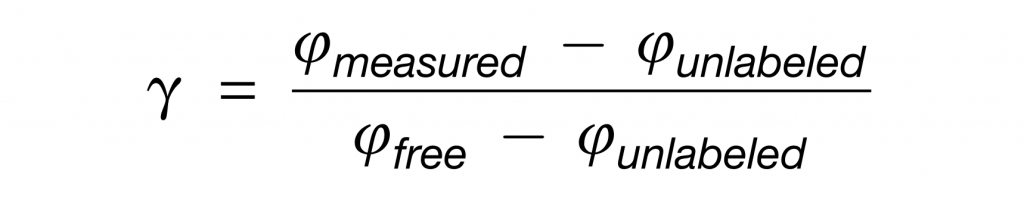
然后,对于每个肽,模拟了未标记(Iunlabeled)和由游离盲肠氨基酸(Ifree)合成的肽的同位素包膜模式。标量γ可以通过将测量的肽同位素分布(Imeasured)与Iunlabeled和Ifree的线性组合拟合来确定。
注意,当一个菌属使用的特定营养素超过该营养素对盲肠游离氨基酸的贡献时,γ将大于1。
具体来说,测量的每个肽的γ如下:

对于细菌属水平的原料贡献程度的测量,分析中只保留测量超过3个肽的属,多肽的中位数为γ-genus。
对于细菌科水平,仅分析在蛋白质组学中始终检测到的属,以及在 16S rRNA 基因扩增子测序中检测到 (> 0.5%) 的属的上一级科。
每种营养物质对菌属的贡献程度的定量公式如下:
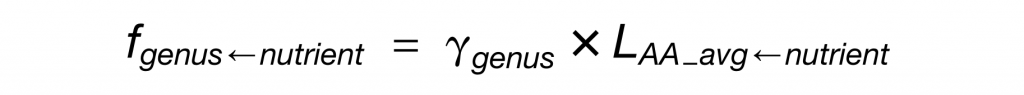

LAA_avg-nutrient为各营养物质对细菌蛋白质的贡献程度,其计算公式如下:
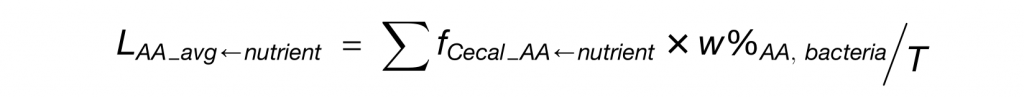

1 微生物组消耗较少的可消化膳食成分
微生物群影响宿主生理学的主要机制是通过分泌代谢产物。研究人员在门静脉和体循环以及盲肠内容物中测量了微生物衍生的50多种代谢产物的绝对浓度。
微生物群相关代谢物的绝对浓度和来源
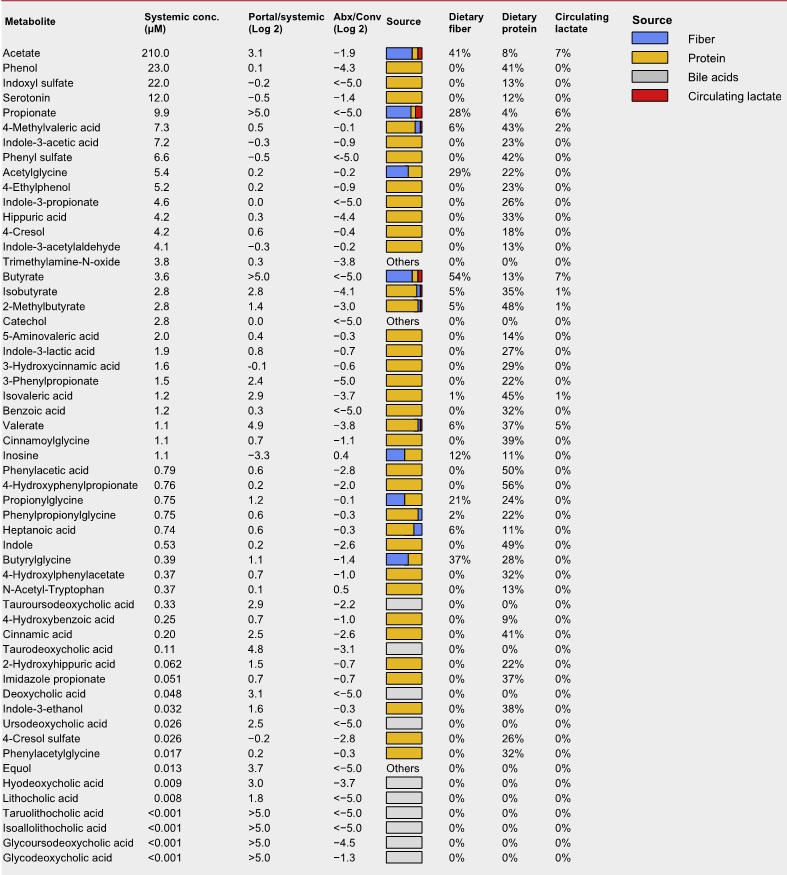
上表可以看到,与全身血液相比,大多数在门静脉循环中升高,除两种(肌苷和N-乙酰色氨酸主要来源于宿主)外,其余均被抗生素治疗耗尽。
门静脉血中主要排泄产物是短链脂肪酸。
其他相对丰富的微生物群产物是芳香族氨基酸发酵产物(苯酚、吲哚硫酸盐和3-苯丙酸盐)和支链脂肪酸(戊酸盐、异戊酸盐,4-甲基戊酸、异丁酸盐和2-甲基丁酸盐)。
探索肠道微生物产物的膳食输入:淀粉、菊粉
研究人员通过口服管饲法、淀粉(易消化葡萄糖聚合物)和菊粉(易消化果糖聚合物,即可溶性纤维)喂养小鼠:
13C淀粉灌胃后,标记的葡萄糖、乳酸和丙氨酸迅速出现在门脉循环中,并占大多数淀粉碳(约75%)。
13C菊粉和13C淀粉有什么不同?
13C菊粉灌胃后,没有观察到大量标记的果糖、葡萄糖、乳酸和丙氨酸,取而代之的是标记的门静脉代谢产物以短链脂肪酸的形式缓慢出现,约40%的菊粉碳成为短链脂肪酸,其余未消化并随粪便排出。
膳食菊粉,而不是淀粉,在盲肠内容物中广泛标记糖酵解和TCA中间体和氨基酸。
藻类蛋白大量标记了微生物群衍生的门静脉代谢物:短链脂肪酸、支链脂肪酸和芳烃(吲哚、吲哚-3-丙酸盐和3-苯丙酸盐)。
“难以消化的碳水化合物和蛋白质直接为微生物组提供营养,并通过微生物产物间接为宿主提供营养。”
研究中发现宿主循环系统中的乳酸,3-羟基丁酸以及尿素能为肠道细菌提供营养。
如图A,将同位素标记的营养物质通过静脉输注到小鼠的全身血液循环中。 2.5 小时后收集血清和粪便以量化每种营养物质对相应菌群代谢物的碳贡献。
图BCD表示了13C标记的各种营养物质在小鼠的血液和粪便中的含量,可见乳酸和 3-羟基丁酸有进入肠道菌群中,而其余大部分营养物质如柠檬酸盐、葡萄糖、氨基酸等都没有进入到肠道菌群中。
图F为15N标记的营养物质,可见尿素也同样被菌群大量利用。
为了定量确定微生物代谢物的来源,研究人员给小鼠喂食部分纤维、脂肪或蛋白质13C标记的标准食物,盲肠标记在12小时内达到稳定状态。
为了说明循环营养输入,研究人员还注入了13C乳酸或3-羟基丁酸。
这些研究确定了大多数微生物群中心代谢物中的碳供给:
接下来,研究人员检查了微生物组游离氨基酸的输入,并用15N标记的膳食蛋白和注入的尿素进行追踪。
与哺乳动物不同,大多数肠道细菌具有合成所有20种蛋白质氨基酸的生物合成能力。
然而,研究人员观察到“必需氨基酸”主要来源于膳食蛋白质,哺乳动物无法制造,需要在细菌中表达广泛的生物合成途径。
“非必需氨基酸”主要在肠道微生物群中合成,使用膳食菊粉和循环乳酸作为碳源。
抗生素或无菌小鼠中的微生物群消耗有利于盲肠中氨基酸的积累(基于同位素追踪研究),这些氨基酸主要来自膳食蛋白质和微生物合成的氨基酸的消耗。
膳食蛋白质是必需氨基酸和非必需氨基酸的主要氮源,宿主尿素对非必需氨基酸也有很大贡献。
研究人员的发现如下:
【1】必需氨基酸,尽管能够由微生物群合成,但主要来自饮食,不经历任何碳重排;
【2】与TCA连接最紧密的非必需氨基酸基本上由微生物群合成,使用来自纤维的碳,通过中心代谢反应与其他碳争夺;
【3】转氨反应部分地将来自饮食衍生氨基酸的氮与来自宿主尿素的氮混合。
研究人员发现,许多微生物来源的代谢物来源于到达结肠的未吸收膳食蛋白。假设这些代谢物的循环水平将取决于膳食蛋白质到达结肠微生物群的程度。
为了控制这一点,研究人员给小鼠喂食的食物中,一部分蛋白质(酪蛋白,部分到达结肠微生物群)被游离氨基酸(基本上在小肠中完全吸收)取代。
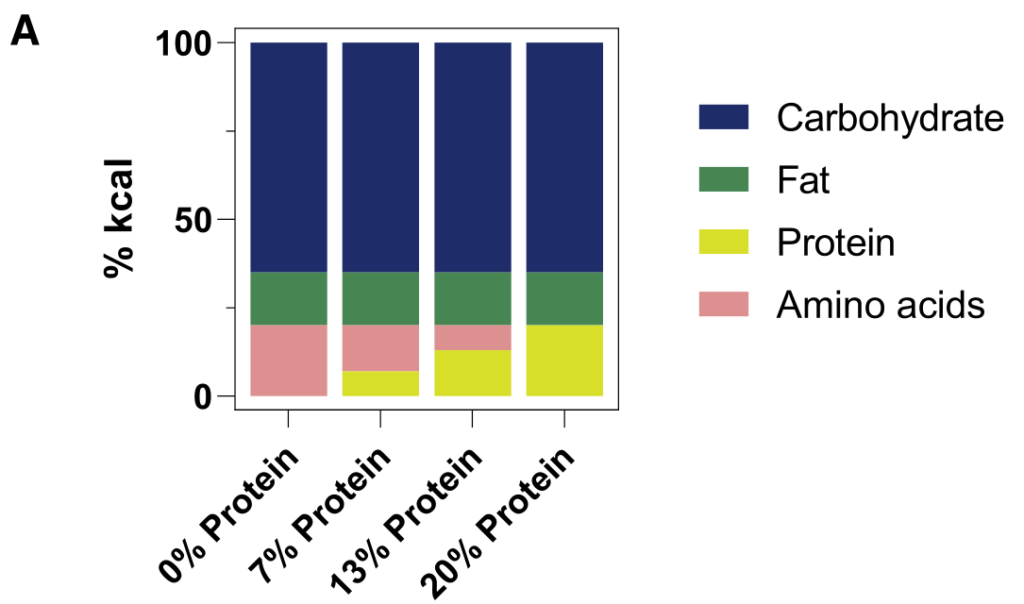
2周后对全身血液进行代谢组学研究。含有较少完整蛋白质和更多游离氨基酸的饮食往往会增加循环氨基酸水平。
重要的是,蛋白质衍生的循环微生物代谢物(酚类、吲哚类和酰基甘氨酸)串联下降。
“微生物代谢物营养来源的知识可用于操纵其系统水平。”
研究人员通过结合13C营养标记和蛋白质组学来定量不同微生物的碳原料。
每种13C标记的营养素(膳食菊粉、膳食藻蛋白或循环乳酸)提供24小时,这足以在肠道细菌中实现稳态标记。
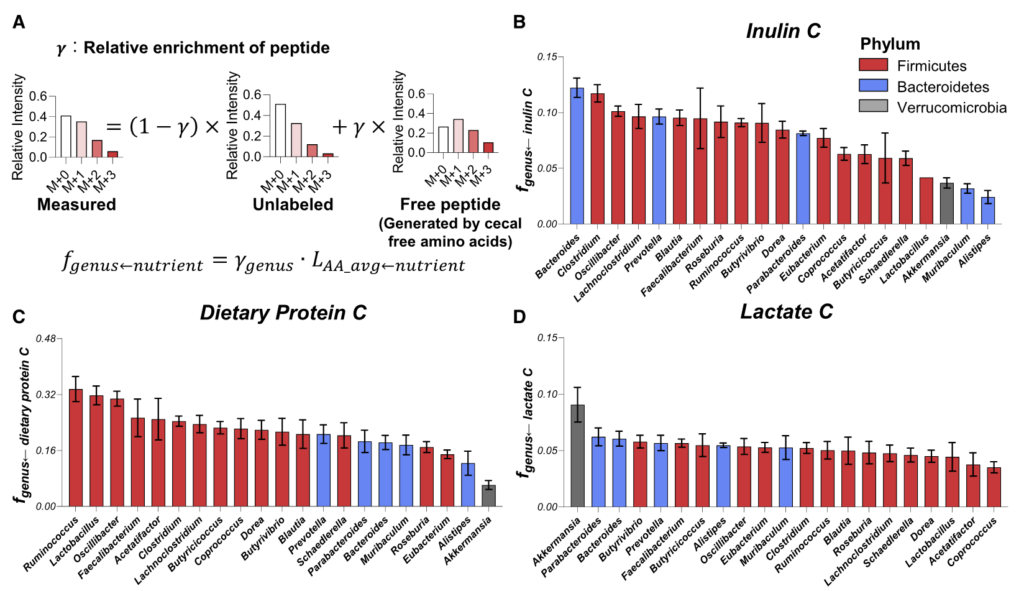
如同B-D,分别计算了在膳食中使用的菊粉和蛋白质以及乳酸在各细菌内的喜好程度,这个喜好程度也就是将在细菌特异性肽上被同位素标记的程度进行了量化。
结果可见:
拟杆菌属和梭状芽胞杆菌利用菊粉的程度是 Akkermansia、Muribaculum 或 Alistipes 的 4 倍多。
总体而言,厚壁菌门下的菌属比拟杆菌门的使用膳食中的蛋白质(厚壁菌0.237±0.052;拟杆菌0.175±0.031,p=0.02)。
Akkermansia通常被认为是一种促进健康的肠道微生物,使用的菊粉和蛋白质最少。相比之下,它使用了来自宿主的循环乳酸最多。
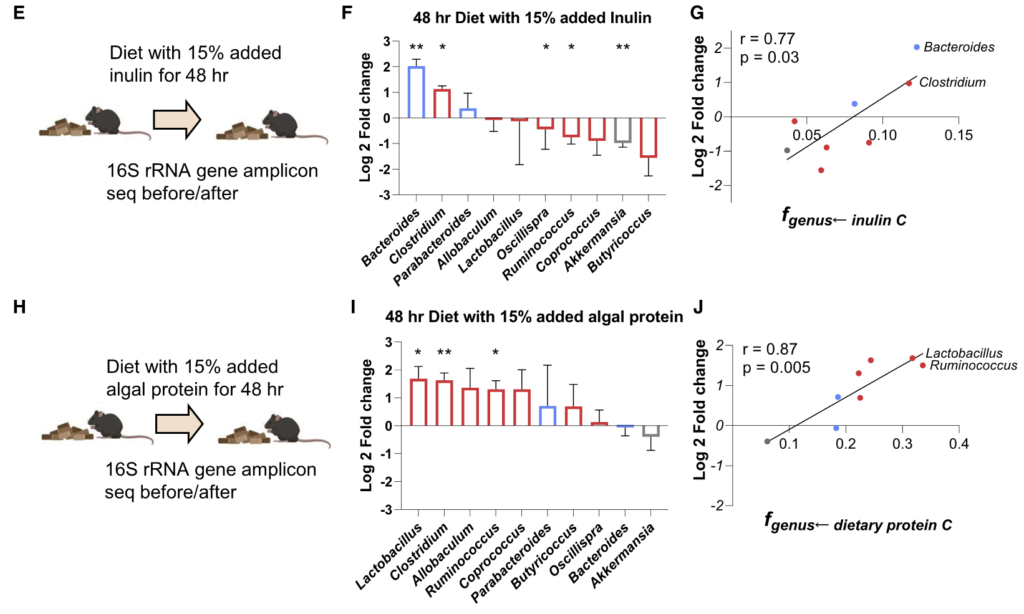
为了知晓这些细菌的营养偏好是否能预测饮食变化后的肠道菌群的组成变化。研究人员给小鼠喂食富含菊粉或藻类蛋白的饮食 2 天,并通过 16S rRNA 测序测量微生物组的组成。
结果如图F和I:
利用最多菊粉的拟杆菌属在高菊粉饮食后增加了4倍;
另一种利用较多菊粉的梭状芽胞杆菌也增加了2倍;
利用较少菊粉的菌属要么没有变化,要么略有下降;
富含藻类蛋白饮食的实验结果同理。
图G和J计算了这两种营养物与对其利用程度最高的前两名菌属相对丰度的相关性,p<0.05呈显著相关。
“不同肠道细菌的营养偏好有助于解释饮食操作后微生物组分的变化。”
最后,研究人员转向不同肠道细菌的氮源偏好,比较15N标记的膳食蛋白喂养和15N尿素输注。
高度利用膳食蛋白质中碳的细菌属也高度利用膳食蛋白中的氮,这与细菌蛋白质组中完整吸收的膳食蛋白质中的氨基酸一致。
厚壁菌喜欢从膳食蛋白质获取氮
在厚壁菌门成员中,偏好尿素氮的属往往是菊粉的疯狂使用者,即使用菊粉和尿素合成自己的氨基酸。这包括一些脲酶阴性菌属,它们可能通过交叉喂养获得尿素氮。
此外,在厚壁菌中也看到了一些属更喜欢从膳食蛋白质中获得氮,而其他属更喜欢循环尿素。
静脉注射尿素以提高循环尿素浓度后,偏好尿素的厚壁菌以及阿克曼菌的丰度大幅增加。
拟杆菌喜欢从宿主分泌的蛋白质中获取氮
与厚壁菌相比,拟杆菌对膳食蛋白质和循环尿素氮的利用率较低,这提出了一个关键问题:
拟杆菌如何获得氮?
肠道微生物群的一些成员(如拟杆菌和阿克曼菌)能够消化宿主分泌的蛋白质,如粘蛋白。
假设宿主分泌的蛋白质是拟杆菌氮的关键来源。为了探索这种可能性,研究人员进行了长期15N标记的赖氨酸和精氨酸输注(12、18和36小时),以标记结肠中的宿主蛋白。
尽管没有直接给微生物组喂食,但在36小时输注后,赖氨酸和精氨酸确实起作用,这与通过宿主蛋白进行的标记一致。这种标记优先发生在拟杆菌和阿克曼菌中。
膳食和分泌宿主蛋白的氮贡献呈负相关,与某些肠道细菌优先消耗膳食蛋白和其他宿主蛋白一致。
“膳食蛋白质和循环尿素是厚壁菌的主要氮原料,而分泌的宿主蛋白质为拟杆菌提供氮。”
研究人员开发了定量同位素追踪方法来测量肠道细菌的营养偏好。除了膳食纤维和分泌的宿主蛋白外,还将膳食蛋白和循环宿主乳酸、3-羟基丁酸和尿素确定为喂养肠道细菌的重要营养素。排除了其他循环宿主营养素(如葡萄糖和氨基酸)对结肠微生物群的直接贡献。
一项关键技术成就是能够从不同碳源和氮源追踪到细菌特异性肽,从而揭示复杂和竞争性肠腔环境中不同细菌的营养偏好。
厚壁菌门倾向于从膳食蛋白质获得氨基酸,而拟杆菌门更多地依赖宿主分泌蛋白。同样,一些厚壁菌门(如梭菌属)大量利用纤维(菊粉),而其他厚壁菌门则不利用纤维。
动物饮食干预实验发现,拟杆菌属和梭菌属是转化纤维最活跃的菌属。宿主循环代谢物水平也可能影响微生物组的营养获取和最终组成。
本文提供了关于哪些营养素喂养肠道微生物群以及哪些细菌更喜欢哪些营养素的基础知识。
文中所开发的方法具有广泛的应用前景,最终将有助于全面和定量地了解饮食-微生物-健康的关系。
参考文献:Zeng X, Xing X, Gupta M, Keber FC, Lopez JG, Lee YJ, Roichman A, Wang L, Neinast MD, Donia MS, Wühr M, Jang C, Rabinowitz JD. Gut bacterial nutrient preferences quantified in vivo. Cell. 2022 Sep 1;185(18):3441-3456.e19. doi: 10.1016/j.cell.2022.07.020. PMID: 36055202; PMCID: PMC9450212.

谷禾健康


写在前面
《基于肠道菌群思考饮食》
所谓“病从口入”,饮食是人类健康的核心。
有没有发现这样一种现象?我们的日常饮食正在向西式饮食过渡,与此同时,各类慢性病正日渐侵蚀。
近期在《CELL》子刊上接连发表了好几篇关于肠道菌群和健康饮食以及疾病的发生等循证科研研究和综述,谷禾长期的肠道菌群检测实践中也发现各种干预措施和用药治疗均需要考虑饮食营养的部分,结合肠道菌群的干预将起到更有效更长期的效果,例如炎症性肠病患者干预治疗期间,需要结合营养调整或补充来重塑肠道菌群和免疫平衡。
一系列严格的前瞻性队列研究和随机对照试验为健康饮食的特征提供了有力的验证:
蔬菜和水果、全谷物带来益处,加工食品带来有害影响。
然而,仍然存在许多争议,大家可能对“健康饮食”这个概念停留在模糊层面,依然存在许多困惑例如:


本文我们试着从微生物组科学的角度,讨论基于食物的饮食指南和健康饮食的各个方面,尽可能将我们已了解的科学知识融入其中,当然讨论仅限于针对普通人群的健康促进和疾病预防建议,这也是基于肠道菌群和食物营养的饮食推荐的目的。
以食物为基础的饮食指南提供了关于食物、食物组分和饮食模式的建议,以实现营养素的参考摄入量,预防慢性病,并维持普通人群的整体健康。
下表显示了过去10年中更新的国外基于食品的饮食指南的关键信息的非详尽概述,这些指南分为最常见和补充食品组:蔬菜和水果、谷物制品、乳制品、肉类和肉类替代品和加工食品。
非详尽的国家以食物为基础的膳食指南清单和一般成年人的食物组推荐摄入量



doi.org/10.1016/j.chom.2022.04.016
2022年4月26日,《中国居民膳食指南(2022)》发布,提出了平衡膳食八准则:
不同的饮食文化国家的指导原则,有很强的一致性,所有指南都在告诉我们:
建议限制或避免添加高糖、高盐和高饱和脂肪的食品,一些国家特别提到避免加工、超加工和/或包装食品。
一半以上的饮食应包括蔬菜、水果和谷物产品,全谷物优先于精制谷物。这些饮食需要补充少量的动物性蛋白质(鱼类、瘦肉、家禽、鸡蛋和乳制品)和/或植物性蛋白质(豆类、坚果)。
下面,我们在微生物组的背景下讨论饮食指南,重点是人类研究,尽管存在局限性(例如,大剂量给药和向人类转化),但动物模型在建立分子机制和因果关系方面存在优势。
在所有饮食指南中:
✔建议食用蔬菜、水果、全谷类、豆类和坚果等全植物食品
✖ 应限制摄入添加糖、盐或饱和脂肪的加工食品
膳食纤维
全植物食品是膳食纤维的唯一相关天然来源,膳食纤维是一种难以消化的碳水化合物聚合物,在化学成分、物理化学性质和生理效应方面有很大的不同。
与肠道微生物群特别相关的是可发酵纤维,也称为微生物群可获得碳水化合物(MAC),如果它们显示出与对微生物群的选择性影响相关的既定健康益处,则为益生元。
纤维为抑制粘液聚糖代谢的微生物提供生长基质,防止肠道粘液耗尽、细菌侵入粘液层、下游炎症和感染。
纤维发酵的主要最终产物是短链脂肪酸 (SCFA),即乙酸盐、丙酸盐和丁酸盐,它们会引起多种生理效应。
这些影响包括:
高纤维饮食可降低心脏病发作、中风和心血管疾病的风险。
近期,一项纵向、随机、交叉设计主要研究两种流行的纯化纤维,阿拉伯木聚糖 (AX) 和长链菊粉 (LCI),结果发表在《CELL Host & Microbe》.
该研究显示了单个纯化纤维对微生物组的影响,深入了解了纤维补充剂的影响以及纤维诱导的胆固醇降低背后的机制。
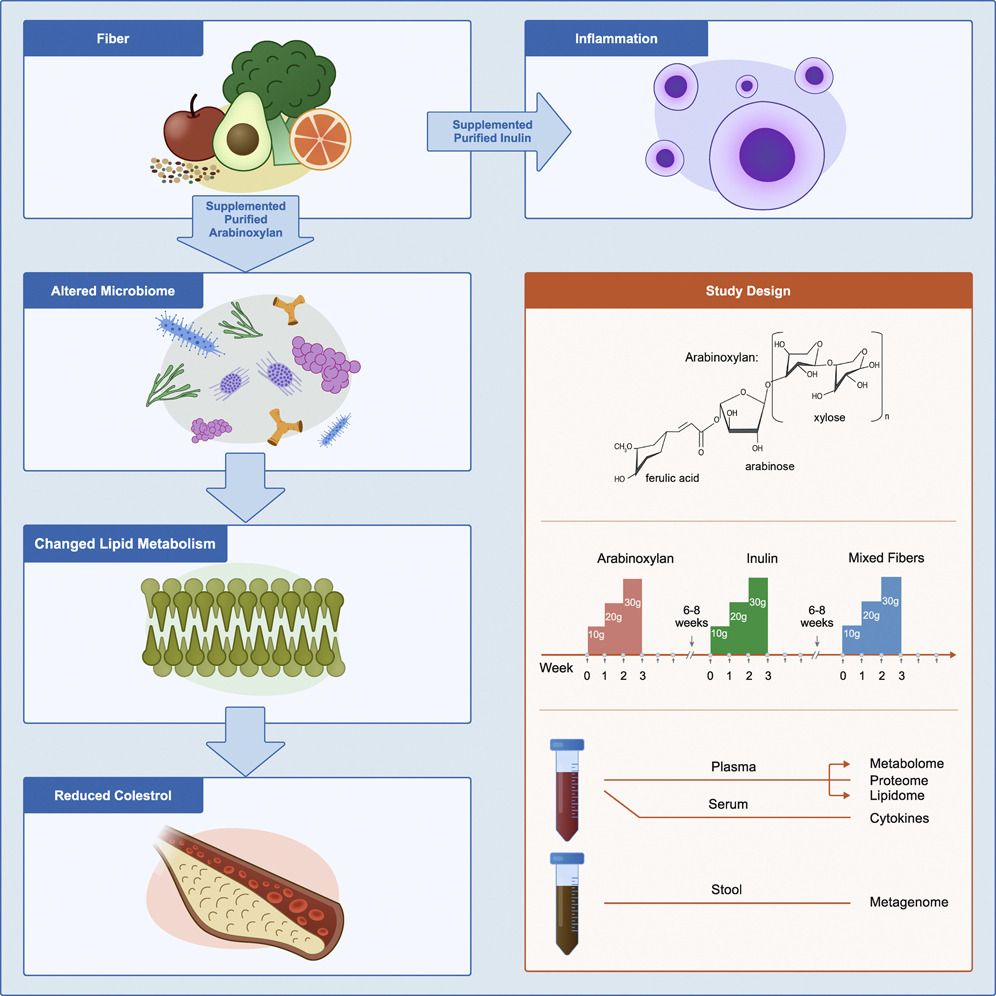
doi.org/10.1016/j.chom.2022.03.036
结果表明 ↓↓↓
阿拉伯木聚糖:
长链菊粉:
总的来说,每一种纤维都可引起个体化和纤维特异性的生化和微生物群反应,因此,膳食纤维对健康的益处因人而异,可能还取决于具体摄入的纤维类型和摄入剂量。
植物化学物质
全植物食品的另一个关键成分是植物化学物质,它们是非营养和生物活性化合物,通常与膳食纤维结合,赋予植物颜色、风味、气味和涩味。
大多数 (90%–95%) 的植物化学物质不会被小肠吸收,因此会与肠道微生物群进行双向相互作用。
肠道微生物负责通过去甲基化、开环和脱羟基等过程对植物化学物质进行生物转化,从而增加它们的生物利用度、吸收以及抗氧化和免疫调节作用。
全植物食品对肠道微生物群的影响及对宿主生理、免疫、代谢和疾病风险的影响
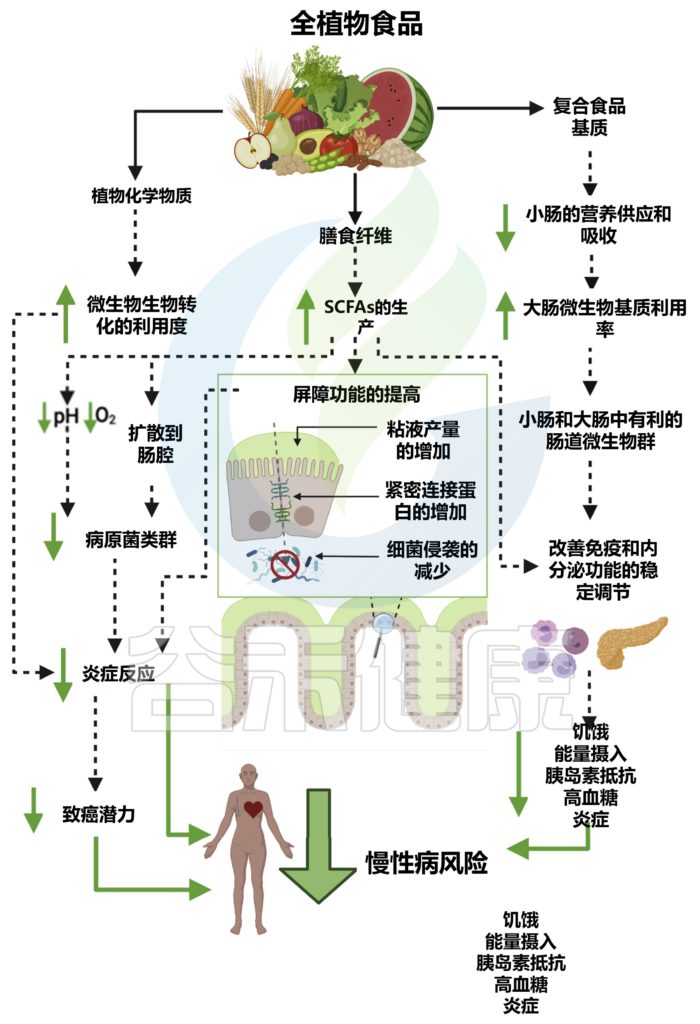
doi.org/10.1016/j.chom.2022.04.016
尽管全植物食品具有有益的效果,但工业化国家的全植物食品消费量始终低于建议的水平,而包装食品经过不同程度的加工。
根据NOVA食品分类工具,食品加工程度是饮食质量的主要驱动因素,加工程度最高的食品被归类为“超加工食品”。其有害影响(增加能量摄入和体重增加)已在严格控制喂养研究中得到实验验证。然而,超加工食品的定义或此类食品的示例几乎没有一致性,关于避免所有超加工食品而不考虑其个别营养属性的全面建议存在争议。
加工食品如何带来不利影响?
加工食品的功能特性与全植物食品有着根本的不同。加工食品通常具有较高的能量密度,并且缺乏植物细胞中存在的三维结构。因此,营养素主要是无细胞的(即不包含在细胞内),更容易被宿主消化,这增加了营养素吸收动力学。
这些容易发酵的营养素可能会促使小肠中的细菌过度生长,以及不利的微生物组成和代谢状况,从而对免疫和内分泌功能产生负面影响,而结肠微生物群无法获得这些营养素。
例如,高果糖玉米糖浆已被证明会导致小鼠脂肪肝和葡萄糖不耐受,其方式与肠道微生物群组成和功能变化有关。
加工食品:食品添加剂影响菌群
加工食品中的食品添加剂,可以进一步提高口感和保质期,但是会影响肠道微生物群。
合成乳化剂羧甲基纤维素(CMC)和聚山梨酯-80损害肠道屏障功能,导致微生物群侵占上皮,促进野生型小鼠的代谢异常和轻度炎症,以及遗传易感小鼠的结肠炎,其方式与肠道微生物群有因果关系。
在另一种乳化剂甘油单月桂酸盐的小鼠中也观察到类似的结果。人类短期食用CMC也会显著改变微生物群组成,降低粪便短链脂肪酸水平,并诱导细菌侵入粘液层。
加工食品中的高盐含量也可能改变微生物群。盐的摄入降低了乳酸菌的丰度,这与小鼠小肠固有层淋巴细胞和人类外周血淋巴细胞中辅助性T细胞17的数量增加以及血压升高有关。
另一项针对小鼠的研究报告了类似的结果,其中高盐饮食降低了乳酸菌的丰度,增加了促炎基因的表达,并加剧了两种不同疾病模型中的结肠炎。
因此,现有证据表明,加工食品和全植物食品对人类健康的对比效应部分是由肠道微生物组介导的。
加工食品对肠道微生物群的影响及对宿主生理、免疫、代谢和疾病风险的影响
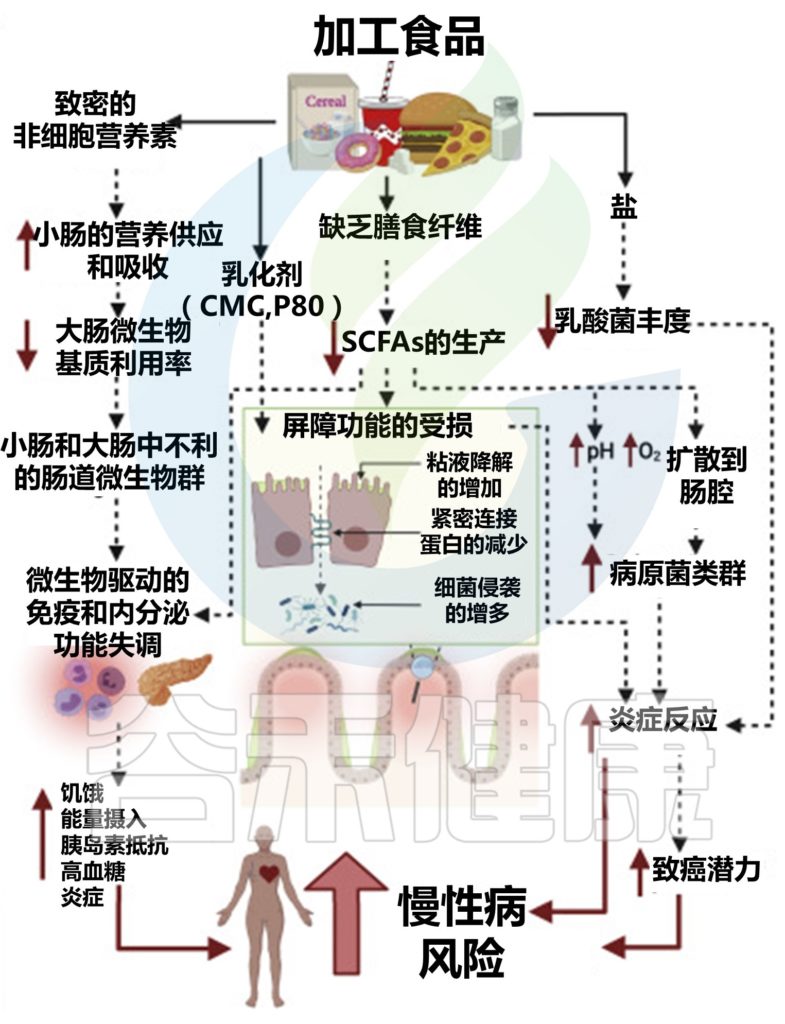
doi.org/10.1016/j.chom.2022.04.016
更多关于食品添加剂与菌群详见之前的文章:
你的焦虑可能与食品添加剂有关,警惕食品添加剂引起的微生物群变化
根据饮食指南,健康饮食的最大组成部分应该是蔬菜和水果。目前关于蔬菜水果预防多种慢性病的能力的科学证据十分有力。
▸蔬菜水果富含膳食纤维
水果和蔬菜每份可提供高达 8g 的膳食纤维,并含有多种纤维,包括果胶、菊糖、纤维素、木聚糖、棉子糖和水苏糖。这些纤维引发微生物组独立(例如延迟的大量营养素吸收)和微生物组依赖(例如短链脂肪酸介导的胰岛素抵抗衰减)生理效应。
人类对照喂养实验发现,富含菊糖的蔬菜可提高双歧杆菌水平,促进饱腹感,并减轻体重。
在喂食高脂肪饮食的小鼠中,抗生素治疗降低了菊糖的代谢益处,比如降低这些益处:在小肠中诱导肠促胰岛素激素胰高血糖素样肽1和对代谢综合征的保护,这表明微生物群的因果作用。
▸蔬菜和水果也是植物化学物质的重要来源
包括多酚、硫代葡萄糖苷、萜类、植物甾醇和生物碱。
蔓越莓提取物是一种多酚类物质的丰富来源,在高脂高糖饮食的小鼠中诱导代谢改善(例如,减少内脏肥胖和改善胰岛素敏感性),这与 Akkermansia muciniphila的增加有关。
注:Akkermansia muciniphila菌在动物和人类中都显示出有益生理效应。关于该菌详见之前文章:
肠道重要菌属——Akkermansia Muciniphila,它如何保护肠道健康
Nature | AKK菌——下一代有益菌
从临床研究和临床前研究中,也有新的证据表明微生物组在西兰花的生理效应中发挥作用。后者为西兰花衍生的硫代葡萄糖苷生物活性转化为化学预防性异硫氰酸酯奠定了拟杆菌的遗传和生化基础。
总的来说,现有证据支持肠道微生物群在调解蔬菜和水果对健康的影响方面的作用。
食物对宿主-微生物相互作用的影响以及它们如何与膳食指南中的建议保持一致

doi.org/10.1016/j.chom.2022.04.016
扩展阅读:常见水果对肠道菌群、肠道蠕动和便秘的影响
大多数饮食指南建议食用全谷物而非精制谷物。全谷物的麸皮层被去除以生产精制谷物,含有植物化学物质(如阿魏酸、类黄酮)和膳食纤维。
▸ 全谷物含有独特纤维,抗炎作用与菌群相关
全谷物含有独特的半纤维素纤维,如木聚糖和β-(1→ 3,1 → 4)—除纤维素、抗性淀粉和低聚糖外,还有葡聚糖。全谷物能够降低慢性病风险的证据基础是令人信服的。人类干预试验表明,全谷物的抗炎作用与肠道微生物群的变化是并行的。
在健康成年人中,显示了对健康有益微生物的富集,如双歧杆菌和产丁酸菌(直肠真杆菌Eubacterium rectale、Roseburia faecis, Roseburia intestinalis),产短链脂肪酸菌Lachnospira增多,粪便乙酸盐和总短链脂肪酸增加,肠杆菌科减少。
虽然微生物组的变化为抗炎作用提供了潜在的解释,但这些研究之间并不一致;进一步的研究报告没有任何影响,也没有确定因果关系。
▸ 全谷物的部分代谢益处是由肠道菌群介导的
将人类研究与小鼠实验相结合的研究表明,微生物组对全谷物的健康影响具有因果关系。
食用大麦仁面包的人被分为“响应者”和“无响应者”,“响应者”的葡萄糖代谢因干预而得到改善。
响应者普氏菌/拟杆菌属比率和Prevotella copri丰度升高,以及编码复杂多糖降解的微生物基因在应答者中升高。
关于Prevotella copri菌详见:肠道重要基石菌属——普雷沃氏菌属 Prevotella
无菌小鼠灌胃普氏杆菌或含有P.copri的“应答者”人源性微生物群后,喂食标准食物后,葡萄糖耐量有所改善,这在机制上与肝糖原储存增加有关。
进一步的研究证实,在基线时含有高水平的普雷沃氏菌,会导致体重过重,且食用富含全谷物的饮食的个体的体重减轻。
一些饮食指南(加拿大的食品指南、巴西人口的饮食指南和英国的Eatwell指南)建议经常食用植物蛋白食品(如豆类、坚果),因为它们对人类健康有益。
豆类
豆类富含纤维,特别是纤维素、果胶、甘露聚糖、水苏糖、棉子糖和抗性淀粉。
豆科植物还含有黄酮醇等植物化学物质,黄酮醇是一种已知的抗炎类黄酮亚类,还含有酚酸,与谷物中的酚酸相比,豆类中酚酸的生物利用率更高。
新出现的证据表明,肠道微生物群在豆科植物的健康影响中发挥着作用。
例如,绿豆补充剂减少了喂食高脂肪饮食的小鼠的体重增加和脂肪积累,但没有减少喂食相同饮食的无菌小鼠的体重增加和脂肪积累。
坚果
坚果是不饱和脂肪酸、纤维和植物化学物质的来源,所有这些都可能影响宿主与微生物的相互作用。
在一项控制喂养研究中,每天补充核桃可增加粪杆菌属(Faecalibacterium)、罗氏菌属(Roseburia)、梭菌(Clostridium)和戴阿利斯特菌(Dialister)。
在一项杏仁对照喂养研究中观察到了类似的成分变化,显示罗氏菌属(Roseburia)、梭菌(Clostridium)、戴阿利斯特菌(Dialister)和毛螺菌属(Lachnospira)的相对丰度增加。
坚果对罗氏菌属丰度的影响也已在meta分析中得到证实。
主要产丁酸菌如Roseburia intestinalis能够将坚果中难以消化的聚糖β-甘露聚糖代谢为丁酸盐。
坚果中的omega-3脂肪酸也可能增强罗氏菌属,因为在人类中,补充核桃的饮食和不含核桃的具有相同脂肪酸组成的饮食都增加了罗氏菌属的相对丰度。
除了坚果中的纤维和不饱和脂肪酸外,肠道微生物还将植物化学物质鞣花单宁和鞣花酸代谢为尿石素,尿石素是一种生物活性抗炎化合物。
总之,增加坚果摄入量似乎在一定程度上通过影响微生物群的功能成分对宿主健康有益。
抗炎看到目前研究将植物性蛋白质食品的益处与微生物组联系起来,但该领域的研究仍处于初步阶段。
植物性蛋白质比动物性蛋白质更不易消化并因此为结肠微生物提供潜在的底物,这可能导致产生有益的生物活性代谢物,例如色氨酸代谢。然而,蛋白水解微生物发酵也可能有害(见下文讨论——红肉和加工肉)。目前尚不清楚植物蛋白微生物发酵产生的代谢结果是否不同于动物蛋白,需要进一步研究。
饮食指南一直鼓励把鱼类作为优质蛋白质来源和良好的脂肪酸组成。
多脂鱼类
多脂鱼类是长链ω-3脂肪酸二十碳五烯酸(EPA)和二十二碳六烯酸(DHA)的主要天然膳食来源之一。
观察研究和干预研究的有力证据表明,多脂鱼摄入量的增加具有心脏保护作用,肠道微生物群可能是这些健康影响的潜在中介。
鱼油
小鼠实验表明,与猪油(富含饱和脂肪)相比,鱼油降低了 Toll 样受体激活和白色脂肪组织炎症,这与改善胰岛素敏感性有关。
除了这两种脂肪对微生物群组成的不同影响(猪油增加Bilophila菌)之外,与传统猪油喂养的小鼠相比,喂食鱼油的无菌小鼠和喂食抗生素的小鼠的盲肠微生物群定植后,体重减轻,白色脂肪组织炎症减轻。
这项研究为肠道微生物群在饱和脂肪的炎症作用中的因果作用提供了证据。
尚未在人类身上做出因果推论,但在多脂鱼和其他饮食来源中发现的 omega-3 脂肪酸已被提议作为候选益生元,因为它们被特定的肠道微生物利用。
在一项随机对照试验中,鱼源性 omega-3 补充剂增加了Coprococcus的丰度,与富含甘油三酯的脂蛋白水平呈负相关。
因此,肠道微生物组可能是多不饱和脂肪酸心脏保护作用的介质。
以上各种饮食推荐都接近一种饮食模式,那就是地中海饮食,作为现在十分流行的饮食模式之一,
地中海饮食有什么独特之处?
对菌群有哪些影响?
接下来详细了解一下地中海饮食。
我们逐渐开始有这样一个认识,健康不止是受个别食物的影响,而是受其相互联系和协同效应的影响。因此,最近更新的几项饮食指南,如《中国饮食指南2022》、《2020-2025年美国人饮食指南》、《加拿大食品指南》等对饮食模式的关注发生了转变。
食物之间的相互作用也与它们对肠道微生物群的影响有关,人们对饮食模式,尤其是那些具有既定健康益处的饮食模式(如地中海饮食)如何通过微生物群组成和功能变化影响宿主健康感兴趣。
▸ 地中海饮食结合了许多对宿主微生物相互作用具有有利影响的食物组
地中海饮食推荐蔬菜、水果、全谷类、豆类、坚果和橄榄油作为主食,适度摄入鱼类、家禽、鸡蛋和乳制品,并限制摄入红色和加工肉类及加工食品。
在对体重超标的个体进行的一项随机对照试验中,与对照饮食相比,坚持地中海饮食可降低血浆胆固醇浓度,并增加了产丁酸菌普拉梭菌(F. prausnitzii)和罗氏菌属(Roseburia) 的丰度。F.prausnitzii和Roseburia也是通过随机森林模型确定的分类群,这些模型可以最好地预测饮食依从性得分。
▸ 地中海饮食改善老年人认知
在一项大型多中心随机对照试验中,该试验涉及了连续12个月食用地中海饮食的老年人。这些分类群与认知功能的改善呈正相关,与炎症标志物和虚弱呈负相关。
▸ 地中海饮食降低心肌梗死风险
在一项前瞻性队列研究中,长期坚持地中海饮食与F. prausnitzii, Eubacterium eligens, Bacteroides cellulosilyticus 呈正相关。这项研究进一步表明,与那些携带P.copri的人相比,P.copri丰度较低且坚持地中海饮食的人心肌梗死风险较低。
对地中海饮食的微生物组学研究强调了其在饮食建议中的重要性。最近部分国家更新的饮食指南推荐了类似地中海饮食的饮食模式,如阻止高血压的饮食方法(DASH)饮食,并对微生物组在调节其健康影响中的作用进行了研究。
虽然饮食指南已经演变为反映新的可用证据,并普遍同意其建议,但争议仍然存在。
在这里,我们讨论了宿主-微生物相互作用如何提供洞察力来帮助解决这些争议。
红肉含有必需的微量营养素,是重要的优质蛋白质来源。然而,大多数饮食指南建议只适度食用,同时建议避免食用加工肉类(盐腌、腌制和烟熏的肉类)(表1)。
▸ 红肉和加工肉类引争议?
在癌症预防方面,这些评估由国际癌症研究机构(IARC)和世界癌症研究基金会共同进行。尽管有这些一致的建议,但2019年的一系列系统性审查得出结论,成年人应该继续目前的红肉和加工肉摄入量,理由是它们与不良健康结果的联系的证据不足。这引发了营养领域的争议。
肠道菌群为这场争论提供了另一个视角
肠道微生物对红肉和加工肉类的蛋白水解发酵会导致产生潜在的有害代谢物,例如氨、对甲酚和硫化氢。
▸ 红肉和加工肉→产硫化物菌发酵→可能致癌
硫化氢是由脱硫弧菌等细菌发酵含硫氨基酸产生,并作为粘液溶解剂,可以增加小鼠肠道通透性。
这使得同样存在于红肉和加工肉类中的血红素能够增加肠道中的过度增殖和增生,抑制肿瘤抑制基因,并激活致癌基因,所有这些都是致癌的潜在原因。
抗生素给药抑制了小鼠的这些影响,表明肠道微生物群的因果作用。在人类中,产生硫化物的细菌和途径与晚期结直肠癌密切相关。
▸ 红肉和加工肉→TMAO→心血管疾病
肠道微生物将肉制品中大量存在的左旋肉碱和磷脂酰胆碱转化为三甲胺,三甲胺被含肝黄素的单加氧酶 (FMO) 氧化为三甲胺-N-氧化物 (TMAO)。
TMAO 在血浆中循环并由肾脏有效排泄,并在动物模型中与心血管疾病有因果关系。观察性研究报告了 TMAO 水平与心血管疾病风险之间的强正相关关系。然而,动物实验并不一致,而人类中的 TMAO 水平经常被混淆并可能由肾功能下降引起。
此外,TMAO 范式与将富含 TMAO 及其前体的脂肪鱼与有益的心脏代谢结果联系起来的流行病学发现不一致。
肉类以外的食物(即十字花科蔬菜)可以抑制 FMO3 的活性,使研究结果更加复杂。
为了阐明源自肉类产品的 TMAO 的因果贡献,需要在人类中进行足够持续时间的控制喂养试验,以评估经过验证的心血管疾病的替代标志物。
▸ 加工肉类含有瘦红肉中不存在的其他化合物,这些化合物可能会放大微生物介导的有害影响
大多数加工肉类富含饱和脂肪,可刺激小肠中的肝胆汁酸分泌。
一些初级胆汁酸逃脱肠肝再循环并进入大肠,在那里它们被微生物转化为次级胆汁酸。其中,脱氧胆酸和石胆酸可引起氧化应激和 DNA 损伤,并与结肠癌发生有关。
一项荟萃分析发现,在结直肠癌患者中,肠道微生物组功能特征的特征是次级胆汁酸的产生增加。而一项系统评价表明饱和脂肪会降低微生物的丰富度和多样性。
此外,加工肉类中使用的固化剂硝酸盐和亚硝酸盐构成微生物生物转化为N-亚硝基化合物的底物,它们会导致 DNA 烷基化损伤,因此具有致癌性。
▸ “适度食用瘦肉红肉和避免加工肉” 的科学依据
鉴于上述几点,红肉和加工肉类产生的肠道微生物代谢物的毒性与解释它们对健康的影响有关。
尽管有证据表明红肉是结肠癌的危险因素,但蛋白质发酵产生的代谢物(如硫化氢和氨)毒性较低,目前还没有归类为人类致癌物。
相比之下,仅来自加工肉类的微生物代谢产物(N-亚硝基化合物和次生胆汁酸)具有更高的毒性和致癌性。
考虑到可能的剂量-反应关系,毒理学考虑证明了IARC/世界卫生组织专家小组对红肉(2A组,可能致癌物)和加工肉(1组,致癌物)进行风险分类的合理性,以及当前的饮食指南:适度食用瘦肉红肉和避免加工肉。
▸ 长期存在争议:乳制品应在多大程度上纳入健康饮食?
大多数饮食指南建议使用脱脂和低脂(0%-2%)乳制品,并避免使用高脂乳制品(>25%,如某些奶酪、奶油制品和黄油)。然而,对于全脂乳制品还没有达成共识(∼3.5%),其有害影响受到质疑,一些饮食指南(表1)不鼓励使用。
▸ 乳脂肪和肠道微生物群之间的相互作用
一项开创性的研究表明,乳源性饱和脂肪诱导牛磺酸结合胆汁酸,促进产生硫化氢的致病菌Bilophila wadsworthia的爆发,从而引发遗传易感小鼠的结肠炎。
另一项针对小鼠的研究证实了这些发现,其中主要来源于乳脂的高脂肪饮食增加了B. wadsworthia的丰度和盲肠胆汁酸水平,从而导致肠道屏障功能障碍和代谢综合征。
这些动物模型强调了乳源性饱和脂肪对微生物群动态平衡的潜在有害影响,支持了建议限制高脂肪乳制品的饮食指南。
据我们所知,目前缺乏评估全脂乳制品中饱和脂肪水平是否影响微生物群的良好控制的人类干预试验,此类研究有必要,能为未来的饮食指南提供信息。
“低脂”、“低碳”,这对减肥人士来说,是非常熟悉的。
目前的膳食指南没有涉及到限制脂肪或碳水化合物摄入是否会支持最佳健康。
▸ 这些饮食如何影响微生物群代谢以及对健康的长期影响?
低脂饮食通常富含蔬菜、水果、全谷物和植物蛋白质,因此提供有益的饮食成分,改变微生物代谢。
相反,低碳水化合物饮食通常脂肪和/或蛋白质含量较高,因此纤维含量较低,这导致产生对结肠健康有害的代谢物。
这在一项随机对照试验中得到证实,在该试验中,高蛋白低碳水化合物饮食增加了N-亚硝基化合物的浓度,降低了丁酸盐和抗炎酚类化合物的水平。
在另一项研究中,高脂肪、低碳水化合物饮食导致健康年轻人的肠道微生物群、粪便代谢组学特征和血浆促炎介质发生不利变化。
这些发现引起了人们对低碳水化合物饮食的长期健康结果的担忧,并与前瞻性队列研究的荟萃分析一致,该研究显示,食用动物性低碳水化合物饮食的参与者死亡率最高。
因此,有必要通过针对微生物组的方法改善低碳水化合物饮食。
当代国家饮食指南高度一致,与研究人员对饮食如何影响与健康相关的微生物-宿主相互作用的理解一致。
这种一致性是显著的,证实饮食指南的观察性和干预性研究没有考虑微生物组。这表明肠道微生物群是饮食生理效应的关键中介,鉴于哺乳动物解剖学、生理学、免疫和代谢的中心方面在整个进化过程中都是由饮食-微生物-宿主相互作用形成的,这一功能可能在进化上根深蒂固。
虽然营养学和微生物组学学科之间的一致性在很大程度上验证了当前的饮食指南,但研究人员认为,更系统地结合营养素影响宿主-微生物相互作用的分子基础知识,有可能加强和创新人类营养。
下面概述了微生物组学观点可以推进营养策略的机会,然后提出了一个整合肠道微生物组学的研究框架,以进行实验验证。
对人类及其肠道微生物群共同进化的假设最有说服力的支持来自对母乳低聚糖(HMO)功能特征的解释。母乳构成了饮食、微生物组和人类健康之间进化关系重要性的范例。
现代饮食可能在进化上与人类生理不匹配,这可能是慢性病流行的重要驱动因素。
目前建议女性和男性每天摄入 25 克和 38 克纤维。一系列系统评价和荟萃分析支持了更高摄入量的论点,表明每天摄入超过25–29克的纤维将带来额外的益处。参与者每天从蔬菜、水果和坚果饮食中摄入100克以上的纤维,可以显著降低血清胆固醇水平,增加粪便中的短链脂肪酸。
在一项人体试验中,非裔美国人和南非农村人(分别习惯食用低纤维和高纤维饮食)交换了他们的饮食,导致非裔美国人的黏膜增殖率和结肠炎症(结肠癌风险的生物标志物)下降,而南非农村人在这些试验中经历了不利的变化。
饮食交换的效果与次级胆汁酸丰度和短链脂肪酸生成的相反变化相关。对饮食微生物群相互作用如何影响人类生理学的进化考虑,可以为饮食建议、有针对性的营养策略以及对抗慢性病风险的食品开发提供信息。进化的考虑也为微生物组的恢复策略奠定了基础。
工业化造成了非传染性慢性病的增加和微生物组的枯竭,其特征是微生物多样性减少,利用碳水化合物的酶的能力降低,发酵减少,以及黏液降解生物的富集。
尽管将微生物群恢复到祖先的状态可能是不现实的,也可能是不可取的,但人们对开发微生物群恢复策略、重建与健康相关的功能特征的兴趣越来越大。
全植物食物
这种策略得到了一项人类干预研究结果的支持,该研究测试了富含全植物食物(每天提供45克纤维)的饮食,这会提高短链脂肪酸的产量,并增加聚糖降解碳水化合物活性酶的相对丰度。
仅关注膳食纤维的微生物群落恢复方法不太可能补充丢失的微生物物种。已经提出了重新引入因工业化而丢失的菌群的建议。虽然从长远来看这是一种很有希望的方法,但许多消失物种的致病性未知,这使得它们转化为营养策略具有挑战性。
发酵食品
另一种方法是发酵食品,定义为通过理想的微生物生长和膳食成分的酶转化生产的食品,其结果通常具有增强的营养特性。
发酵食品的例子有开菲尔、酸奶、康普茶、豆豉、泡菜等。发酵食品在当前的饮食趋势中排名很靠前,在全球范围内广受欢迎。如果生吃,发酵食品通常含有大量的活微生物,这些微生物具有长期的安全食用历史。
▸ 关于发酵食品健康益处的研究
发酵食品的益处已通过一项定性系统综述进行了总结,该综述对观察性和实验性研究进行了评估,报告称酸奶、开菲尔和其他发酵牛奶与胃肠道健康、2型糖尿病和癌症风险以及体重管理的良好结果相关。
此外,一项针对120000多名参与者的大型观察性研究发现,体重增加与酸奶摄入量之间存在显著的负相关关系。然而,来自随机对照试验的证据非常稀少,发酵食品才刚刚开始被推荐到饮食指南中。
一项随机对照试验对每天包括6份发酵食品的饮食进行了试验,发现增加了微生物组的多样性,并减少了几种促炎细胞因子和趋化因子。
需要额外的设计良好的随机对照试验,并验证替代终点,以证明将发酵食品纳入饮食建议的合理性。此类研究应考虑到一些发酵食品(如发酵香肠、一些奶酪和加糖酸奶)的有害营养方面,如大量的盐、饱和脂肪、糖和固化剂,可能会超过活微生物的潜在益处。
益生菌、合生元
理论上,也可以通过饮食益生菌和合生元(益生菌和益生元的组合)来实现微生物群落的恢复策略。
有大量研究在许多临床环境中探索这些策略,并提出了不同的机制,例如其免疫调节作用。如果使用人体胃肠道固有的细菌菌株,益生菌可以成功地长时间定植。
这种策略可以使微生物群多样化,但据了解,这一点尚未得到系统的测试。
也有人提议探索每日摄入活微生物的饮食建议,以促进健康。为了使这一概念可行,需要进行流行病学研究和随机对照试验,以测试益生菌在预防慢性病方面的价值。
为了提高全人口的饮食质量,建议重新配方而不是取消加工食品。
这样的尝试将需要食品工程的创新,这将受益于饮食-微生物-宿主相互作用的考虑。例如,可以用不可消化的可发酵淀粉和其他纤维部分替代食品中的白面粉,从而改变加工食品的固有特性(如纤维含量、血糖生成指数和营养物质消化率),以抵消对肠道菌群和宿主代谢的有害影响。
在一项统计模型研究中,这一方法已具有广泛的影响,该研究预测,如果英国50%的加工食品的市场份额中添加约3克纤维,2型糖尿病和心血管疾病的风险将降低70%以上。
类似的方法可以应用于重新引入其他生物活性化合物,如植物化学物质,可能与膳食纤维联动。
与其依靠个人改变他们的饮食习惯改善健康,不如用更多的重新配方加工食品来改善饮食质量,而不显著改变饮食习惯。
确定“健康微生物群”
虽然很难定义什么是“健康”微生物群,但肠道微生物群的特定分类群和功能特征,特别是受饮食影响的分类群和功能特征(例如,与健康相关的SCFA和次生胆汁酸)与健康结果有关。
一旦确定了促进健康的分类群和微生物群特征,就可以用营养策略来针对它们。
例如,代谢有害的低碳水化合物或高肉类饮食,可通过补充可发酵纤维,以将微生物代谢从蛋白质转化为碳水化合物发酵,改善肠道屏障完整性,并通过短链脂肪酸诱导全身代谢益处。鉴于膳食纤维和低碳水化合物饮食的独立代谢效应,它们的结合可能产生协同效应。
扩展阅读:健康的人类微生物组
使用膳食纤维增强假定的健康菌群
通过这种方式促进菌群及其代谢产物。这种方法基本上符合益生元的概念,益生元被定义为宿主微生物选择性利用的底物,具有健康益处。
不幸的是,这一定义并没有就什么构成“选择性”效应、这种效应如何与健康益处因果关联以及如何区分益生元和膳食纤维提供明确的指导。这些担忧导致欧洲食品安全局规定,益生元不能标记为益生元,但必须标记为膳食纤维。
饮食指南中也没有提到食物来源的益生元,这很不幸,因为目标微生物组调节的总体概念很有希望。
▸ 用膳食纤维获得可预测的微生物群组成变化
研究人员提出了一个概念框架,利用该框架,可以使用含有离散结构的膳食纤维(定义为“一种独特的化学结构……与细菌基因组中编码的基因簇相一致”),来获得可预测的和理想的微生物群组成变化。
在人体试验中对该框架进行了实验测试,在该试验中,IV型抗性淀粉的细微结构差异将短链脂肪酸输出导向丙酸盐或丁酸盐,丙酸盐或丁酸盐具有不同的代谢和生理功能。
有针对性的营养微生物组调节对于饮食指南和治疗性食品都有巨大的前景,但关于微生物组的哪些方面应该有针对性的问题仍然存在,需要进行随机对照试验来证明这些策略是否能转化为改善健康的结果。
即使控制良好的饮食干预措施也会对个体产生不同的影响。这对目前在饮食指南中应用的“一刀切”方法提出了质疑。
精确营养(也称为“个性化营养”)的新兴领域旨在利用人类的个性,首先确定哪些特定的特征可以预测对饮食干预的反应,然后相应地调整营养,以实现不同人群的相同反应。
考虑到肠道微生物群对饮食的高度个性化反应,微生物群检测是针对慢性病预防和治疗的精准营养策略的关键组成部分,以及其他个人特定因素(例如,遗传学、基线代谢和体力活动)。
▸ 应用:同样的减肥方式,哪类人群更容易达到效果?——机器学习帮你预测
机器学习方法可以应用于大型参与者队列,以确定哪些特征可以预测健康结果。例如,机器学习算法利用参与者关于血液参数、饮食习惯、微生物组组成和其他因素的数据,准确预测标准化膳食的餐后血糖反应。
这类预测可以受益于微生物组和宿主遗传数据的结合,如一项研究所示,基线Prevotella/拟杆菌比率高,可以预测唾液淀粉酶基因拷贝数较低的受试者,在食用富含膳食纤维、全谷物、水果和蔬菜的饮食后体重减轻更大。
▸ 应用:微生物组结合手机用于饮食监测——机器学习为你提供高维数据,最终实现个性化建议
国家饮食指南目前没有考虑精确或个性化的方法,其实施将在人口规模上具有挑战性。然而,这样做的技术已经存在,并且可以利用微生物组测序和智能手机应用程序用于饮食监测的组合,最终为机器学习算法提供高维数据,以便将个性化营养建议反馈给用户。
▸ 精准营养可持续的条件
预测模型的多中心验证必须在不同人群(包括非西方国家和发展中国家)中进行,以确定其广泛适用性并鼓励进一步完善。精确营养方法将取决于营养和微生物学科之间的持续合作,其在全人口范围内的实施将需要监管机构、专业协会和决策者的大量额外投入。
有关饮食微生物组与宿主相互作用的信息有可能进一步验证、完善和创新饮食建议。
将肠道微生物组纳入饮食指导,需要有证据证明微生物组在饮食生理效应中的机制和因果作用。确定肠道微生物群在人类疾病易感性中的作用的因果关系仍然是一个挑战,这在营养学研究中更加复杂,因为饮食、肠道微生物群和人类健康之间的相互作用是复杂和多方向的。
饮食、肠道微生物群和人类健康之间复杂的、多向的因果关系
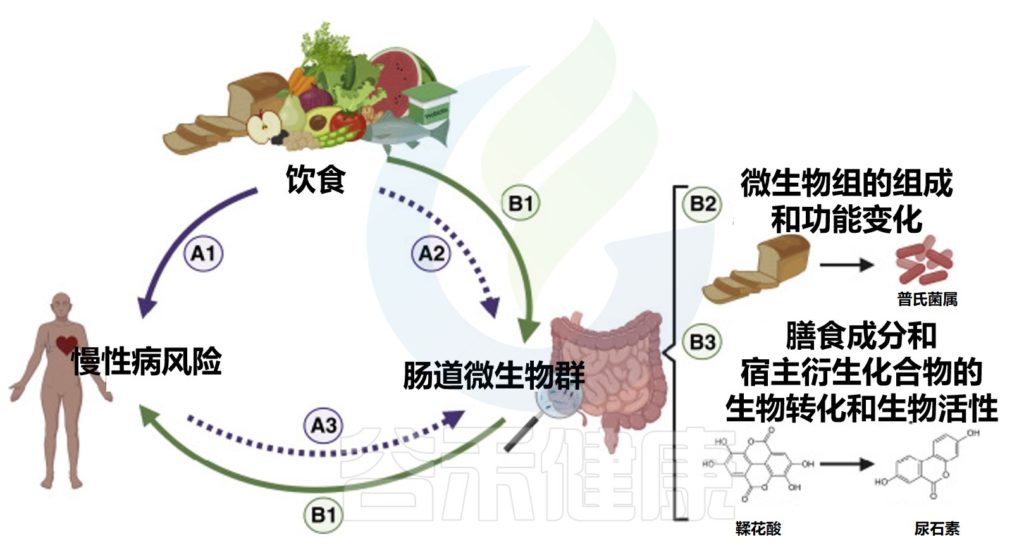
doi.org/10.1016/j.chom.2022.04.016
评估微生物组在营养中作用的人类研究因微生物组的复杂生态特征(个体间、地理和时间变异性)和营养研究的局限性(甚至适用于随机对照试验)而变得更加复杂(例如,难以评估饮食摄入和遵守研究方案、饮食成分的共线性和混杂因素)。
在设计未来的营养研究时,必须考虑这些复杂性,以阐明哪些因素(包括微生物组)介导了饮食对健康的影响。
研究人员推荐概述饮食微生物组研究最佳实践指南的优秀评论,并通过使用三大支柱将肠道微生物组整合到营养研究的所有阶段的实验框架来扩展这些评论。
一、微生物群的发现为健康饮食提出假设
肠道微生物群可以在既定饮食策略验证之外为营养提供信息,并有助于确定微生物群特征作为未来的营养目标。
目前已经确定了肠道菌群与人类疾病状态之间的关联。多元组学技术(如宏基因组学、宏蛋白质组学、代谢组学)和先进分析方法的使用,已经建立了支撑生物途径的潜在机制和因果基础。高质量、大规模队列研究有助于为饮食指南提供证据基础。
最近的研究扩展了这一框架,并将微生物组学和营养流行病学结合起来,以阐明微生物组学在饮食诱导的生理效应中的作用。可以发现微生物组、健康和特定饮食或饮食成分之间的联系。
这些相互作用的潜在机制和生物学合理性及其作为营养靶点和诊断标记的价值可在实验系统中得到证实。
二、微生物组与人类干预试验的整合
▸随机对照试验:营养学金标准
▸交叉设计:允许消除个人特定因素的个体间差异
▸干预试验:确定因果关系,直接告知饮食指南
随机对照试验是建立人类因果关系的营养学金标准,如果它们表现良好,在证据层次结构上要高于观察性试验。这种层次结构也适用于微生物研究。营养随机对照试验可以使用微生物组流行病学中应用的相同多组学方法进行扩展,以整合肠道微生物组并有效测试特定饮食微生物组的相互作用。可以将有效预测疾病风险的替代标记物与微生物组终点(例如成分变化、功能和代谢物)和生物过程的分子标记物相结合,假设这些生物过程将肠道微生物组的代谢活动与宿主免疫代谢相联系(即肠道激素、细胞因子、TMAO和肠道屏障完整性标记物),以证实研究结果并提供假定的机制解释。
交叉设计对于以微生物组为终点的随机对照试验具有优势,因为参与者作为自己的对照,这允许消除个人特定因素的个体间差异(例如微生物组、遗传学、代谢物谱和基线临床测量)。研究应进一步控制其他混杂变量,如人口统计学(年龄和性别)和生活方式因素(习惯性饮食和药物使用)。在这种情况下,大便稠度和饮酒是意外的混杂因素。应使用分层随机化来平衡参与者根据年龄和性别等因素分配给治疗组的情况,并应收集有关混杂变量的详细信息,以便在统计分析中对其进行控制。通过对自由生活或定居的参与者进行充分的控制喂养研究,提供所有食物,包括足够的磨合期,可以消除习惯性饮食的实质性混杂因素。这样的研究进行起来困难且昂贵,但已经在微生物组领域成功应用。
干预试验可以确定饮食对健康影响的因果关系,从而直接告知饮食指南。如果微生物组被整合,随机对照试验也可以为微生物组在饮食健康影响中的作用提供假定的机制解释,并为精确营养策略提供基于诊断微生物组的生物标记。回归和相关分析可用于确定饮食诱导的微生物组分/功能变化与临床和机械终点之间的关联。此外,机器学习模型可以确定是否可以通过对微生物组或受微生物组影响的宿主生物过程的影响来预测饮食诱导的生理变化。然而,除非这些试验和分析采用特定的实验和统计方法进行扩展,否则它们无法确定肠道微生物群在饮食影响中的因果作用。
三、关于微生物组在饮食影响中作用的机制洞察和因果推断
▸收集纵向数据,使用数学方法,解开因果
▸通过动物模型扩展确定潜在机制
▸最常用和最复杂的建立菌群因果关系的模型
▸动物模型存在局限性,但对完善假设很宝贵
可以在随机对照试验中收集纵向数据,以便进行因果推断,因为必须使用中介分析等方法,在时间上先于效应。其他数学方法,如结构方程建模和贝叶斯网络,也可以解开饮食、微生物组和人类健康之间的因果关系。可以说,最有希望直接在人类身上进行因果推理的实验设计是“干预主义框架”,当针对假定原因的干预产生有利影响时,就可以推断因果关系。这种方法可适用于具有良好特征效果的饮食干预,以测试针对微生物组特征或给予微生物代谢物(假定原因)是否能产生预期效果。
人类研究可以通过动物模型扩展,以确定微生物组的因果作用,确定微生物组中的因果成分,并确定潜在机制。可以在无菌动物疾病模型中测试单独或作为群落的特定微生物(灵知动物)或与人类饮食干预生理效应相关的微生物代谢产物。灵长类动物允许去除或添加特定的微生物,以确定微生物群落中的致病成分,并且可以用模仿人类饮食的饲料进行挑战。
人类微生物相关(HMA)啮齿动物是最常用和最复杂的建立微生物组因果关系的模型,将人类粪便微生物群移植到啮齿类动物的疾病模型中。HMA动物可以非常有效地比较对饮食干预有或没有反应的人类微生物组,特别是如果假设差异是由特定微生物的存在或不存在引起的。然而,HMA动物在做出关于饮食诱导的人类微生物组组成变化的因果推断方面有很大的局限性。不提供活微生物的饮食不太可能从微生物群中添加或移除微生物物种,但只会改变群落中的相对比例。这种变化不太可能在受体动物身上复制,因为形成微生物群的生态和进化力量与供体中的不同。
尽管存在局限性,但动物模型,尤其是如果其微生物群得到良好控制并与多组学方法相结合,有能力补充人类研究,因为它们建立了饮食影响的机制基础。通过使用更好地模仿人类生理学的动物,如猪和灵长类动物,这些研究可以进一步改进。虽然不需要从机理和因果研究中获得见解,来确定和确认饮食成分的健康影响或基于微生物组的生物标记物的效用,但这对于完善假设非常宝贵,可以创新制定有针对性的营养策略,并为饮食建议提供额外的证据。
将肠道微生物组整合到营养学研究的所有阶段的实验框架,以促进对微生物组在健康饮食中的作用的理解:
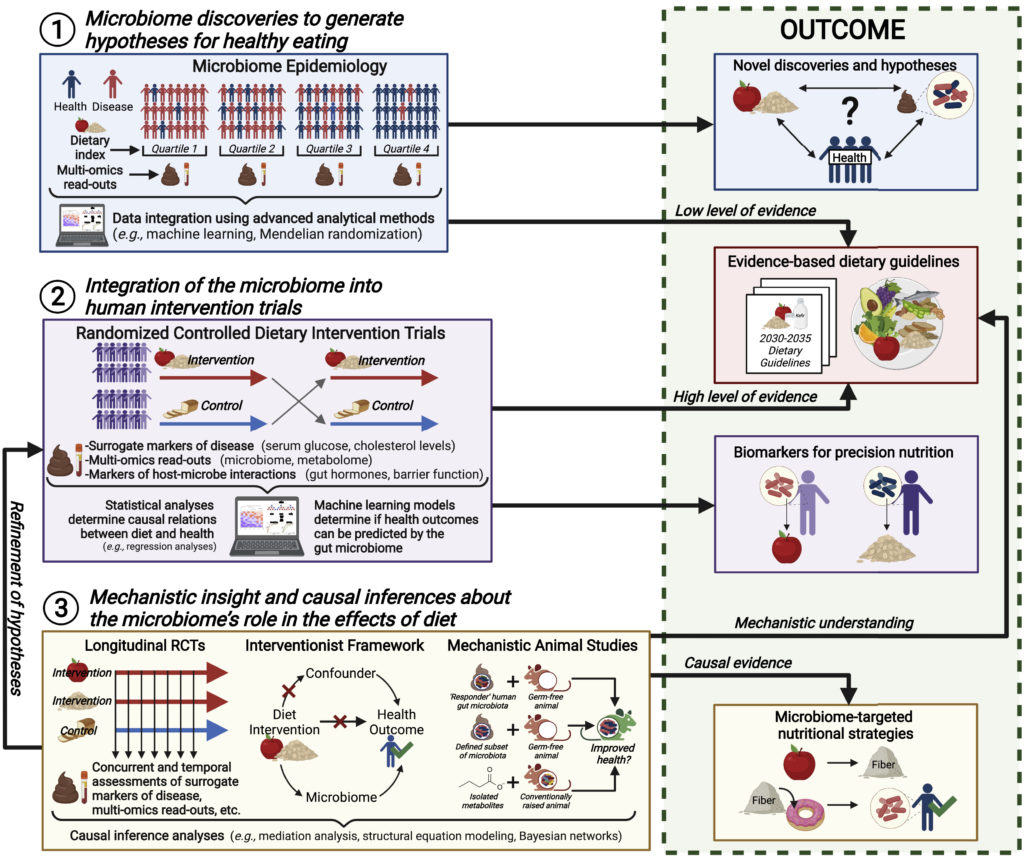
doi.org/10.1016/j.chom.2022.04.016
饮食与疾病密切相关,而疾病又与微生物群相关。肠道微生物群可能是构成营养研究的“黑匣子”,饮食—微生物群的相互作用可能有助于建立饮食生理效应的机制基础。
这两个领域有很强的生物学和进化论理由来扩展已经活跃和正在进行的合作,以加深对如何通过饮食优化健康的理解。
应将以微生物组为中心的终点纳入营养科学的各个方面,以加强饮食指南的证据基础。营养微生物学研究有可能全面了解健康饮食的各个方面,从而有助于解决饮食相关疾病的预防和管理。
谷禾也将不断提供关于肠道微生物群如何影响和介导饮食化合物、特定食物和饮食模式的生理效应的机制的前沿研究进展,应用到肠道菌群健康检测,不断更新已有框架,为大众提供针对微生物群的营养策略。
主要参考文献:
Armet AM, Deehan EC, O’Sullivan AF, Mota JF, Field CJ, Prado CM, Lucey AJ, Walter J. Rethinking healthy eating in light of the gut microbiome. Cell Host Microbe. 2022 Jun 8;30(6):764-785. doi: 10.1016/j.chom.2022.04.016. PMID: 35679823.
Lancaster SM, Lee-McMullen B, Abbott CW, Quijada JV, Hornburg D, Park H, Perelman D, Peterson DJ, Tang M, Robinson A, Ahadi S, Contrepois K, Hung CJ, Ashland M, McLaughlin T, Boonyanit A, Horning A, Sonnenburg JL, Snyder MP. Global, distinctive, and personal changes in molecular and microbial profiles by specific fibers in humans. Cell Host Microbe. 2022 Jun 8;30(6):848-862.e7. doi: 10.1016/j.chom.2022.03.036. Epub 2022 Apr 27. PMID: 35483363; PMCID: PMC9187607.
Deehan EC, Walter J. The Fiber Gap and the Disappearing Gut Microbiome: Implications for Human Nutrition. Trends Endocrinol Metab. 2016 May;27(5):239-242. doi: 10.1016/j.tem.2016.03.001. Epub 2016 Apr 11. PMID: 27079516.
Guthrie L, Spencer SP, Perelman D, Van Treuren W, Han S, Yu FB, Sonnenburg ED, Fischbach MA, Meyer TW, Sonnenburg JL. Impact of a 7-day homogeneous diet on interpersonal variation in human gut microbiomes and metabolomes. Cell Host Microbe. 2022 Jun 8;30(6):863-874.e4. doi: 10.1016/j.chom.2022.05.003. Epub 2022 May 27. PMID: 35643079.

谷禾健康

Faecalibacterium prausnitzii(F. prausnitzii),普氏栖粪杆菌,又名:普拉梭菌,是人类肠道菌群中最重要的细菌之一,占健康人粪便样本中检测到的细菌总数的 5-15%,是丁酸的重要生产者之一,具有抗炎作用,维持细菌酶的活性,保护消化系统免受肠道病原体的侵害。
已经证实,患有慢性便秘、乳糜泻、肠易激综合征和炎症性肠病(包括克罗恩病和溃疡性结肠炎)的个体中这种微生物的数量减少。此外,在 2 型糖尿病、结直肠癌和银屑病患者中观察到较低水平的F. prausnitzii。
F. prausnitzii 数量的变化可能是人类肠道菌群失调的迹象,科学家们将其视为下一代益生菌的候选者。
Faecalibacterium prausnitzii,革兰氏阴性,对氧极度敏感,是柔嫩梭菌类群的优势菌,属于梭菌科,厚壁菌门。该物种属于Clostridium cluster IV分组的Clostridium leptum group柔嫩梭菌类群,是该类群的最优势菌种,占64%左右。一般中文翻译柔嫩梭菌指的就是这个类群,其代表物种就是普氏栖粪杆菌,又名普拉梭菌。
普拉梭菌的扫描电子显微镜图像
Plateforme MIMA 2, T. Meylheuc
直到 2000 年代中期,16S rRNA 文库的高通量测序和粪便 DNA 的宏基因组分析显示F. prausnitzii是人类肠道中最丰富的细菌之一,人们才完全意识到该物种对人类健康的重要性细菌总数的 5–15%。
在婴儿早期,Faecalibacterium prausnitzii 的数量非常低,并在原定殖细菌建立后增加。同时,在克罗恩病 (CD)和溃疡性结肠炎 (UC) 等各种炎症性肠病 (IBD) 以及结直肠癌 (CRC) 和 2 型糖尿病中观察到 F. prausnitzii 水平降低。
F. prausnitzii 生长繁殖肠道理化调节
F. prausnitzii 生长的最佳 pH 值范围在 5.7 和 6.7 之间,这是在结肠中发现的 pH 值范围。尽管在 5-5.7 的 pH 值范围内菌株之间的耐受性存在差异,但在 3.5 和 4.5 的 pH 值之间没有观察到生长。这表明 pH 值影响F. prausnitzii 沿肠道的分布。
该菌在健康受试者和肠道疾病患者的十二指肠(pH 范围 5.7-6.4)和回肠末端中也被检测到。据报道,溃疡性结肠炎和克罗恩病患者经常出现酸性大便,因此仍有待证明肠道局部 pH 值是否会调节F.prausnitzii 在肠道疾病(如 IBD)患者中的丰度和组成。
F. prausnitzii 对胆汁盐生理浓度的轻微增加也高度敏感,这为克罗恩病患者表现出的F. prausnitzii丰度减少提供了一个合理的解释,因为这些患者的胆红素浓度增加,尤其是那些有回肠疾病受累和接受过肠切除术的患者。
此外,据报道,分离株之间的耐受性存在差异,尤其是胆盐浓度为 0.1% (wt/vol) 时,表明胆汁盐浓度的变化可能决定F. prausnitzii亚型组成的变化。由于克罗恩病患者的胆盐成分也发生改变,需要进一步研究以确定F. prausnitzii是否对某些类型的胆盐成分具有更高的敏感性,并确定不同的胆汁盐谱是否会改变 F. prausnitzii 亚型组成。
F. prausnitzii对氧极为敏感,但它能够通过在黄素和半胱氨酸或谷胱甘肽存在的情况下使用细胞外电子转移来承受肠粘膜中发现的低水平氧气 。最近,已经证明,当与这些抗氧化剂和菊粉作为冷冻保护剂一起配制时,菌株 A2-165 可以在环境空气中保持活力 24 小时。因为氧气梯度在定义结肠中微生物的空间组织方面具有重要作用,确定F. prausnitzii亚型之间的氧耐受性是否存在差异,以及它是否与粘膜的发炎状态相关,将会很有趣。
F. prausnitzii的必需营养素的可用性和有效性可能会影响其在肠道中的分布。
F. prausnitzii的全局营养需求已经通过人工策划的代谢重建进行了调查,该重建确定了几种氨基酸和维生素作为生长因子。体外实验表明,F. prausnitzii可以通过代谢交叉喂养直接或间接地代谢纤维。乙酸盐消耗是F. prausnitzii生产丁酸盐的主要驱动力。当这两个物种在低聚果糖上一起生长时,青春双歧杆菌释放的醋酸盐的消耗已被证明会刺激F. prausnitzii。F. prausnitzii可以代谢来自宿主和饮食来源的底物,菌株之间存在一些差异。
F. prausnitzii分离株可以使用简单碳水化合物生长良好,但菌株之间在发酵更复杂碳水化合物的能力方面存在一些差异,例如那些来自宿主或饮食的碳水化合物,尽管大多数F. prausnitzii菌株都能够发酵菊粉,但研究结果表明,其中只有两种菌株可以在该基质上生长良好。这支持观察到该益生元在营养干预中对该物种的刺激,并表明只有F. prausnitzii种群的一些成员受到菊粉的选择性刺激。
该物种的菌株利用肠腔中发现的其他多糖的能力有限,例如阿拉伯半乳聚糖、木聚糖和可溶性淀粉。大多数分离株可以在苹果果胶上生长,并且能够使用一些果胶衍生物。体外研究表明,在生理条件下,F. prausnitzii可以在某些类型的果胶发酵中发挥关键作用,并且可以与其他肠道细菌成功竞争这种底物。
最近一项基于普拉梭菌 A2-165菌株功能代谢图的研究预测其无法合成氨基酸丙氨酸、半胱氨酸、蛋氨酸、丝氨酸和色氨酸。通过对其他F. prausnitzii菌株基因组的进一步分析,已经观察到维生素和辅助因子(例如:生物素、叶酸、烟酸、泛酸、吡哆醇和硫胺素)的营养缺陷型,并且菌株之间似乎存在与核黄素生产有关的一些差异,这可能是由于菌株间差异。
相比之下,该物种已被预测为钴胺素生产者。有证据表明,一些 IBD 患者易患钴胺素缺乏症,但尚未确定这种情况的原因。由于缺乏一致的临床数据表明 IBD 患者易患这种缺陷,因此确定它是否与肠道中钴胺素生产者的消耗有关会很有价值。
F. prausnitzii与肠道微生物群其他成员的关系
F. prausnitzii在肠道中与C. coccoides组和 Bacteroidetes 的几个成员共同发生。有人提出F. prausnitzii可能依赖其他物种(如拟杆菌)进行交叉饲养。在共培养实验中,观察到F. prausnitzii 的发酵活动过程中,多形拟杆菌(B. thetaiotaomicron)正在发酵果胶。表明后者产生的乙酸盐促进了F. prausnitzii 的生长。此外,B. thetaiotaomicron对果胶的初始发酵可以释放果胶衍生物,然后可以被F. prausnitzii使用。
最近对大鼠模型的研究表明,F.prausnitzii 需要预先存在后,B.thetaiotaomicron才能在肠道定植。已经反复观察到无法维持F. prausnitzii单相关动物模型 并且还描述了一种小鼠模型,其中F. prausnitzii植入胃肠道需要用大肠杆菌预先制备,表明F. prausnitzii和其他菌群存在密切关系。在 IBD 患者中发现了这两种物种之间的相关性,根据疾病位置观察到正相关或负相关。这表明一个种群对另一个种群的影响,尽管还不能排除宿主因素的影响。
F. prausnitzii 的分类学和系统发育
已经确定Faecalibacterium属与厚壁菌门、梭菌类和瘤胃球菌科中的梭菌群 IV的成员有关。目前,F. prausnitzii是唯一成功分离的粪杆菌属物种。
最近对分离株的系统发育表征确定该物种包括两个系统发育群,它们具有 97% 的 16S rRNA 基因序列相似性。其中分类群 I 的减少是肠道炎症的更敏感标志物。迄今为止,大部分可用的基因组和生理数据是使用分类群 II 菌株收集的。
F. prausnitzii基因组之间的平均核苷酸同一性水平非常低,而基因组的可塑性却很高。两个基因组可以根据功能基因互补的差异进行分离,尽管这种划分与基于保守基因系统发育的分离不完全一致,突出了水平基因转移在塑造F. prausnitzii 中的重要性基因组。两个基因组之间的差异主要在于与碳水化合物分解代谢相关的基因(例如分类群 I中预测的唾液酸酶基因)和氨基酸的互补,以及防御机制。
值得注意的是,最近对健康和患病肠道样本中物种多样性和丰度的研究表明,存在其他F. prausnitzii系统发育型,并且不能排除Faecalibacterium属内其他物种的存在。这些已经通过分析粪便样本中总体细菌群落的分子方法估计,代表大约 2% 的粪杆菌序列,并使用物种特异性引物。有趣的是,罕见的系统发育型主要是从患有肠道疾病的受试者身上发现的。
产生丁酸,维持肠道稳态
Faecalibacterium prausnitzii 一直被报道为肠道中发现的主要丁酸盐生产者之一。丁酸盐在肠道生理学和宿主健康中起着至关重要的作用。丁酸盐结肠细胞的主要能量来源之一,能够保持肠道内壁的完整性,防止病原体通过肠道进入人体。它刺激绒毛的生长 (肠道屏障上的微观挤压)并促进粘蛋白的产生,粘蛋白是一种覆盖肠道内壁的保护性凝胶。此外,丁酸盐通过增加紧密连接蛋白的合成和抗菌肽的产生来参与结肠屏障的增强。
肠道屏障依赖于 T 连接蛋白来防止肠道内容物泄漏到血液中。丁酸盐通过激活 AMP 活化蛋白激酶 (AMPK) 来促进紧密结合的 T 连接蛋白。
丁酸盐可以通过调节肠腔 pH 值来调节肠道微生物群,这对产生短链脂肪酸的细菌有益,丁酸盐可以保持上皮缺氧并限制硝酸盐呼吸依赖性细菌的过度生长以维持肠道稳态。
丁酸盐是由某些细菌在乳酸发酵过程中产生的。然而,乳酸也是硫酸盐还原菌的首选能源,硫酸盐还原菌会产生硫化物并抑制结肠细胞对丁酸的氧化。IBD 中产生丁酸盐的菌的消耗可能会削弱已经脆弱的肠上皮细胞,导致共生或低致病性细菌的入侵,随后激活免疫活性细胞。
产生丁酸,抗炎抗肿瘤
丁酸盐可以通过抑制 NF-κB 转录因子激活、上调 PPARγ和抑制干扰素γ来减轻肠黏膜炎症。
此外,F. prausnitzii已被证明可以产生许多具有抗炎特性的物质,包括一种 15 kDa 的蛋白质“微生物抗炎分子”(MAM),它能够抑制肠上皮中的 NF-κB 通路细胞和预防小鼠 IBD 模型中的结肠炎 。而且,丁酸盐可能通过抑制组蛋白脱乙酰酶活性来抑制炎症反应,导致组蛋白过度乙酰化和抑制 NF-κB 活性。
Faecalibacterium prausnitzii 上清液已被证明可以抑制促炎性 IL-8 免疫蛋白的产生。
F. prausnitzii 还通过激活产生 IL-10(一种抗炎蛋白)的 T 细胞来帮助对抗炎症。
F. prausnitzii 的抗炎机制
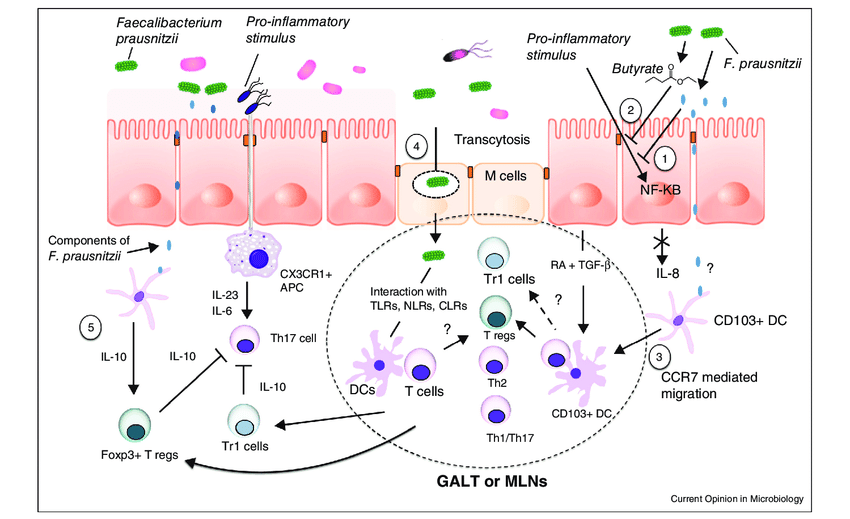
额外的抗炎特性归因于该菌还通过其诱导耐受性细胞因子谱的能力(促炎细胞因子如 IL-12 和 IFN-γ 的分泌非常低,而抗炎细胞因子 IL-10 的分泌增加)。这些抗炎作用部分与能够阻断 NF-κB 激活、IL-8 产生和调节性 T 细胞产生上调的分泌代谢物有关。
这些微生物可以产生莽草酸,一种常见于植物中的酸,能够防止脂多糖 (LPS) 引起的炎症。LPS 是一些细菌外膜上的毒素,可以激活免疫系统并引起炎症。F. prausnitzii 还可以制造水杨酸,这有助于防止细菌形成生物膜,这是一些引起感染的微生物的常见特征。
体外研究表明,丁酸盐还具有抗肿瘤作用,包括减少肠上皮细胞中肿瘤坏死因子 (TNF) 的分泌,并诱导肿瘤细胞的分化和凋亡,从而抑制肿瘤生长。
F. prausnitzii 及其代谢物对小鼠结肠炎发挥保护作用,改善了肠道菌群失调,增加了细菌多样性和产生短链脂肪酸菌的丰度,降低了血清 TNF-α 和Proteinbacteria,酸杆菌门和拟杆菌。这些发现将为F. prausnitzii 的抗炎抗肿瘤作用提供进一步的证据,其具有治疗 IBD 的潜力。

下一代益生菌特性
大多数商业益生菌不包括有益健康的优势肠道微生物菌株,这就是为什么这些益生菌菌株不会在人类肠道中定殖并且它们的作用仅在很短的时间内持续存在的原因。如今,人们对使用共生细菌作为潜在的益生菌剂越来越感兴趣。
使用厌氧细菌和 EOS(氧敏感) 细菌是成功分离可能影响肠道稳态的代表性菌株的先决条件。Faecalibacterium prausnitzii从健康志愿者分离株:F. prausnitzii作为新一代益生菌在使用中向前迈进了一步。
但是下一代共生益生菌必须满足与传统益生菌相同的标准。它们应该:
(i) 被隔离和充分表征
(ii) 达到安全要求,例如可接受的抗生素抗性或缺乏裂解和粘附能力
(iii) 作为益生菌考虑,在被隔离之前对宿主表现出有益的影响
关于F. prausnitzii,虽然对其安全性知之甚少,但该物种作为下一代益生菌具有明显的潜力。到目前为止,所有F. prausnitzii分离株具有一些特征,例如:
(i) 缺乏上皮细胞粘附、质粒、抗微生物和溶血活性
(ii) 存在 DNAse 活性。部分的F. prausnitzii菌株产生的 D-乳酸微弱,不会对宿主产生代谢有害影响,无法降解黏蛋白。
仅使用参考菌株F. prausnitzii A2-165在体外和体内分析了其有益的抗炎作用和生物膜形成菌株 HTF-F。由于益生菌特性通常是菌株特异性的,需要单独研究来评估其他F. prausnitzii分离菌株的抗炎特性。
人体微生物组中 F. prausnitzii 的丰度受其健康状况和结肠环境的影响。这一点尤其重要,因为在患有消化系统疾病的患者中发现了低于平均水平的F. prausnitzii。
克罗恩病和溃疡性结肠炎
肠道微生物群可能通过两种机制驱动致病性,即“促炎”菌群的扩张或微生物群保护性化合物的限制。
健康肠道和IBD患者中 F. prausnitzii
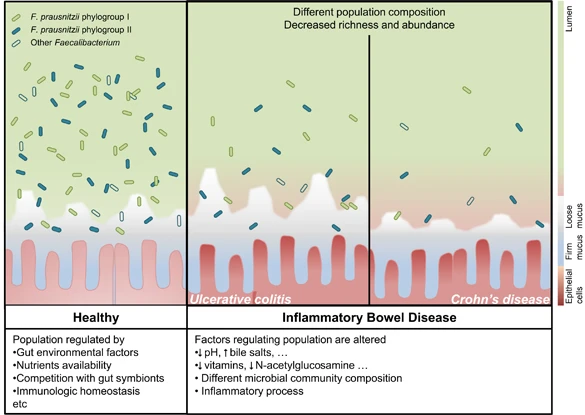
在IBD患者中,肠道环境的改变可能会影响F. prausnitzii 的种群组成和负荷。这些差异可以通过监测来鉴别IBD亚型。
在过去的十年的研究已发现,相当数量的降低的F. prausnitzii水平与某些形式的IBD相关。虽然F. prausnitzii水平与溃疡性结肠炎活动和隐窝炎之间的联系是有争议的,在克罗恩病中F. prausnitzii枯竭,尤其是在疾病发作和克罗恩病的回肠部位。
◥ 治疗后F. prausnitzii恢复
在粪便和活检样本中使用的各种证明方法表明,对克罗恩病患者有效的各种抗炎和抗菌治疗,包括大剂量皮质醇、英夫利昔单抗、干扰素-α2b 和利福昔明,都能够恢复正常水平的F. prausnitzii。
因此,有人提出F. prausnitzii的消耗不是克罗恩病的致病事件,而是产生过量活性氧 (ROS) 的粘膜炎症的结果。这导致粘膜相关和管腔 EOS 培养物显着减少,包括F. prausnitzii。
如何解释Faecalibacterium prausnitzii在溃疡性结肠炎和克罗恩病患者中减少?
◥ pH值
健康的肠道 pH 值 (5.7–6.7) 为 F. prausnitzii 的生长提供了最佳条件,而高酸性环境会抑制 Faecalibacterium prausnitzii 的生长。尤其是,溃疡性结肠炎和克罗恩病患者的大便呈酸性,这可能有助于解释为什么他们的微生物群中缺乏 F. prausnitzii .
◥ 胆汁
胆汁及其成分水平略微升高也会影响 F. prausnitzii 丰度。胆汁是肝脏分泌的一种物质,可在消化过程中分解脂肪。胆汁盐已被证明可以抑制 Faecalibacterium prausnitzii。
研究人员假设这种对胆汁的敏感性可以解释为什么这些微生物在克罗恩病患者中较少。患有这种炎症性疾病的人往往有较高水平的胆红素,胆红素是一种赋予胆汁颜色的色素,特别是如果他们已经切除了部分肠道。
溃疡性结肠炎中, F. prausnitzii有缺陷的肠道定植,这在缓解期短、既往有频繁复发史和结肠炎大范围扩展的患者中更为严重。此外,复发后,F. prausnitzii在维持缓解时逐渐恢复,1年后达到接近参考种群的值。
基于F. prausnitzii菌株特性的治疗探索
科学研究人员提出假设,即F. prausnitzii 的高定植可能会阻止溃疡性结肠炎中粘膜炎症过程的重新激活。这一假设应在未来的研究和增加肠道F. prausnitzii 的策略中得到证实需要在患有广泛疾病的患者中对人群进行验证。
F. prausnitzii菌株还可以利用N-乙酰氨基葡萄糖,一种在肠粘膜中发现的糖蛋白的成分。有趣的是,用这种化合物进行治疗可能会改善克罗恩病,因为它可以作为炎症、受损肠道软组织的愈合因子。因此,鉴于F. prausnitzii能够发酵这种碳水化合物,探索在接受这种治疗的克罗恩病患者中恢复这种有益肠道细菌的效果将是有意义的。
镇痛特性
慢性腹痛是肠易激综合征或炎症性肠病等肠道疾病患者的常见症状之一。研究人员表明,在炎症性肠病的患者中,肠道中的Faecalibacterium prausnitzii细菌的丰度降低。
这些研究人员已经证明,由于压力动物模型(产生绞痛起源的内脏超敏反应),F. prausnitzii丰度的减少与内脏超敏反应的出现有关。对于人类,这种超敏反应会导致肠道不适或腹痛。
科学家已经证明,通过给这些动物服用F. prausnitzii,它们恢复了正常的绞痛敏感性。
结直肠癌
丁酸盐的抗炎功能包括抗癌特性。这种短链脂肪酸是一种组蛋白脱乙酰酶 (HDAC) 抑制剂:它抑制癌细胞的活性和生命周期。这促使研究人员调查 Faecalibacterium prausnitzii 在结直肠癌中的作用。
结直肠癌患者产丁酸盐的细菌较少,包括Faecalibacterium prausnitzii。有人提出,由于丁酸盐产量较低,肠道内壁细胞可能更容易受到损伤,这可能会增加患癌症的风险。
扩展阅读:结直肠癌防治新策略——微生物群
乳腺癌
F.prausnitzii 通过抑制IL-6/STAT3通路抑制乳腺癌细胞的生长。菌群代谢物与菌群结合(如Faecalibacterium与磷胆碱结合)可能是乳腺癌的一种新的检测方法。
糖尿病
糖尿病是一种代谢性疾病,其中由于胰岛素功能受损,身体失去控制血糖的能力。升高的循环葡萄糖会损害血管并增加患心脏病的风险。
许多针对人类的研究已经确定了肠道微生物组的失衡(生态失调),这会削弱肠道内壁,并使不良分子进入体内,从而破坏其他器官。科学家们认为,生态失调先于并促进了糖尿病的发展。
糖尿病患者的厚壁菌门水平较低,包括 Faecalibacterium prausnitzii 和其他可强化肠道内壁的产丁酸盐细菌。
研究表明,脂多糖是在某些革兰氏阴性细菌上发现的促炎分子,当肠道屏障薄弱时,它会进入血液并损害胰腺中的B 细胞功能,产生胰岛素的细胞。
过敏反应
Faecalibacterium prausnitzii通过调节肠道微生物群和短链脂肪酸的产生来缓解屋尘螨引起的过敏性哮喘。
补充F. prausnitzii可阻断嗜酸性粒细胞、中性粒细胞、淋巴细胞和巨噬细胞的流入,减轻病理变化。此外,活的和死的F. prausnitzii给药都降低了白细胞介素 IL-4、IL-5、IL-13 和免疫球蛋白 G1 的水平,提高了调节性 T 细胞 (Treg) 的比例,改善了微生物生态失调并增强了短链脂肪酸生产。
Faecalibacterium prausnitzii部分通过肠道微生物群调节和短链脂肪酸产生发挥抗哮喘作用,表明其作为预防过敏性哮喘的益生菌剂的潜力。
肝病
研究人员研究了口服F. prausnitzii治疗对高脂肪喂养小鼠的影响。与高脂肪对照小鼠相比,F. prausnitzii处理的小鼠肝脏脂肪含量、天冬氨酸氨基转移酶和丙氨酸氨基转移酶较低,肝脏中脂肪酸氧化和脂联素信号传导增加。肝脏脂质组学分析显示,几种甘油三酯、磷脂和胆固醇酯的含量减少。内脏脂肪组织中的脂联素表达增加,皮下和内脏脂肪组织对胰岛素更敏感。
且F. prausnitzii治疗增加了肌肉质量,这可能与增强线粒体呼吸、改变肠道微生物群组成和改善肠道完整性有关。总的来说,F. prausnitzii治疗可改善肝脏健康,减少小鼠脂肪组织炎症,需要进一步研究以发现其治疗潜力。
其它
肠道菌群中F. prausnitzii的消耗与微生物失调有关,同时伴随着广泛的代谢和/或免疫介导的慢性疾病,包括银屑病、高血压、心脏和肾脏疾病。
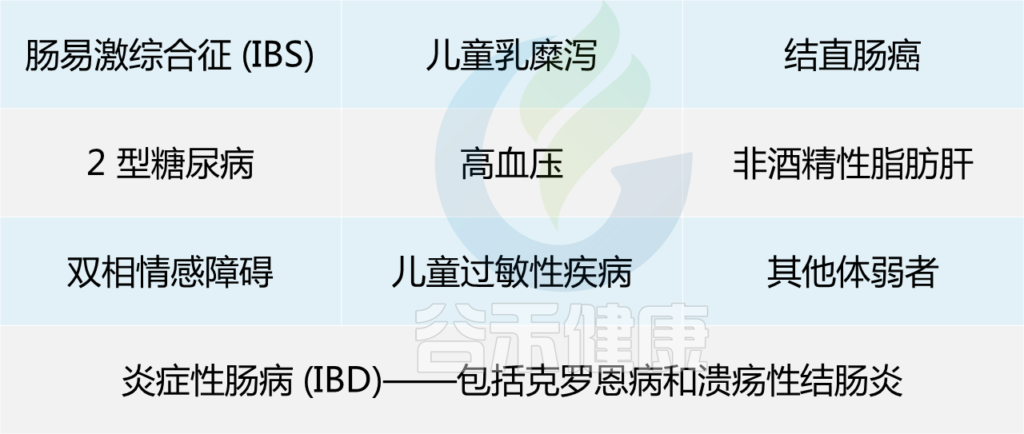
在以下患者中已显示低水平的F. prausnitzii:
综上所述,F. prausnitzii 对人体肠道和健康非常重要,以至于它被称为肠道中的“关键物种”。现在问题来了:如何增加肠道中这种细菌的数量?
能否添加到酸奶中补充?
不能。
添加到酸奶中或作为补充剂的典型细菌在暴露于空气(氧气)时能够存活。然而,F. prausnitzii是“氧敏感的”,它们在暴露于空气后几分钟内就会死亡。研究人员将这种有益细菌视为“未来的益生菌”,目前正在进行研究以找出它可以轻松储存并暴露在空气中几个小时而不会死亡的方法。所以目前没有办法服用益生菌F. prausnitzii补充剂。
除此之外还能做什么呢?
饮食。
高动物肉、高动物脂肪、高糖、高度加工食品和低纤维饮食(典型的西化饮食)会降低F. prausnitzii 的数量,而高纤维、低肉的饮食会增加 F. prausnitzii 的数量。
——高纤维
我们可以做的第一个提高 F. prausnitzii 的数量的办法, 就是增加饮食中的纤维。增加膳食纤维会增加丁酸盐,丁酸盐与结肠健康有关,具有抗炎和抗癌作用。
高纤维是包括:全谷物、蔬菜、水果、坚果、种子和豆类。此外,还要主要吃多样化的植物性饮食,也就是说大量的植物性食物。多样性似乎很重要——不同类型的纤维喂养不同的细菌。
——益生元
菊粉型果聚糖和阿拉伯木聚糖等益生元可增加F. prausnitzii的数量。

一项研究表明,猕猴桃胶囊的摄入增加了功能性便秘患者的Faecalibacterium prausnitzii 丰度。
虽然F. prausnitzii可能是肠道中一种重要的有益细菌,但是它与其他有益细菌的关系,它们是交叉喂养的。研究表明青春双歧杆菌是利用菊粉主要物种并刺激Faecalibacterium prausnitzii的增加。
主要参考文献:
Heinken, A., Khan, M. T., Paglia, G., Rodionov, D. A., Harmsen, H. J. M., & Thiele, I. (2014). Functional Metabolic Map of Faecalibacterium prausnitzii, a Beneficial Human Gut Microbe. Journal of Bacteriology, 196(18), 3289–3302. doi:10.1128/jb.01780-14
Rivière, A., Selak, M., Lantin, D., Leroy, F., & De Vuyst, L. (2016). Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Frontiers in Microbiology, 7. doi:10.3389/fmicb.2016.00979
Lopez-Siles, M., Duncan, S. H., Garcia-Gil, L. J., & Martinez-Medina, M. (2017). Faecalibacterium prausnitzii: from microbiology to diagnostics and prognostics. The ISME Journal, 11(4), 841–852. doi:10.1038/ismej.2016.176
Moreno-Indias, I., Sánchez-Alcoholado, L., Pérez-Martínez, P., Andrés-Lacueva, C., Cardona, F., Tinahones, F., & Queipo-Ortuño, M. I. (2016). Red wine polyphenols modulate fecal microbiota and reduce markers of the metabolic syndrome in obese patients. Food & Function, 7(4), 1775–1787. doi:10.1039/c5fo00886g
Richardson, D. P., Ansell, J., & Drummond, L. N. (2018). The nutritional and health attributes of kiwifruit: a review. European Journal of Nutrition. doi:10.1007/s00394-018-1627-z
Blatchford, P., Stoklosinski, H., Eady, S., Wallace, A., Butts, C., Gearry, R., … Ansell, J. (2017). Consumption of kiwifruit capsules increases Faecalibacterium prausnitzii abundance in functionally constipated individuals: a randomized controlled human trial. Journal of Nutritional Science, 6. doi:10.1017/jns.2017.52
Lopez-Siles M, Duncan SH, Garcia-Gil LJ, Martinez-Medina M. Faecalibacterium prausnitzii: from microbiology to diagnostics and prognostics. ISME J. 2017 Apr;11(4):841-852. doi: 10.1038/ismej.2016.176. Epub 2017 Jan 3. PMID: 28045459; PMCID: PMC5364359.
Ramirez-Farias C, Slezak K, Fuller Z, Duncan A, Holtrop G, Louis P. Effect of inulin on the human gut microbiota: stimulation of Bifidobacterium adolescentis and Faecalibacterium prausnitzii. Br J Nutr. 2009 Feb;101(4):541-50. doi: 10.1017/S0007114508019880. Epub 2008 Jul 1. PMID: 18590586.
Varela E, Manichanh C, Gallart M, Torrejón A, Borruel N, Casellas F, Guarner F, Antolin M. Colonisation by Faecalibacterium prausnitzii and maintenance of clinical remission in patients with ulcerative colitis. Aliment Pharmacol Ther. 2013 Jul;38(2):151-61. doi: 10.1111/apt.12365. Epub 2013 Jun 3. PMID: 23725320.
Munukka, E., Rintala, A., Toivonen, R. et al. Faecalibacterium prausnitzii treatment improves hepatic health and reduces adipose tissue inflammation in high-fat fed mice. ISME J 11, 1667–1679 (2017). doi.org/10.1038/ismej.2017.24