 国家高新企业 | ISO9001认证
国家高新企业 | ISO9001认证 二级病原微生物安全实验室
二级病原微生物安全实验室- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
国家高新企业 | ISO9001认证 二级病原微生物安全实验室
谷禾健康

你敢相信吗?从粪便中提取出具有强烈粪臭味的物质,当用酒精稀释上千倍后,脱胎换骨变成了一种香味。这就是一种吲哚衍生物——3-甲基吲哚(又名粪臭素)
吲哚,是所有花香类原精的关键成分,这种物质在低剂量1-3%浓度时,具有放松的作用,协同苯基酯,能强效抗痉挛,增添感官愉悦和魅力。然而,如果简单将吲哚等同于香水,未免太少看它的功效。
在谷禾特定肠道菌群检测报告中炎症部分或毒素部分有吲哚这个指标。
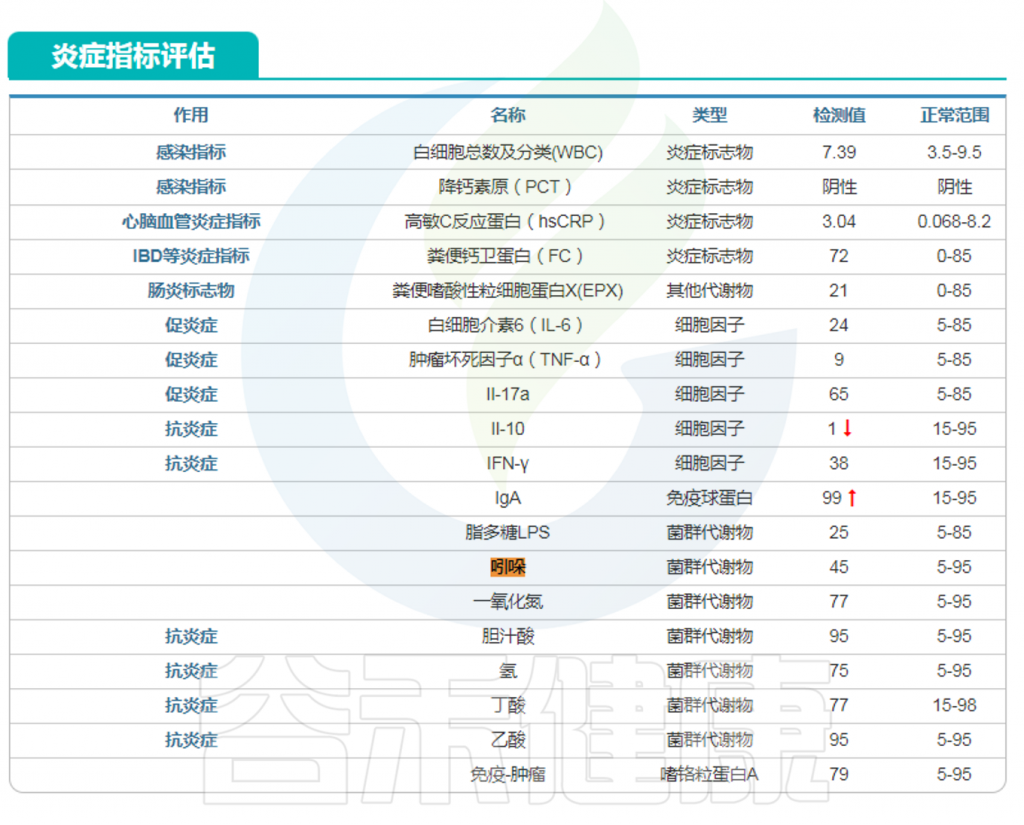
<来源:谷禾肠道菌群检测数据库>

<来源:谷禾肠道菌群检测数据库>
很多人可能对与菌群和健康相关的血清素,色氨酸,脂多糖,组胺等指标比较熟悉,但是对吲哚还是比较陌生,实际上吲哚与肠道微生物群和人类健康和生活关系密切,例如,吲哚衍生物可以影响胃肠道的蠕动,可能会影响便秘或腹泻等状况;吲哚及其衍生物已被证明可以加强肠道屏障功能,有助于防止病原体和有害物质进入血液。
事实上,由吲哚衍生而来的各种化学物质,对人体神经调节和抗病原微生物,有极为积极的作用。在人体中存在着许多这样神奇的吲哚类物质。内源性吲哚及其衍生物是源自肠道微生物群的色氨酸代谢物,具有一系列生物活性。
最近的研究表明,吲哚有助于维持人体肠道的生物屏障,主要通过激活AhR和PXR受体来发挥抗炎活性,从而影响免疫系统的功能,显著改善肠道健康(炎症性肠病、出血性结肠炎、癌症结直肠癌),并进一步促进人类健康(糖尿病、中枢系统炎症和血管调节)。
作为细胞间信号分子,吲哚还控制细菌生理学的各个方面,例如产吲哚细菌的孢子形成、质粒稳定性、耐药性、生物膜形成和毒力。相比之下,许多不产生吲哚的细菌、植物和动物产生多种加氧酶,可能会干扰吲哚信号传导。
吲哚还是身体神经系统与内分泌系统正常与否的风向标。所谓的体臭,其实就是人体内吲哚含量增高的表现。吲哚在细菌生理学、生态平衡以及人体健康中发挥着重要作用。
然而,吲哚的毒性影响也不容忽视。过量的吲哚会对情绪和行为产生不利影响,硫酸吲哚酚是一种吲哚衍生物,具有肾毒性和心血管毒性。羟吲哚还可能降低神经元兴奋性。
吲哚与肠道微生物群之间的相互作用以及吲哚对人类健康的影响是一把双刃剑。本文主要讲述了内源性吲哚的生物合成、吲哚对肠道功能及微生物群的影响,为吲哚与疾病的相关性提供了科学依据。
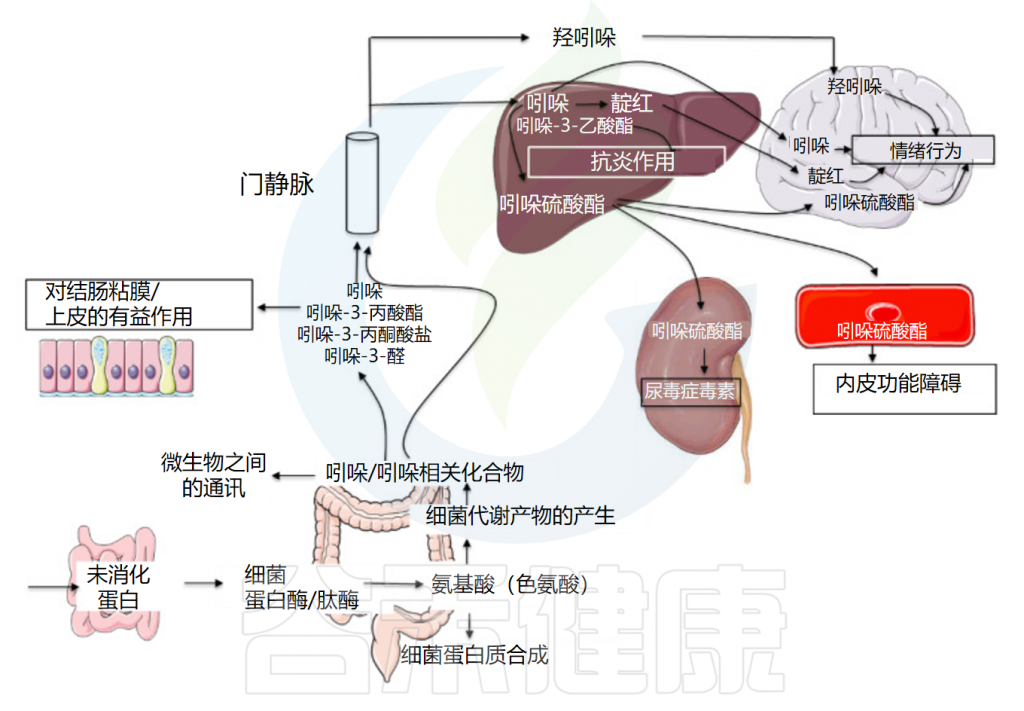
Tennoune N,et al., Microorganisms.2022
吲哚(Indole)是一种有机化合物,是吡咯与苯并联的化合物,又称苯并吡咯。吲哚及其同系物和衍生物广泛存在于自然界,主要存在于天然花油,如茉莉花、苦橙花、水仙花、香罗兰等。
而内源性吲哚及其衍生物是来源于肠道菌群的色氨酸代谢物,具有一系列生物活性。内源性吲哚类物质包括吲哚本身以及吲哚的代谢产物,如吲哚-3-甲酸、吲哚-3-丙酸、硫酸吲哚酚等。
吲哚相关化合物的结构
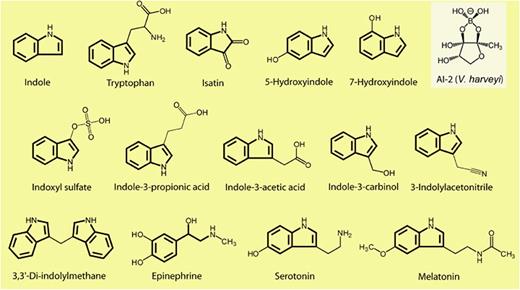
Lee JH, Lee J.FEMS Microbiol Rev.2010
肠道菌群产生吲哚类物质
细菌蛋白酶和肽酶从可用的膳食中释放氨基酸,包括色氨酸。而色氨酸是吲哚的主要前体。大部分色氨酸在小肠中被吸收,约4%~6%的色氨酸在结肠中被肠道菌群分解代谢为吲哚。
色氨酸作为一种必需氨基酸,主要从富含蛋白质的食物中获取,是唯一含有吲哚的氨基酸。
肠道中的色氨酸通过三个主要途径进行代谢:1) 犬尿氨酸途径(摄入的色氨酸的95%);2)细菌色氨酸代谢(4-6%);
3) 血清素途径 (1-2%)
✦ 肠道菌群主要通过色氨酸酶将色氨酸转化为吲哚
肠道细菌主要通过色氨酸酶(TnaA)的作用将色氨酸转化为吲哚;色氨酸酶TnaA可逆地将色氨酸降解为吲哚、丙酮酸和氨。
值得注意的是,在哺乳动物中,吲哚完全来源于细菌代谢活动,因为宿主细胞不具有产生这种化合物的代谢能力。
色氨酸也可以通过肠道微生物群直接或间接转化为几种吲哚相关化合物,包括吲哚-3-丙酮酸、吲哚-3-乳酸、吲哚-3-丙酸、吲哚-3-乙酰胺、吲哚-3-丙烯酸、吲哚乙醛、吲哚-3-乙酸、3-甲基吲哚(粪臭素)等。
✦ 内源性吲哚产生的机制
大肠杆菌(E. coli)是研究最广泛的吲哚合成生物体,它可以使用多种机制(抑制、转录衰减和反馈抑制)来调节色氨酸代谢中的色氨酸操纵子(Trp operon)和tna操纵子的表达。
在色氨酸缺乏的条件下, trp操纵子的表达升高,而由TnaC(24个氨基酸前导肽,以前称为TnaL)、TnaA(色氨酸酶)和TnaB(通透酶)组成的tna操纵子的表达受到抑制,因为转录终止因子(Rho)依赖性终止发生在tna操纵子中。
结果,当色氨酸水平较低时,TnaA和TnaB的表达以及吲哚的产生受到抑制。在富含色氨酸的条件下,转录终止因子依赖性转录终止被消除,因此吲哚产量增加。
因此,细胞外色氨酸和其他氨基酸直接影响吲哚的产生。
// 拓展:渗透酶通过转运也会影响吲哚水平
此外,三种渗透酶(Mtr、TnaB和AroP)在不同环境的色氨酸转运中发挥不同的作用,因此可以直接影响吲哚的水平。
Mtr通透酶主要负责转运吲哚,而TnaB通透酶对于色氨酸的摄取至关重要。尽管色氨酸的生产成本高昂,但细胞仍然利用色氨酸途径大量生产和分泌吲哚。
由色氨酸产生的吲哚可以通过细胞膜蛋白进行转运。例如,大肠杆菌和霍乱弧菌在丰富的培养基中可以分泌高达0.6 mM的吲哚。
在大肠杆菌中,外排蛋白AcrEF部分负责吲哚的输出,因为AcrEF突变体的吲哚排泄低于其野生型菌株。Mtr通透酶主要负责输入吲哚,因为吲哚不被mtr突变体吸收。
然而,最近有人提出,吲哚由于其疏水性,可能可以直接扩散通过细胞膜。因此,有必要清楚地了解吲哚的进出口情况。
大肠杆菌中吲哚生物合成和吲哚信号传导
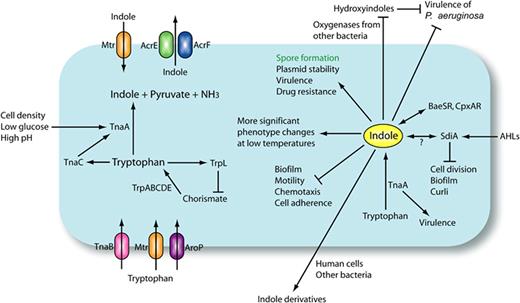
Lee JH, Lee J.FEMS Microbiol Rev.2010
人体粪便中吲哚的浓度较高,通常范围为0.25毫摩尔浓度~1.1毫摩尔浓度,但可能由于人群之间肠道微生物群组成的差异以及膳食蛋白质消耗水平不同而存在差异区别。
注:由于吲哚似乎是通过结肠上皮吸收的,结肠中吲哚的浓度可能高于粪便中记录的浓度,远端大肠的浓度高于近端部分的浓度。
尽管大多数生物体都含有色氨酸代谢途径,但只有能编码tnaA的细菌才能合成吲哚。表达色氨酸酶(tnaA)的细菌,例如大肠杆菌(E. coli)、梭菌属(Clostridium spp.)、拟杆菌属(Bacteroides spp.)和普通变形杆菌(Proteus Vulgaris),可催化色氨酸直接转化为吲哚。
✦ 梭状芽孢杆菌代谢产生吲哚-3-丙酸
已经证明吲哚-3-丙酸是由梭状芽孢杆菌(Clostridium sporogenes)产生的。粪臭素(3-甲基吲哚)是由梭菌属和拟杆菌属产生的。
此外,产孢梭菌(C. sporogene)可以将色氨酸转化为吲哚-3-丙酮酸(IPyA),并在吲哚-3-丙酮酸脱羧酶的催化下产生吲哚-3-乙醛(IAAld)。然后,IAAld 被厚壁菌门、变形菌门、拟杆菌门和放线菌门的成员脱羧,产生吲哚乙酸 (IAA)。
✦ 一些病原菌也可以产生吲哚
色氨酸代谢很复杂,许多菌株参与吲哚衍生物的制造。总的来说,数据支持迄今为止有至少85种细菌已被证明可以产生吲哚。
这些细菌包括许多病原菌,例如创伤弧菌、流感嗜血杆菌、多杀巴氏杆菌(pasteurella multocida)、四种志贺氏菌菌株、植物克雷伯菌(Klebsiella planticola)和普通变形杆菌。值得注意的是,一些革兰氏阳性菌株,包括B.alvei和粪肠球菌(E.faecalis)也产生吲哚。
✦ 一些拥有tnaA基因的细菌却无法产生吲哚
有趣的是,一些细菌,如杀鲑气单胞菌(Aeromonas salmonicida)、宋氏志贺氏菌(Shigella sonnei)、克氏耶尔森氏菌(Yersinia kristensenii)和Seudovibrio spp.,已经失去了合成吲哚的能力,尽管这些菌株的染色体上有一个tnaA基因同源物。
例如,与正常志贺氏菌菌株不同,一些在tna操纵子中具有点突变、插入或缺失的志贺氏菌不产生吲哚,这可能是由于某些适应性优势。虽然只是推测,但一些个体可能通过利用当地细菌群体的信号来避免产生吲哚的成本。
其他可以产生吲哚的细菌:
Alkaliphilus metalliredigens
Bacillus alvei
Clostridium novyi
Clostridium limosum
Clostridium tetani
Corynebacterium acnes
Desulfitobacterium hafniense
Nocardioidessp.
Oribacterium sinus
Propionibacterium acnes
Enterococcus faecalis
Symbiobacterium thermophilum
Aeromonas hydrophila
Aeromonas liquefaciens
Aeromonas punctata
Bacteroides thetaiotaomicron
Bacteroides sp.
Brachyspira hyodysenteriae
Burkholderiasp.
Chromobacterium violaceum
Chryseobacterium gleum
Citrobactersp.
Citrobacter intermedius
Citrobacter koseri
Desulfovibrio vulgaris
Edwardsiella tarda
Enterobacter liquefaciens
Escherichia coli
Escherichia fergusonii
Flavobacteria bacterium
Flavobacteriumsp.
Fusobacterium nucleatum
Fusobacterium polymorphum
Haemophilus influenzae
Haemophilus somnus
Haloarcula marismortui
Halogeometricum borinquense
Hyphomonas neptunium
Klebsiella ornithinolytica
Klebsiella planticola
Micrococcus aerogenes
Morganella morganii
Paracolobactrum coliforme
Pasturella multocida
Pasturella pneumotropica
Photobacterium harveyi
Porphyromonas asaccharolytica
Porphyromonas endodontalis
Porphyromonas gingivalis
Prevotella intermedia
Proteus inconstans
Proteus vulgaris
Providencia alcalifaciens
Providencia rettgeri
Providencia rustigianii
Providencia stuartii
Rhizobium leguminosarumbv.trifolii
Saccharomonospora viridis
Shigella boydii
Shigella dysenteriae
Shigella flexneri
Sphaerophorus varius
Stigmatella aurantiaca
Treponema denticola
Vibrio alginolyticus
Vibrio cholerae
Vibrio fluvialis
Vibrio harveyi
Vibrio hollisae
Vibrio mediterranei
Vibrio nigripulchritudo
Vibrio orientalis
Vibrio parahaemolyticus
Vibrio splendidus
Vibrio tapetis
Vibrio tubiashii
Yersinia frederiksenii
Yersinia intermedia
吲哚类物质的生成可能受到环境因素的严重影响,例如膳食成分、肠道微生物群、碳源、温度、pH值以及生理状态。
★ 肠道微生物群和饮食对吲哚生成有重要影响
吲哚类物质是肠道微生物群产生的膳食色氨酸的代谢产物。因此,肠道微生物组成以及饮食模式的变化会影响其产生。
研究发现吲哚-3-丙酸(IPA)与肠道微生物群的α多样性呈正相关,肠道微生物组的组成可以解释大约20%的IPA循环水平的变化。
高纤维和富含多酚的饮食可以提高吲哚含量
多项研究发现,吲哚-3-丙酸的循环水平与纤维摄入量相关,这可能是通过肠道微生物群的变化来解释的。
此外,富含多酚的饮食会导致肾功能正常受试者的血清IPA显著增加,但肾功能受损的受试者不会显著增加。与此相一致的是,饲喂菊粉60天的猪的盲肠和结肠中微生物组组成的β多样性显著增加,同时IPA含量也显著增加。
地中海饮食会导致吲哚-3-丙酸水平升高
不同的饮食结构也会改变吲哚的水平。例如,地中海饮食仅治疗4天后就导致吲哚-3-丙酸水平升高,而快餐(即薯条和汉堡)则引起相反的结果。
此外,正常饮食喂养的同窝小鼠的血浆IPA水平显著高于生酮饮食喂养的小鼠。一项针对117名超重成年人的研究发现,摄入炸肉会降低肠道微生物群落的丰富度,并导致IPA浓度下降。
发酵食品可能会降低吲哚含量
通过对14名健康男性进行随机交叉研究设计,探讨了发酵乳制品对人类血清代谢组的影响。与牛奶组相比,酸奶摄入组餐后血中吲哚-3-丙酸和吲哚乙酸浓度较低。
同样,C反应蛋白水平轻度升高的健康超重男性在接受抗炎膳食混合物后,饮用500mL餐后奶昔(由300mL蛋奶冻、150mL奶油干酪和50mL鲜奶油组成)。受试者在后期表现出吲哚-3-丙酸血浆浓度降低。
总之,我们推测不同的饮食结构会改变肠道微生物的组成,从而导致吲哚类物质产生的个体差异。
• 细胞群体密度影响吲哚分泌
细胞外吲哚浓度取决于细胞群体密度,其中大肠杆菌和霍乱弧菌在指数早期开始产生吲哚。浓度在固定相中达到最高水平(在富集培养基中高达0.6mM吲哚),并在固定相期间稳定维持。
• 碳源影响吲哚产量
其次,有报道称葡萄糖抑制吲哚生物合成。TnaA的分解代谢抑制得到证实,因为tnaA基因的转录在碳限制期间受到抑制。
此外,tnaAB操纵子被大肠杆菌中的分解代谢调节蛋白环AMP复合物激活。因此,当大肠杆菌数量较多且碳源减少时,会产生相对大量的吲哚。
• 温度和pH影响吲哚生物合成
温度和pH也是影响大肠杆菌吲哚生物合成的重要环境因素。温度从30°C转变到43°C会诱导大肠杆菌中tnaAB基因的表达,但大肠杆菌在44.5°C时失去吲哚生物合成的能力。
注:与37°C相比,吲哚信号传导在较低温度(30°C)下对大肠杆菌基因表达、生物膜形成和抗生素耐药性的控制作用更为显著。
此外,低pH值会抑制大肠杆菌中吲哚的产生,而TnaA是pH9.0时最容易诱导的蛋白质之一。
• 生理状态影响吲哚生成
例如年龄、性别以及个体的生理状态都可能影响内源性吲哚的产生。睡眠模式、压力水平和运动习惯等生活方式因素都可能对内源性吲哚的产生产生影响。
此外,特定的健康状况和疾病,如肠道炎症、肝脏疾病等,可能会改变内源性吲哚的生成或代谢。
• 药物的使用也会影响吲哚生成
一些药物可能影响吲哚类物质的生成。例如,一些抗生素和非甾体抗炎药物被发现可以改变肠道微生物群的组成和功能,从而影响吲哚类物质的生成。
此外,一些药物可能直接干扰色氨酸代谢途径,从而影响吲哚类物质的生成。
小结
因此,细胞群密度、碳源、温度和pH等环境条件直接会影响吲哚的浓度;而饮食、药物的使用以及生活方式可能通过影响肠道微生物群或其他器官间接影响吲哚的生成。
肠道屏障的功能与肠道健康密切相关,在健康中发挥着至关重要的作用。肠道屏障将宿主与肠腔中的微生物隔离,并限制微生物和分子从肠腔的运动。
一旦肠道屏障受损,肠道通透性增加,细菌及其产物(如内毒素)将跨膜并激活单核巨噬细胞系统,促进大量炎症因子的产生,如白细胞介素6和肿瘤坏死因子-α(TNF-α),导致慢性微炎症状态。
吲哚对于控制肠道屏障功效至关重要,包括调节炎症和免疫反应、影响上皮通透性、粘液产生和紧密连接(TJ)形成。
▼
✦ 吲哚减轻非甾体抗炎药引起的肠道炎症
研究了使用吲哚作为非甾体抗炎药肠病的治疗方法。吲哚降低粪便钙卫蛋白浓度以及吲哚美辛诱导的脾和肠系膜淋巴结中中性粒细胞的浸润。
吲哚与吲哚美辛联合使用可减少非甾体抗炎诱导的粘膜转录组变化。粪便钙卫蛋白是肠道炎症的中性粒细胞特异性生物标志物。而中性粒细胞被认为是非甾体抗炎肠病发病的关键因素,可通过释放多种炎症介质引起炎症和组织损伤。
这就说明吲哚能够减轻吲哚美辛引起的小鼠肠道炎症,对非甾体抗炎药肠病具有潜在的免疫调节作用。
此外还发现吲哚减少促炎细胞因子IL-8的产生和趋化因子TNF-α激活的核因子κB(NF-κB)的表达,并诱导抗炎细胞因子IL-10的分泌。
研究表明,共生菌可以限制鼠伤寒沙门氏菌诱导的NF-κB激活,减轻小鼠炎症。因此,吲哚可能是益生菌减少肠道炎症的信号。
✦ 细菌产生的吲哚衍生物增强免疫屏障
吲哚乙酸
卵形拟杆菌(Bacteroides ovatus)产生的吲哚乙酸(IAA)增加了CD103+ /CD11c+免疫群体的丰度,而 CD103+ /CD11c+免疫细胞对于维持肠道免疫稳态和诱导耐受性免疫反应至关重要。
此外,吲哚乙酸与树突状细胞上的芳基氢受体结合并驱动IL-22的产生。在炎症性肠病个体中,IL-22 刺激上皮再生并减少炎症,并且在肠道炎症的调节中发挥关键作用。
注:除卵形拟杆菌外,其他拟杆菌产生的吲哚也可以激活免疫细胞上的AhR,从而产生IL-22。
在脂多糖存在的情况下,肠上皮细胞中表达的Reg4通过增加乳酸菌及其代谢物吲哚乙酸的比例来维持免疫稳态,从而促进肠组织中IL-35 +B调节细胞的产生和积累。这些结果表明吲哚乙酸可以调节IL-35 的产生,并影响肠道免疫屏障。
吲哚-3-乳酸
罗伊氏乳杆菌(Lactobacillus reuteri)产生吲哚-3-乳酸(ILA),ILA是Trp的吲哚衍生物,可激活CD4+ T细胞中的AhR并下调转录因子ThpoK,使CD4 + T细胞分化为具有免疫调节功能的CD4+ CD8αα +双阳性上皮内T淋巴细胞(DPIELs)。通过这些机制,吲哚-3-乳酸可以促进肠道屏障功能并减少炎症。
生活在母乳喂养婴儿胃肠道中的婴儿长双歧杆菌亚种也可产生吲哚-3-乳酸,从而显著减弱TNF-α和脂多糖诱导的肠上皮细胞中促炎细胞因子IL-8的增加。吲哚-3-乳酸还增加AhR靶基因CYP1A1和NRF2靶向基因谷胱甘肽还原酶2、超氧化物歧化酶2和NAD(P) H脱氢酶的 mRNA 表达,这可能是母乳喂养婴儿肠道炎症的重要调节剂。
此外,另一项研究表明,婴儿双歧杆菌分泌的吲哚-3-乳酸对未成熟肠道具有抗炎作用。这些数据为生产吲哚-3-乳酸益生菌的生产和饮食建议提供了重要的见解。
吲哚-3-丙酸
吲哚-3-丙酸(IPA)下调肠上皮细胞介导的炎症细胞因子TNF-α,同时上调配体蛋白编码mRNA,从而调节肠道屏障功能,缓解肠道炎症。
吲哚-3-丙酸还通过激活AhR显著诱导肠上皮细胞上的IL-10受体配体结合亚基(IL-10R1)来调节肠道免疫稳态。
IL-10R1是抗炎细胞因子IL-10的受体,IL-10通过IL-10R1发送抗炎信号,抑制多种细胞(包括肠上皮细胞)过度释放促炎介质。这进一步说明了吲哚-3-丙酸对肠道免疫屏障的保护作用。
吲哚-3-醛
乳杆菌(Lactobacillus)产生的吲哚-3-醛(IAld)有助于AhR依赖性IL22转录,IL-22产生者抑制炎症并保护粘膜表面的免疫生理学。吲哚-3-醛可以替代益生菌,在感染或化学损伤期间保护和维持粘膜完整性。
吲哚-3-醛可用作菌群处理和肠道菌群失调期间的支持疗法。乳酸菌还通过IAld诱导的AhR刺激LPL分泌IL-22,从而激活pSTAT3通路并抑制IL-1β和TNF-α分泌,以保护粘膜免疫屏障.
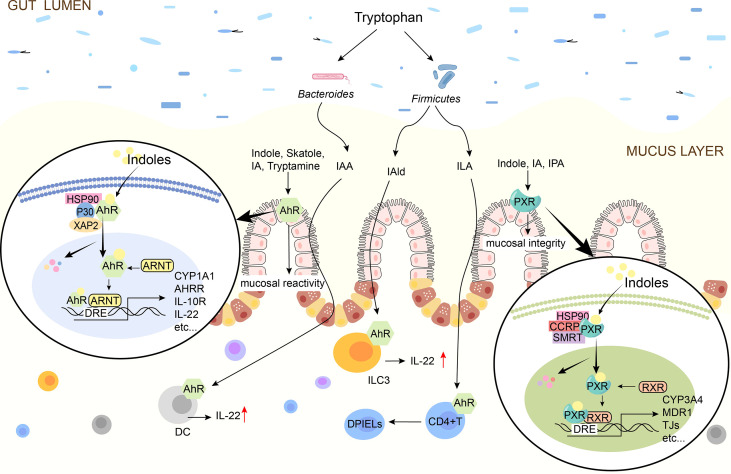
吲哚对肠道免疫的影响
Ye X,et al.Front Immunol.2022
服用非甾体抗炎药时,肠道微生物组以革兰氏阴性菌为主,这种转变会导致肠道损伤。吲哚和吲哚美辛的共同给药可以维持甚至增加厚壁菌门的重要成员,特别是对肠道稳态至关重要的C. cluster XIVa和C. cluster IV。似乎它可以防止拟杆菌的任何增加,这种变化可以减少肠粘膜损伤。
吲哚可通过增殖MUC基因的表达来增加HCT-8细胞对去甲肾上腺素介导的肠出血性大肠杆菌(EHEC)沉降的抵抗力。
吲哚-3-醛(IAld)激活AhR以产生IL-22,IL-22调节粘膜反应,使混合微生物群落存活并抑制白色念珠菌(Candida albicans)定植
✦ 减轻克雷伯菌属的细胞毒性
吲哚调节多种肠道细菌的毒力因子。吲哚增强替利霉素向替利伐林的转化。两者均由克雷伯菌属(Klebsiella spp.)产生,不同之处在于替利伐林是一种细胞毒性降低的吲哚类似物,并且替利伐林与上调孕酮X受体(PXR)反应性解毒基因结合并抑制微管蛋白定向毒性。因此,吲哚以多种方式减轻克雷伯菌属的细胞毒性。
✦ 调节霍乱弧菌和大肠杆菌的毒力基因表达
肠道益生菌代谢产生的吲哚在肠腔内浓度较高,减少了肠道大肠杆菌和啮齿类柠檬酸杆菌(Citrobacter rodentium)致病基因的表达。
吲哚还用作ToxR激动剂,调节肠道内霍乱弧菌(Vibrio cholerae)的毒力基因表达和生物膜产生。吲哚、吲哚甲酸和吲哚乙酸还可下调致病性大肠杆菌菌株的毒力和志贺氏毒素的产生。
此外,吲哚和7-羟基吲哚显著改变铜绿假单胞菌(Pseudomonas aeruginosa)的毒力基因表达,从而降低毒力因子并降低集群运动性。
铜绿假单胞菌用7-羟基吲哚治疗后,胃肠道中有更好的清除率。因此,吲哚可能有助于作为针对肠出血性大肠杆菌等病原体的治疗剂。
✦ 增强肠道机械屏障
吲哚通过诱导许多涉及内皮细胞的基因的表达,包括紧密连接、粘附分子连接(AJ)、肌动蛋白细胞骨架和黏蛋白形成,在体外增强内皮细胞的屏障功能。
将人肠上皮细胞系HCT-8暴露于吲哚以测量其基因表达的变化。结果显示,高剂量的吲哚增强了肠上皮细胞HCT-8中与黏蛋白产生、粘膜屏障增强和上皮间抵抗相关的分子谱。因此,吲哚加强了机械屏障。
✦ 增强肠道粘液屏障
吲哚-3-丙酸通过增加MUC2和MUC4以及杯状细胞分泌产物(TFF3和RELMβ)来增强粘液屏障功能。
吲哚-3-丙酸还促进IL-10释放,IL-10的信号转导增强粘液屏障功能并维持上皮细胞的稳态。
细菌中的吲哚-3-丙烯酸(IA)可增强IL-10的产生和MUC基因的表达,而IL-10对于维持杯状细胞中MUC的产生至关重要。因此,吲哚-3-丙烯酸通过 IL-10间接影响粘液屏障。
吲哚通过AhR或PXR调节肠道屏障
Ye X,et al.Front Immunol.2022
研究发现吲哚在细菌中发挥多种生物学作用,包括孢子形成、耐药性、毒力因子、质粒稳定性和生物膜形成等。
吲哚可以作为细菌间的信号分子
事实上,吲哚可以作为一种细菌信号,参与同一物种内以及不同物种之间的通讯。它在细菌生态系统中充当细胞间和细胞内信号。
注:吲哚还诱导了S. aurantiaca的孢子形成。
通过遗传筛选,在大肠杆菌中研究了吲哚作为细胞外信号分子的可能。结果,四个基因(astD、cysK、gabT和tnaB)被证明在稳定期通过自身产生的细胞外信号的积累而被激活。
在后续研究中,吲哚被证实是astD、gabT和tnaB激活所需的细胞外信号分子。
吲哚作为细胞间信号分子存在哪些争议?
吲哚是否是细胞间信号分子一直存在争议。已经提出了对群体感应信号分子要求的许多标准,如下所示:
1.假定的信号必须在特定阶段产生,吲哚主要在固定相中产生。
2.假定的信号必须在细胞外积累并被特定受体识别。吲哚的化学性质是众所周知的,并且在大多数吲哚调节情况下,化学互补被证明,其中吲哚在稳定期积累并且是已知的细胞外信号。该受体由AcrEF输出并由Mtr输入。
3.假定的信号必须累积并产生一致的响应,吲哚已被证明可以控制孢子和生物膜的形成。
4.重要的是,假定的信号必须引起超出信号代谢或解毒所需的生理变化的反应。吲哚已被证明可以控制毒力、生物膜和质粒稳定性,这些与吲哚无关代谢。
因此,根据这些标准,吲哚有可能成为群体感应分子。
由于上述四个标准仅适合规范的群体感应信号,例如AHL,Monds&O’Toole又添加了两个更普遍适用且具有实用价值的标准:
1.表型变化所需的生理相关信号浓度对细胞无毒。大肠杆菌和霍乱弧菌产生高达0.6mM的吲哚,对细胞无毒,并在该生理浓度下控制许多表型。
2.信号网络在社区层面是自适应的。尽管由于进化实验的困难而难以证明,但我们假设细胞间信号吲哚可能对微生物群落有益,尽管吲哚的生产对个体来说成本高昂。
例如,吲哚在产生吲哚的细菌大肠杆菌和霍乱弧菌中增加了质粒稳定性、耐药性。相反,在不产生吲哚的细菌中,吲哚通过干扰群体感应系统,降低了真菌的细胞生长和铜绿假单胞菌的毒力。因此,我们推测产生吲哚的细菌可能利用吲哚来对抗其他细菌和真核生物。
吲哚和tnaA基因也影响病原菌的毒力。色氨酸酶活性与致病性大肠杆菌有关,因为色氨酸酶活性是LEE1启动子完全激活所必需的。
吲哚会增加肠出血性大肠杆菌中毒力相关的EspA和EspB蛋白(LEE4基因产物)的分泌以及附着和消除病变的形成。
在霍乱弧菌中,吲哚和tnaA基因可能通过诱导毒力相关的分泌蛋白来增加对盘基网柄菌(Dictyostelium discoideum)的抗性。
此外,在流感嗜血杆菌分离株中,大多数血清型 (94-100%)呈吲哚阳性,而无害分离株中只有70-75%呈吲哚阳性。结果表明,吲哚的产生是必要的,但不足以对该菌株产生毒力。
值得注意的是,吲哚通过抑制毒素的产生来减轻克雷伯菌属(Klebsiella spp)的细胞毒性,还通过降低细胞运动性和聚集性来减少李斯特菌(L. monocytogenes)的相关毒力。
吲哚还通过改变基因表达来减少铜绿假单胞菌(不产生吲哚的细菌)中毒力因子的产生。
吲哚降低毒力基因的表达
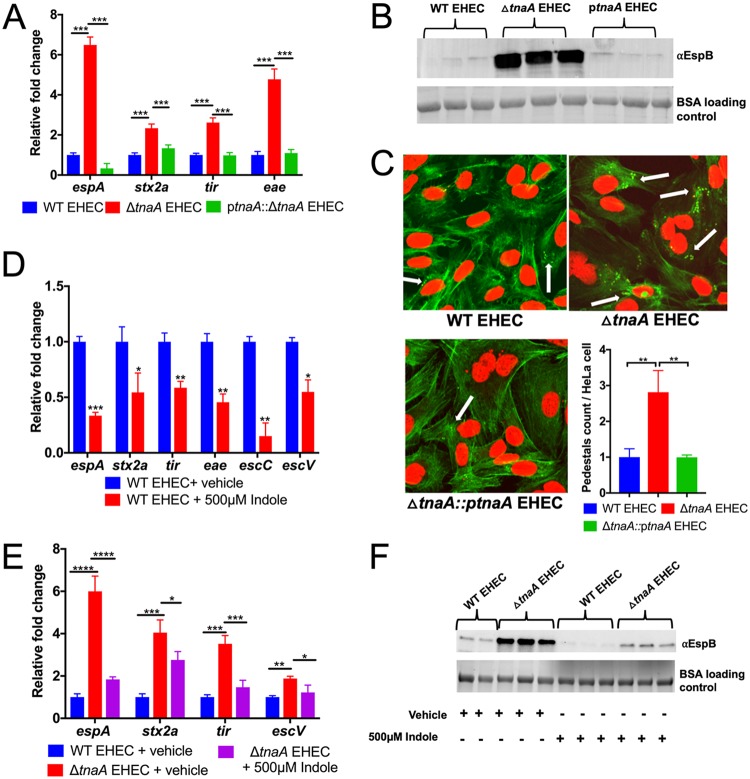
Kumar A,et al.mBio.2019
研究表明,吲哚通过在大肠杆菌中诱导输出基因(mdtEF和acrD)来增加耐药性,其中吲哚通过双组分信号转导系统(BaeSR和CpxAR)发挥作用。
这些双组分信号系统可以用作吲哚传感器。该结果证实了另一项研究,其中吲哚通过BaeSR和CpxAR诱导spy(球形体蛋白Y)基因的表达。
此外,有人认为GadX(AraC型转录因子)、Hfq(sRNA功能的全局调节因子)和RpoS(应激和固定相西格玛S)对吲哚诱导的mdtEF表达是必需的。因此,吲哚可能与多种调节因子相互作用。
增强大肠杆菌的质粒稳定性
吲哚增强大肠杆菌中的质粒稳定性。研究表明,来自大肠杆菌质粒ColE1的小非编码RNA与TnaA结合,有助于防止质粒丢失,而吲哚则延迟细胞分裂。
大肠杆菌中的吲哚还通过抑制gadABCEX、hdeABD和ymgB等耐酸基因来降低耐酸性。大肠杆菌可能会在弱碱性肠道菌群中存在吲哚的情况下关闭耐酸性基因,因为在通过酸性胃存活后不再需要耐酸性蛋白。
降低了运动性
此外,吲哚是一种化学排斥剂,会降低运动性,可能是由于细胞分裂干扰,而肾上腺素和去甲肾上腺素是一种化学引诱剂,会增加大肠杆菌O157: H7的运动性。
此外,吲哚降低细胞对上皮细胞的粘附,而肾上腺素和去甲肾上腺素增加细胞粘附。由于细菌粘附和定植到上皮细胞对于感染很重要,因此假设这也会对细菌毒力产生不同的影响。
吲哚对其他微生物的影响如下所示:

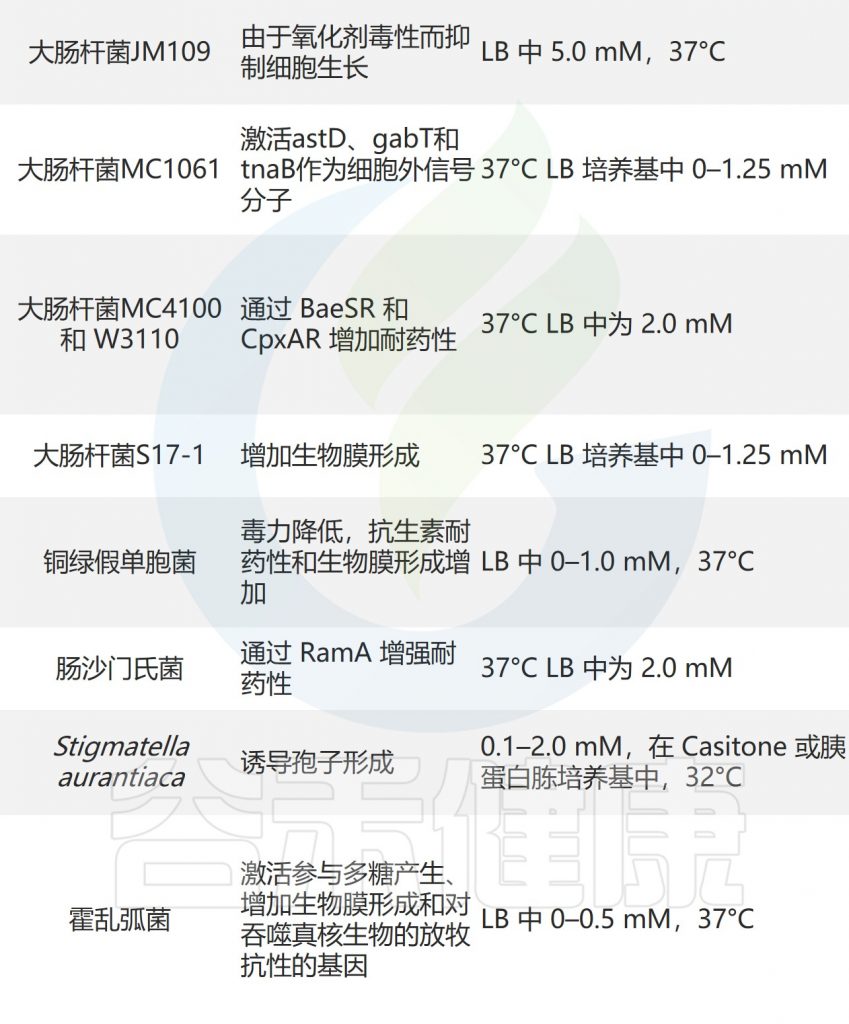
Lee JH, Lee J.FEMS Microbiol Rev.2010
吲哚类物质被肠上皮细胞吸收并扩散到血液中,从而循环到全身并影响各个系统,例如肠道及相关疾病的调节。
谷禾在这里总结了它们对炎症性肠病、出血性结肠炎、结直肠癌、糖尿病、中枢神经系统炎症和血管调节的影响。
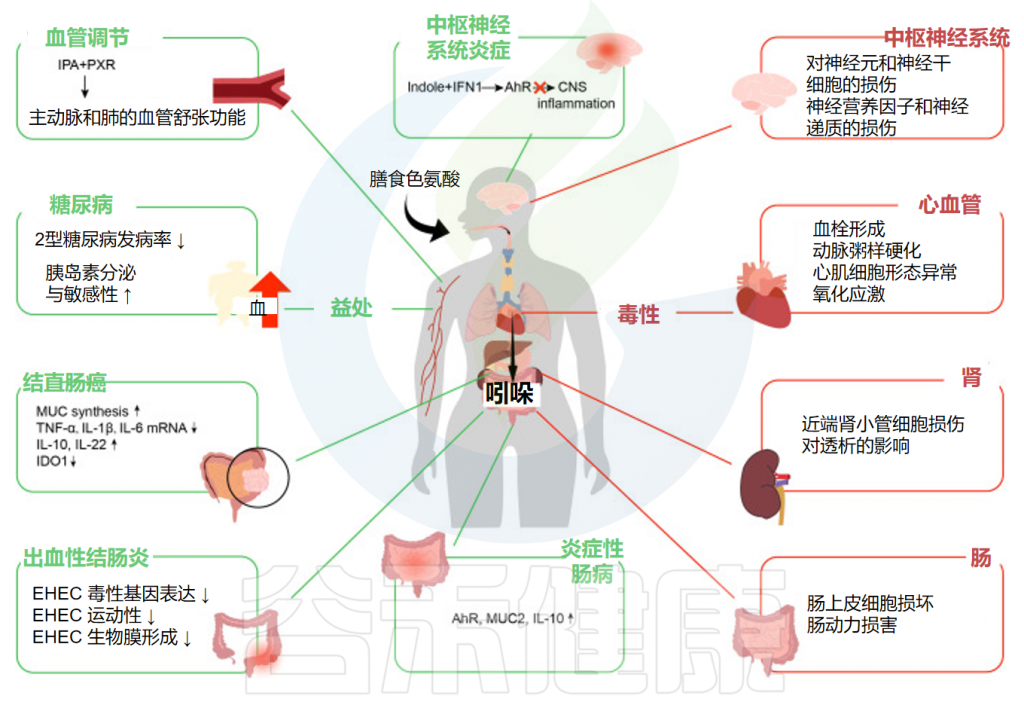
Ye X,et al.Front Immunol.2022
炎症性肠病是克罗恩病、溃疡性结肠炎等一组疾病的总称,是临床上重要的慢性胃肠道炎症性疾病。大量研究表明,吲哚对缓解炎症性肠病症状有一定的作用。
✦ 吲哚类物质增加了抗炎细胞因子
炎症性肠病与粘液层有关,炎症性肠病患者的内部粘液层较薄,MUC2糖基化较低。结果表明,吲哚-3-丙烯酸甲酯(IA)处理小鼠结肠球体可显著增加AhR激活和MUC2基因表达。
即使在脂多糖刺激的共培养物存在下,IA仍能保持其对MUC2基因表达的影响并增加IL-10的产生。杯状细胞产生MUC依赖于IL-10。因此,细菌产生的 IA 有可能增加IL-10的产生和MUC基因的表达,这可能对炎症性肠病患者有利。
吲哚对于肠出血性大肠杆菌的发病机制至关重要。出血性结肠炎是一种出血性腹泻,可导致溶血性尿毒症综合征,由人类病原体大肠杆菌O157:H7引起。
✦ 吲哚减少了肠出血性大肠杆菌的定值
根据表面相关EHEC的DNA微阵列研究,吲哚降低了EHEC中与表面定植和致病性有关的基因表达。
吲哚可以被加氧酶氧化,产生新的种间生物膜信号,从而以多种方式影响相同的表型(生物膜产生)。大肠杆菌K-12和肠道内其他共生菌产生的吲哚可能会限制大肠杆菌K-12生物膜的产生,并通过改变SdiA的表达来降低非致病性大肠杆菌的运动性,从而影响EHEC趋化性和粘附力。粘附测定证实,吲哚可在体外减少EHEC与上皮细胞的附着。
总之,吲哚通过改变肠出血性大肠杆菌毒性基因的表达、减少其运动性和生物膜形成以及减少粘附来减轻肠出血性大肠杆菌感染并改善出血性结肠炎。
根据越来越多的数据,肠道吲哚,特别是色氨酸代谢物,似乎在结直肠癌中发挥着重要作用。吲哚刺激AhR,它可以直接作用于肠道干细胞,维持MUC合成并改善肠道屏障功能。
然而,在炎症相关结直肠癌发生模型中,阻断吲哚-AhR信号通路会显著增加TNF-α、IL-1β和IL-6 mRNA水平。
此外,吲哚衍生物色胺会抑制吲哚胺2,3-双加氧酶(IDO1),该酶参与肿瘤免疫耐受。这些发现表明肠道菌群产生的吲哚可能会减缓结直肠癌的进展。
与健康个体相比,结直肠癌患者的吲哚/色氨酸比率较低,犬尿氨酸(kyn)/色氨酸比率较高。在结直肠癌患者中,犬尿氨酸和IDO1的表达增加,吲哚的产生减少。
吲哚产生的下降削弱了对结肠癌的抑制作用。总体而言,有证据表明微生物色氨酸-吲哚代谢途径的改变在结直肠癌的发病机制中发挥着重要作用。
较高的吲哚-3-丙酸血清浓度可降低患2型糖尿病的可能性,并增强胰岛素分泌和敏感性。
研究发现,与喂食对照饮食的大鼠相比,喂食吲哚-3-丙酸饮食的大鼠的空腹血糖水平显著降低。根据其他研究,吲哚可以调节小鼠结肠成纤维细胞释放胰高血糖素样肽-1(GLP-1)。
✦ 吲哚调节胰高血糖素样肽-1的释放
吲哚在短时间内增加GLP-1的释放,在较长时间内减少其分泌。这些作用结果的产生是因为吲哚可以影响L细胞的两个关键化学途径。
在某种程度上,吲哚可以抑制电压门控K+通道,增加L细胞诱导的动作电位持续时间,并导致Ca2+进入大量增加,从而快速促进GLP-1的分泌。吲哚抑制了NADH脱氢酶,随着时间的推移减缓ATP的生成并降低GLP-1的释放。
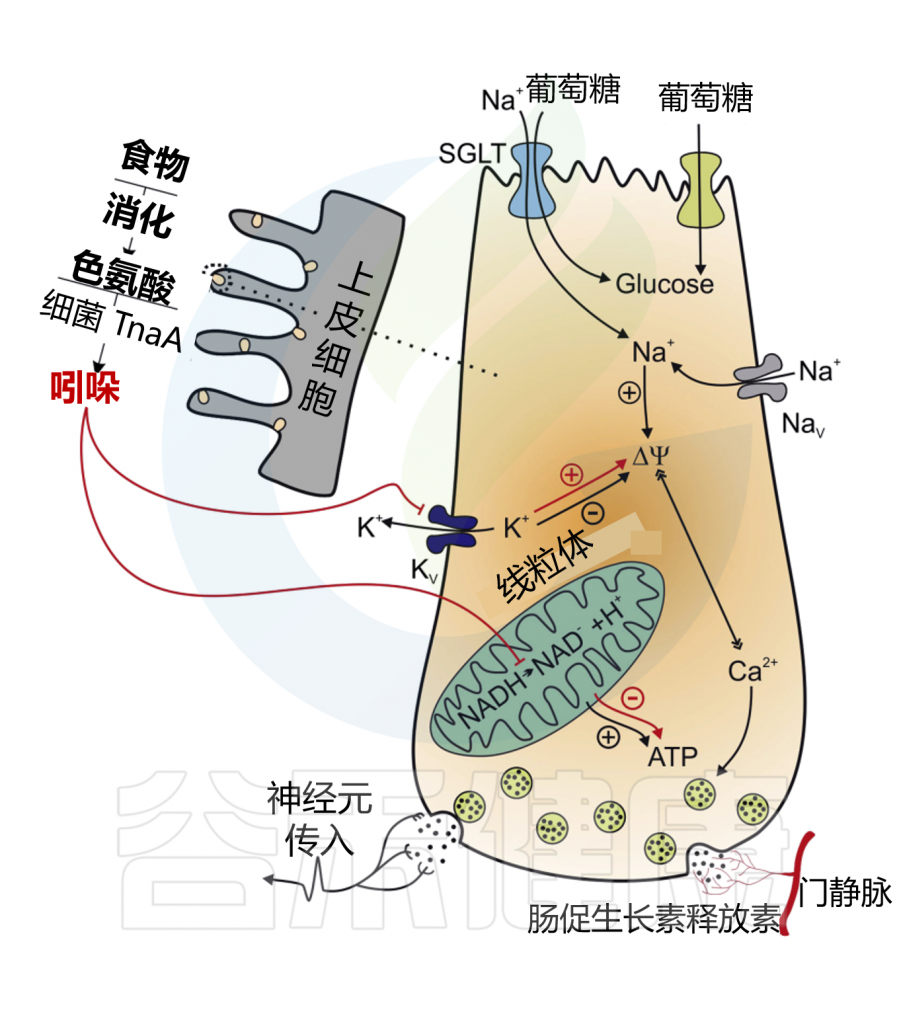
Chimerel C,et al.Cell Rep.2014
肠道内的吲哚可能还会影响食欲,同时,吲哚-3-丙酸具有强大的抵抗氧化应激的能力,这表明这种代谢物可以保护β细胞免受代谢和氧化应激相关的损伤以及淀粉样蛋白积累。因此,肠道微生物群代谢产生的吲哚-3-丙酸对2型糖尿病具有保护作用。
体外实验发现,吲哚丙酸也是嗜肺军团菌(Legionella pneumophila)生长的强效抑制剂,且抑菌效果随着吲哚丙酸浓度和暴露时间的增加而增强。因此,吲哚丙酸可以抑制细菌引起的肺部炎症。
HIV阳性患者通常更容易感染结核病,这是死亡率增加的重要因素,两种疾病的共同感染可能会因抗逆转录病毒药物和抗结核药物之间的相互作用而使治疗复杂化。
值得注意的是,吲哚丙酸可能是此类治疗中潜在的药物成分。有研究发现,接受抗逆转录病毒治疗的HIV患者体内吲哚丙酸含量显著下降,这是区分HIV感染者与健康人的重要标志物。
这种作用的机制尚不清楚,但部分原因是肠道屏障受损导致脂多糖和脂多糖结合蛋白易位,从而加剧全身炎症反应。此外,接受抗逆转录病毒治疗的HIV患者也容易出现非酒精性脂肪性肝病/非酒精性脂肪性肝炎,进一步说明了吲哚丙酸在各种疾病中的多种治疗潜力。
相比之下,吲哚丙酸是一种小分子代谢产物,具有良好的药代动力学特性,很容易被宿主吸收,充分发挥治疗作用。
未来,吲哚丙酸有望通过其作为抗结核病和抗逆转录病毒药物的补充的潜力来改善某些疾病的现有治疗选择。
吲哚丙酸可以触发孕烷X受体(PXR),一种生物量激活的核受体,存在于包括血管内皮在内的各种组织中,以调节内皮功能。
吲哚丙酸通过PXR调节主动脉和肺动脉导管中激动剂诱导的内皮依赖性松弛。这种调节主要是由于内皮一氧化氮合酶产生的一氧化氮的改变,而内皮一氧化氮合酶受到吲哚丙酸介导的PXR激活的抑制。
然而,破坏肠道微生物群并降低吲哚丙酸丰度的抗生素治疗会通过改变内皮PXR途径来改变吲哚丙酸的血管舒张作用。
而微生物代谢的吲哚丙酸补充可以提高全身吲哚丙酸水平并刺激PXR表达,从而逆转抗生素治疗引起的主动脉和肺动脉中激动剂诱导的内皮依赖性血管舒张的增强。
注:吲哚与血管的相互作用可能会显著改变传统传染病或结肠手术相关的抗生素治疗,导致微生物群落紊乱。
尽管吲哚对于改善肠道甚至全身疾病至关重要,但它们中的一些仍然会产生负面影响,例如硫酸吲哚酚(IS)、吲哚基-β-d-葡萄糖苷和吲哚乙酸。
硫酸吲哚酚是最重要的肾毒性代谢产物之一,其肾毒性已在临床研究中得到广泛证实。此外,一些吲哚类物质还会对胃肠系统、神经系统和心血管系统产生毒性。
越来越多的证据表明吲哚和吲哚化合物对大脑代谢、生理和宿主行为有影响。在拥有产生吲哚的大肠杆菌的大鼠中,吲哚的慢性过量产生已被证明会增强焦虑样行为和抑郁。
另一项研究还发现,给予常规大鼠肠道内吲哚可以激活一个称为迷走神经背侧复合体的大脑核。通过比较与非吲哚产生大肠杆菌菌株或吲哚产生大肠菌菌株单体相关的小鼠,发现肠道微生物群的长期高吲哚产生增加了慢性压力对整体情绪行为不利影响的敏感性。
✦ 可能影响儿童自闭症的发病率
在人类中,一项研究发现,患有自闭症谱系障碍的儿童在大便样本中吲哚水平较健康儿童低,而3-甲基吲哚水平升高。 这两种吲哚与患者中更常见的梭菌属细菌有关。
肠道微生物代谢活性产生的血浆吲哚升高与肝性脑病有关,这是由肝功能障碍引起的神经精神障碍,以及个体意识、行为和个性的变化。
此外,NutriNet-Santé观察性前瞻性研究的结果表明,尿液中吲哚和吲哚化合物浓度与反复发作的抑郁症状呈正相关。这种相关性提出了这样一个假设:肠道微生物过度产生这些化合物可能在人类情绪障碍的出现中起到作用。
硫酸吲哚酚由肠道菌群和宿主共同代谢。肠道菌群用酶分解食物中的色氨酸生成吲哚,然后由门静脉携带至肝脏,在细胞色素P450酶和磺基转移酶的作用下转化为硫酸吲哚酚。
硫酸吲哚酚通过上调IRF1表达、抑制动力相关蛋白1表达并干扰线粒体自噬通量来诱导肠上皮细胞(IEC)损伤。硫酸吲哚酚通过以浓度依赖的方式增加活性氧的释放,在IEC-6细胞中引起氧化应激。
硫酸吲哚酚还抑制NRF2激活,降低抗氧化防御细胞系统并抑制血红素加氧酶-1、NAD(P)H 脱氢酶和超氧化物歧化酶的表达。
在IEC-6细胞中,硫酸吲哚酚处理可显著增加TNF-α的释放、环氧合酶-2和诱导型一氧化氮合酶的产生以及硝基酪氨酸的合成,表明肠上皮细胞是硫酸吲哚酚诱导的肠道炎症的靶点。
与硫酸吲哚酚一起培养的结肠显示出收缩性降低,这表明该毒素可能对结肠平滑肌细胞产生有害影响并导致肠道蠕动受损。
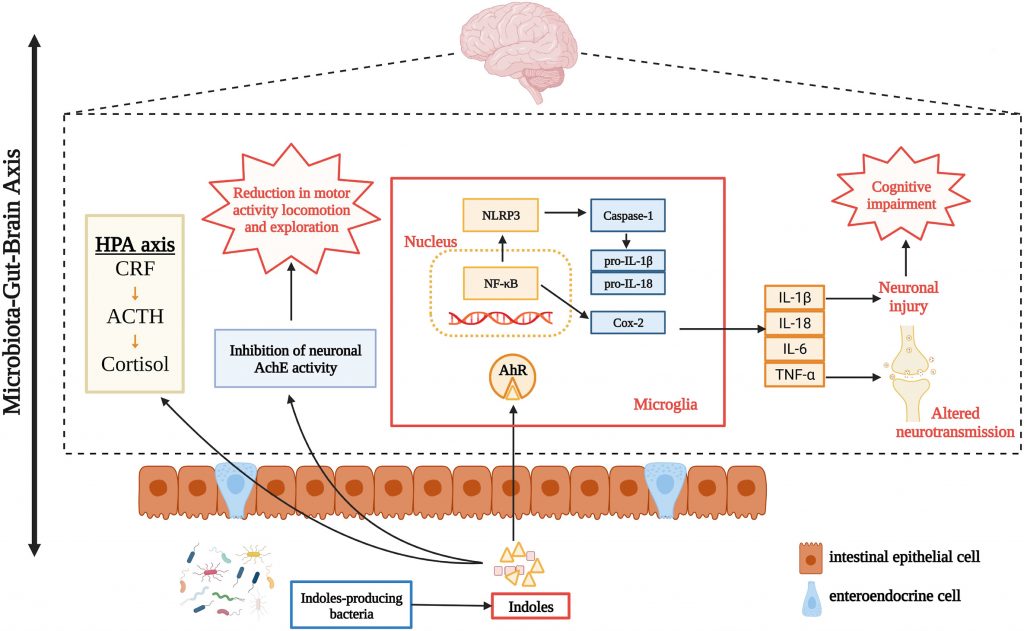
微生物群代谢物硫酸吲哚酚在中枢神经系统疾病的发生中也具有致病作用。众所周知,慢性肾病和心血管疾病常常是由硫酸吲哚酚引起的。此外,肾功能不全会导致尿毒症毒素在大脑中积聚,导致中枢神经系统功能异常。
✦ 损伤神经元和神经干细胞
其次,硫酸吲哚酚可损伤神经元和神经干细胞,损害神经营养因子和神经递质,诱发氧化应激和神经炎症。例如,通过作用于中枢神经系统胶质细胞,硫酸吲哚酚促进神经炎症并表现出促炎作用。
小鼠表现出情绪障碍和神经元退化的行为证据,如焦虑、抑郁和认知功能障碍。这些相应的器质性病变伴随着这些行为变化。这也表明硫酸吲哚酚和中枢神经系统疾病之间似乎存在病理联系。
吲哚对神经和神经精神疾病的影响
编辑
Zhou Y,et al.Neuropharmacology.2023
硫酸吲哚酚对大脑产生有益或有害的影响取决于含量:
通过比较患有抑郁症的志愿者和健康参与者,鉴定出了22种尿液代谢物,并且两组受试者之间的丰度存在差异。患有严重抑郁症的个体尿液中硫酸吲哚酚浓度低于健康人。在志愿者中,血清硫酸吲哚酚浓度与精神焦虑和相关的基于功能磁共振成像的神经特征有关。
在实验性自身免疫性脑脊髓炎的小鼠模型中,每日腹腔注射硫酸吲哚酚(10mg/kg)可调节星形胶质细胞活性,并通过芳基烃受体对中枢神经系统发挥抗炎作用。相反,当大鼠口服较高剂量(100和200mg/kg)的硫酸吲哚酚时,观察到空间记忆受损以及运动和探索活动减少。
另一项研究发现,单次腹腔注射更大剂量(800mg/kg)的硫酸吲哚酚会引起与神经元坏死相一致的大脑组织学变化。硫酸吲哚酚在15-60微摩尔范围内使用时,会诱导原代星形胶质细胞产生自由基氧,并导致海马神经元细胞死亡。在10微摩尔浓度下,硫酸吲哚酚通过氧化应激诱导人星形胶质细胞凋亡。
在慢性肾病(CKD)患者中,硫酸吲哚酚与心血管健康和全因死亡率相关。硫酸吲哚酚是血液透析患者发生心血管疾病的重要因素。在血液透析中,发现游离形式硫酸吲哚酚水平与成纤维细胞生长因子23呈正相关,与C-C基序趋化因子15、补体成分C1q受体、串珠素(Perlecan)、博来霉素水解酶、分化簇166抗原和信号淋巴细胞活化分子家族成员5呈负相关。
这些蛋白质在血管修复和内皮生长中发挥着至关重要的作用。硫酸吲哚酚还可以通过增加血小板过度活跃、提高血浆促凝血水平和产生促凝血颗粒来导致血栓形成和动脉粥样硬化。硫酸吲哚酚通过增加炎症、心脏纤维化、心肌细胞增殖来影响心脏组织。
并且强有力的证据表明,硫酸吲哚酚在肾功能不全的情况下积累时是有害的。硫酸吲哚酚会损害近端肾小管细胞并诱导炎症和纤维化发展。肾脏通过肾小管分泌实现硫酸吲哚酚的高清除率,而硫酸吲哚酚与血浆蛋白结合超过90%,受到蛋白结合的限制,并且血液透析患者的血浆水平相对较高,这也表明与肾脏疾病密切相关。
啮齿动物口服新霉素(一种广谱抗生素)会降低脑内羟吲哚含量,因此表明羟吲哚至少部分源自肠道微生物群代谢活动。
有趣的是,最近在人类粪便样本中发现了羟吲哚,从而证实肠道微生物是这种代谢物的来源。在人类粪便中存在的细菌代谢物中,羟吲哚被发现是主要的芳基烃受体激活剂之一。
大鼠海马切片的体外实验表明,羟吲哚可能与电压门控钠通道相互作用,增加产生动作电位的阈值,从而大大降低神经元的兴奋性。
大胆推测
这些结果说明羟吲哚可能与已知的神经抑制化合物具有一些共同特征,从而能够降低神经元的兴奋性
吲哚及其衍生物可以影响肠道健康甚至是全身其他器官的健康,而过高或过低的水平都可能带来不利影响,导致一些疾病的发生。
调节人体内吲哚水平的机制主要有3种:首先,它可以通过调节与吲哚产生相关的肠道微生物群的丰度来影响吲哚的水平。
先前的研究发现,特定的肠道微生物群成员在色氨酸转化为吲哚的过程中发挥着不可或缺的作用,例如产孢梭菌(Clostridium sporogenes),它已被证明可以通过氧化和还原途径产生吲哚乙酸和吲哚-3-丙酸。在大肠杆菌中也发现了吲哚活性转运蛋白,在乳杆菌中也发现了色氨酸酶的表达,并有助于将色氨酸转化为吲哚。
其次,饮食调整可以增加或减少色氨酸的摄入量,从而间接影响体内的吲哚水平。
最后,直接作用于AHR受体激动剂的药物可能会发挥与吲哚类似的作用。
益生菌可以通过多种机制与体内微生物群相互作用,对宿主健康产生有益影响。
临床试验表明,益生菌可以抵消肠道微生物失衡造成的负面影响,并可导致吲哚产生属及其衍生物的富集,如乳杆菌(Lactobacillus)。在另一项临床试验中,具有抗炎作用的双歧杆菌属在益生菌治疗后的丰度也显著增加。
益生菌还有可能成为神经和精神疾病的治疗辅助药物。在一项针对抑郁症和海马体功能变化的研究中,接受益生菌治疗的小鼠表现出肠道微生物生态失调的显著缓解,以及较低的肠道炎症介质如IL-1β、TNF-α和iNOS的表达水平。此外,情感行为也被检测到与肠道微生物群组成变化相关的显著改善。
益生元是宿主细菌可以选择性利用并提供健康益处的底物。常见的益生元包括低分子量碳水化合物、多酚和多不饱和脂肪酸。
在实验室和临床试验中,益生元(包括菊粉和低聚半乳糖)已被证明可以显著提高产生吲哚的益生菌(例如双歧杆菌和乳酸杆菌)的丰度。在对肠道微生物失衡进行一定纠正后,吲哚作为具有调节作用的配体,对炎症反应具有抑制作用。
合生元是益生菌和益生元的混合物。在大鼠实验中,合生元治疗逆转了肠道微生物特定菌群丰度的变化。
经过合生元(乳杆菌、双歧杆菌、链球菌和菊粉的组合)治疗的患有慢性肾病的大鼠中,肠道中产生吲哚的产气荚膜梭菌的丰度降低至正常水平。此外,合生元治疗可通过改变肠道微生物丰度来有效调节粪便中吲哚及其衍生物的水平。
合生元治疗由于其对吲哚水平的间接影响,可能成为许多神经系统和神经精神疾病的辅助治疗。最近一项针对阿尔茨海默病患者的临床实验表明,在使用合生元后,神经炎症有所减轻。
值得注意的是,合生元干预的改善效果优于单独的益生元或益生菌干预,这表明复合干预的组合更有可能成为临床治疗的实际应用。
粪便微生物移植(FMT)在先前的几项研究中已被证明对缓解胃肠道微生物失调具有不可否认的作用。通过FMT将产生吲哚的益生菌(乳酸杆菌、双歧杆菌等)移植到患者的肠道中,可以改善肠道微生物群。据观察,肠道中产生吲哚的微生物群的丰度增加,吲哚及其衍生物的水平受到间接影响。
吲哚是色氨酸代谢途径上的关键配体,可以调节和抑制神经和精神疾病的炎症。粪菌移植已被证明在各种疾病和临床治疗中对改善患者的肠道生态起着至关重要的作用,它可以通过吲哚在中枢神经系统疾病治疗中间接发挥作用,并被认为是一种潜在的辅助疗法。
肠道微生物通过色氨酸调节吲哚水平。色氨酸是人体必需氨基酸之一,需要完全依赖饮食摄入。
以下是一些富含色氨酸的常见食物:
蛋白质食物:肉类(牛肉、猪肉、鸡肉、火鸡)、鱼类(鳕鱼、鲑鱼、金枪鱼)、贝类(蛤蜊、扇贝)、蛋(鸡蛋、鸭蛋)、豆类(大豆、黑豆、红豆)、奶制品(牛奶、乳制品)等。
蔬菜和水果:菠菜、花椰菜、蘑菇、芝麻、松子、香蕉、菠萝、樱桃、草莓、蓝莓等。
谷物和豆类:燕麦、大麦、小麦、黑米、糙米等。
坚果和种子:核桃、杏仁、腰果、葵花子、南瓜子、亚麻籽等。
研究表明,0.4%膳食色氨酸可显著减少衰老小鼠的神经退行性病变和神经炎症。在此过程中,补充色氨酸可以通过改变肠道微生物组成并增加阿克曼氏菌等有益细菌的丰度来增强肠粘膜屏障功能。肠道菌群产生的吲哚类物质(如吲哚乙酸、吲哚-3-丙酸)显著升高,可以激活AHR,从而抑制神经炎症。
通过血清靶向代谢组学分析和16s rRNA测序对全身色氨酸代谢和肠道微生物组进行了分析,进一步证实了微生物群-肠-脑轴在调节色氨酸饮食产生吲哚的潜在作用。
虽然色氨酸饮食是一种可能的治疗策略,但过多的色氨酸摄入也被证明具有有害作用,应注意适量摄入。
此外,据报道,间歇性禁食还可能通过调节肠道微生物组成来影响血浆吲哚,从而对神经和神经精神疾病的预后产生有益影响。饮食调节可能是改善吲哚含量的一种简单但有效的方法。
最近的综合研究表明,吲哚通过作用于靶细胞来调节炎症反应和细胞生理。近年来,开发吲哚类作用靶点的药物一直是一个重要的研究热点。
吲哚及其衍生物充当AHR的配体,并通过AHR间接调节星形胶质细胞和小胶质细胞的炎症。因此,直接作用于AHR的激动剂也可以作为内源性AHR配体(如吲哚)的替代品,并参与色氨酸AHR介导的炎症抑制途径。
体外合成的AHR配体2-(1′H-吲哚-3′-羰基)-噻唑-4-羧酸甲酯在一项临床研究中被证明具有抑制自身免疫性脑脊髓炎进展的能力。对AHR具有激动作用的药物有可能取代色氨酸代谢产物,并作为外源性AHR配体激活自限制途径和修复神经和神经精神疾病。
除了靶向AHR,影响吲哚作用途径中其他靶点的药物也可能具有治疗作用,这需要在未来进行更多的研究。
由于吲哚是肠道细菌常见的代谢产物,其重要的生物学作用不容忽视。吲哚与肠道菌群的稳态和肠道健康直接相关,吲哚还可以间接影响其他系统和人体的整体健康。
吲哚对人类健康具有有益影响,通过激活免疫细胞释放抗炎因子如IL-22来调节肠道屏障并帮助维持肠道稳态,抑制包括肠出血性大肠杆菌在内的病原菌的定植,以及增加MUC表达以增强粘液屏障功能。
此外,吲哚在调节肠道微生态方面也起着关键作用。它们抑制有害菌株,并以影响基因表达的方式改变肠道致病菌的毒力,这有助于缓解出血性结肠炎等疾病。同时,吲哚及其衍生物在激活AhR和PXR介导的抗炎途径中至关重要。例如吲哚-3-丙酸通过PXR介导肠细胞TNF-α的下调,并上调编码生长抑素的mRNA,以调节炎症中的肠道通透性和肠道屏障功能。这意味着吲哚及其衍生物有前景的治疗途径。
尽管如此,吲哚也是一把双刃剑,少数衍生物也存在一定的缺点。例如,吲哚肝脏代谢产生的硫酸吲哚酚在高浓度时具有肾毒性和心血管毒性,可通过促进氧化应激、炎症等病理变化而导致多系统功能障碍。
吲哚在肠道稳态和人类健康中发挥着至关重要的作用。许多肠道细菌被鉴定为将色氨酸代谢为吲哚。尽管如此,仍可能存在未鉴定的细菌,因此未来需要使用代谢组学和宏基因组学来进一步表征未知的吲哚产生细菌及其相关代谢途径。
不同浓度的吲哚发挥着不同的生理功能。健康成年人的粪便吲哚浓度是已知的,而人类肠道和血液中许多吲哚衍生物的浓度仍然未知。缺乏在不同环境中测定吲哚及其衍生物浓度的定量方法,未来需要开发简单快速的测定方法。
未来仍需进一步研究吲哚与人类健康之间的关系。需要研究多种吲哚组合对宿主生理学的影响,并将这些发现与临床环境相结合,以开发相关疾病的新疗法。
主要参考文献
Zhou Y, Chen Y, He H, Peng M, Zeng M, Sun H. The role of the indoles in microbiota-gut-brain axis and potential therapeutic targets: A focus on human neurological and neuropsychiatric diseases. Neuropharmacology. 2023 Nov 15;239:109690.
Tennoune N, Andriamihaja M, Blachier F. Production of Indole and Indole-Related Compounds by the Intestinal Microbiota and Consequences for the Host: The Good, the Bad, and the Ugly. Microorganisms. 2022 Apr 28;10(5):930.
Ye X, Li H, Anjum K, Zhong X, Miao S, Zheng G, Liu W, Li L. Dual Role of Indoles Derived From Intestinal Microbiota on Human Health. Front Immunol. 2022 Jun 17;13:903526.
Lee JH, Lee J. Indole as an intercellular signal in microbial communities. FEMS Microbiol Rev. 2010 Jul;34(4):426-44.
Jiang H, Chen C, Gao J. Extensive Summary of the Important Roles of Indole Propionic Acid, a Gut Microbial Metabolite in Host Health and Disease. Nutrients. 2022 Dec 28;15(1):151.
Kumar A, Sperandio V. Indole Signaling at the Host-Microbiota-Pathogen Interface. mBio. 2019 Jun 4;10(3):e01031-19.
Chimerel C, Emery E, Summers DK, Keyser U, Gribble FM, Reimann F. Bacterial metabolite indole modulates incretin secretion from intestinal enteroendocrine L cells. Cell Rep. 2014 Nov 20;9(4):1202-8.
Lu Y, Yang W, Qi Z, Gao R, Tong J, Gao T, Zhang Y, Sun A, Zhang S, Ge J. Gut microbe-derived metabolite indole-3-carboxaldehyde alleviates atherosclerosis. Signal Transduct Target Ther. 2023 Oct 4;8(1):378.
谷禾健康
同样暴露在含有致病微生物的环境中,有的人很少生病,有的人却很容易感染并生病。这是因为每个人的机体对潜在的病原体产生的免疫应答不同。
★ 固有免疫和适应性免疫保护着人体
人体的免疫应答分为固有免疫(先天免疫)和适应性免疫(后天免疫)。
固有免疫是生物在长期进化中逐渐形成的,是机体抵御病原体入侵的第一道防线。
参与固有免疫的细胞包括白细胞、单核/巨噬细胞、树突状细胞、粒细胞、自然杀伤细胞等。
适应性免疫,是机体针对外来刺激而产生的免疫反应,包括细胞免疫和体液免疫。其中细胞免疫是指T细胞受到抗原刺激后,增殖、分化、转化为效应T细胞,当相同抗原再次进入机体的细胞中时,效应T细胞对抗原的直接杀伤作用及致敏T细胞所释放的细胞因子的协同杀伤作用,而体液免疫是指B细胞产生抗体来达到保护目的的免疫机制。
无论是先天免疫还是后天免疫,免疫细胞之间的平衡和协调互作对于每个个体应对感染和疾病的发展息息相关。
肠道微生物,病原体感染以及饮食成分在调节和诱导这些免疫细胞形成和平衡中发挥不可替代的作用。
肠道菌群主要分布在胃肠道,尤其结直肠部位,所以从上面说肠道对于免疫的作用不言而喻。而且肠道还有一个重要功能是感知外部信号并做出反应。
肠道具有大量的淋巴组织和大量分散的先天性和适应性免疫细胞。肠道及相关微生物的免疫过程也越来越多地涉及控制身体其他部位的疾病发展。这反应在肠道免疫相关的文章数量的激增。
注意,由于胃肠道沿其长度存在不同的功能和结构差异,每个部分都有不同的生理作用和免疫学成分。
小肠的免疫系统侧重于保护表面上皮细胞消化和吸收食物的能力,使其免受感染。
这些机制包括: 产生IL-17和IL-22的T细胞和先天性淋巴样细胞;抗菌肽的生产;和具有先天和细胞溶解效应功能的上皮内T细胞。调节性T细胞有助于防止对饮食抗原的超敏反应。
大肠(结肠)不参与消化,但它是大量对健康至关重要的共生微生物的储存库。结肠免疫系统将这些微生物识别为潜在的危害,并在不驱逐它们的情况下与它们“保持一定距离”。
这涉及上皮细胞更新,粘液层的产生、IgA 等抗体的产生以及大量调节性T和B细胞的存在。
免疫功能的这些不同方面由不同种类的次级淋巴器官或免疫细胞提供服务,并受到局部环境因素的影响,例如饮食成分和细菌代谢物。这其中肠上皮细胞、肠道黏膜、肠道微生物互作对于肠道免疫和全身健康的重要贡献是普遍得出的共识。
因此,进一步研究肠道微生物群与肠道免疫系统之间的相互作用机制,对于我们深入了解这种复杂关系的本质具有重要意义,并为免疫相关疾病的预防和治疗提供新的思路。
本文主要分享肠道的结构和不同部分的功能,肠道免疫涉及的主要免疫细胞及其来源和能量代谢,还有肠道微生物群在其中起到的重要作用,及其失调与免疫相关疾病的关联。
免疫中涉及的一些专业术语
Th1/Th2轴:Th1和Th2细胞是两种不同类型的辅助T细胞。Th1细胞主要分泌干扰素γ和IL-2等细胞因子,参与细胞免疫应答;而Th2细胞则主要分泌IL-4、IL-5、IL-10和IL-13等细胞因子,参与体液免疫应答。Th1/Th2轴的平衡与否,与多种疾病的发生和发展密切相关。Th2状态下容易诱发过敏性疾病;Th1状态下容易诱发自身免疫性疾病。
Th17/Treg轴:Th17和Treg细胞分别是促炎性和抗炎性的T细胞。Th17细胞主要分泌IL-17等细胞因子,参与炎症反应;而Treg细胞则主要分泌IL-10、TGF-β等细胞因子,参与免疫耐受。Th17/Treg轴的失衡,也与多种疾病的发生和发展密切相关。Th1/Th2和Th17/Treg细胞失衡与肿瘤大小、淋巴结转移和血管浸润有关。
NK/NKT轴和自然杀伤细胞:NK细胞和NKT细胞都是自然杀伤细胞,可以直接杀伤病原体和肿瘤细胞。NK细胞主要通过分泌细胞毒素杀伤靶细胞;而NKT细胞则通过分泌细胞因子和直接杀伤靶细胞等多种方式发挥作用。NK/NKT轴的失衡,也与多种肿瘤,炎症,感染等有关。
DC/T轴:DC细胞是树突状细胞,是免疫系统中最重要的抗原递呈细胞。T细胞则是免疫系统中最重要的效应细胞。DC/T轴指的是DC细胞和T细胞之间的相互作用和调节关系。DC细胞通过递呈抗原,激活和引导T细胞的免疫应答,从而对抗病原体和肿瘤细胞。
目录
第一部分:肠的解剖学和生理学
①肠道的结构
②小肠具体的结构和功能
③大肠具体的结构和功能
第二部分:肠道内丰富的免疫细胞
①肠上皮细胞——肠道免疫系统的协调员
②巨噬细胞——肠道免疫系统的关键哨兵
③先天淋巴细胞——肠道免疫的重要力量
④肠神经细胞——中心调节者
⑤肠潘氏细胞——肠道守护者,控制易感性
⑥肠杯状细胞——肠道中的前哨和护送者
⑦T细胞——免疫强弱,免疫监视
⑧B细胞——免疫耐受,产生抗体
第三部分:肠道微生物群与免疫反应
①肠是共生微生物的主要来源
②肠道微生物群——免疫系统的调控者
③肠道菌群失调与免疫相关疾病
▼
肠道是从口腔延伸至肛门的长管状结构,其主要功能是消化食物、吸收营养和水分以及排泄废物。肠道也是许多病原体的主要进入部位,并且是庞大而多样化的微生物群落家园。
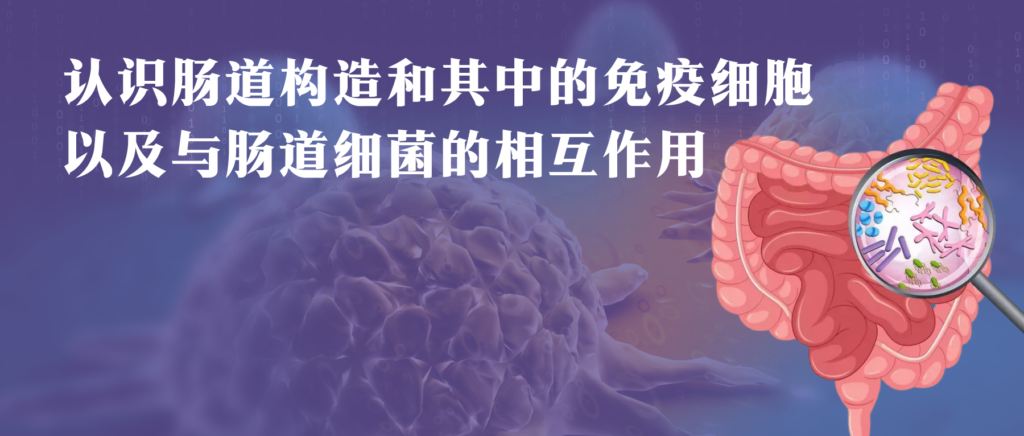
小肠和大肠形成一个连续的管子。小肠始于幽门,止于回盲瓣,回盲瓣是大肠的入口。小肠分为三个主要部分,十二指肠最靠近胃,然后是空肠,最后是回肠。大肠始于盲肠,然后是升(近端)结肠、横结肠、降(远端)结肠和直肠,止于肛门。
小肠和大肠的大小明显不同,人类的小肠由多个长度为6-7米的线圈组成;结肠直径更宽且更短(大约1.5m)。
肠粘膜及其免疫装置的解剖结构
Mowat AM,et al.Nat Rev Immunol.2014
在解剖学上,肠组织通常可分为肠系膜、浆膜、肌层、粘膜下层、固有层、上皮和管腔。
▸ 固有层中有众多免疫细胞
在固有层中发现了许多适应性和先天免疫细胞,包括巨噬细胞、肥大细胞、嗜酸性粒细胞、树突状细胞、B细胞和T细胞。
此外,血管、神经、神经胶质突起延伸至整个固有层和粘膜,与这些结构相关的多种细胞和分子对于肠道功能很重要。
▸ 浆膜包裹并润滑胃肠道
肠系膜与浆膜相邻,浆膜是间皮的最外层,包裹并润滑胃肠道,使蠕动收缩不受抑制。
▸ 肌层是肠组织的最外层
肠组织的最外层统称为肌层。该区域由外部纵向肌肉层和内部圆形肌肉层组成,这些层彼此交织,拉伸。
下面详细描述的结构和细胞区室化将为胃肠道中可能发生的大量神经免疫相互作用提供解剖学参考。与这些结构相关的细胞和分子对于肠道功能非常重要。
▼
✦小肠的主要功能是消化和吸收
小肠的主要功能是营养物质的消化和吸收,起始于胃的幽门,终止于回盲瓣。在人类中,它的长度达到6-7m,按降序由十二指肠、空肠和回肠组成。
小肠的特征是绒毛的指状突起,它延伸到内腔并增加消化活性上皮的表面积,绒毛的丰富表面积,使得它们能够更充分地接触到食物中的营养分子,并将其迅速吸收进入血液循环。
相比之下,表面平坦的盲肠和结肠没有绒毛。
•十二指肠
十二指肠位于胃和空肠之间,是食物从胃进入小肠的第一段。主要功能包括:
•接受来自胃的部分消化食物,继续消化和吸收。
•分泌胰液和胆汁,帮助消化食物中的脂肪和蛋白质。
•吸收营养物质(如碳水化合物、脂肪和部分氨基酸)。
除了来自胃的管腔内容物外,十二指肠每天还通过胆总管接收0.6-1.0L胆汁。胆汁的主要成分包括胆固醇、卵磷脂、胆红素和胆汁盐,后者对食物中的脂肪颗粒具有关键的洗涤作用,并有助于吸附脂肪酸、单甘油酯、胆固醇和其他脂质。
•空肠
空肠位于十二指肠之后,是小肠的一段。主要功能包括:
•进一步消化和吸收食物中未消化的碳水化合物、脂肪和蛋白质。
•吸收维生素、矿物质和水分。
•回肠
回肠是消化系统的一部分,位于空肠之后,是小肠中最后一个部分。它具有以下主要功能:
•营养物质吸收:回肠是主要的吸收器官之一,通过其表面上的细胞吸收并转运剩余的营养物质。
•液体平衡维持:回肠通过吸收水分和电解质维持体内的液体平衡。
•消化废物储存:回肠也起到储存和排泄消化废物的作用。在回肠中,食物残渣逐渐形成粪便,然后被推进结肠,进行最终的排泄。
单糖、二糖和三糖、氨基酸、膳食脂肪、脂溶性维生素(维生素A、D、E和K)或水溶性维生素(维生素B和C)主要存在于十二指肠和空肠中,而回肠是吸收胆汁盐和维生素B12的主要场所。
小肠上段的损伤(如乳糜泻中所见)会导致严重的吸收不良、蛋白质渗漏和营养不良。
✦小肠中有众多的免疫细胞
大多数免疫过程发生在粘膜中,粘膜包括上皮细胞、下面的固有层和粘膜肌层。固有层由松散堆积的结缔组织组成,形成绒毛的支架,并包含粘膜的血液供应、淋巴引流和神经供应。它还包含许多先天性和适应性免疫系统的细胞。
淋巴细胞也存在于上皮细胞中,尽管仅被一层薄的基底膜隔开,固有层和上皮细胞形成非常不同的免疫隔室。它们的组成和功能在整个肠道中也有很大差异。
✦与小肠相关的疾病
乳糜泻
一种自身免疫性疾病,食用麸质会引发白细胞攻击小肠内壁。
克罗恩病
一种导致消化道疼痛性炎症的长期疾病。
肠易激综合症
一种与大脑和肠道之间的信号传导问题相关的常见病症,可导致胃肠道症状。
小肠细菌过度生长
小肠内菌群数量或菌群种类改变,达到一定程度并引起临床表现者。临床表现包括腹痛、腹胀、腹泻或是吸收不良。
消化性溃疡
是位于胃或十二指肠的开放性溃疡。
小肠憩室
肠腔内压力的影响或胚胎时期发育不良使小肠壁薄弱处向外膨出形成的盲囊。
▼
大肠是人体消化系统中的重要组成部分,主要分为盲肠、结肠和直肠。
•盲肠
盲肠是一个封闭的囊,是大肠的第一部分,长约6~8cm,位于腹腔右下部,与回肠交接处有回盲瓣,有孔与阑尾相连,向上续接升结肠。
它充当共生细菌的大储藏室,这些共生细菌参与无法被小肠酶处理的复杂碳水化合物,蛋白质等的发酵消化。
盲肠没有绒毛,粘膜主要由隐窝组成,只有短的平面上皮区域。杯状细胞数量众多,遍布整个隐窝。潘氏细胞很少见。盲肠是整个结肠中最宽的部分,大约20%的结肠癌发生在盲肠。
•结肠
结肠构成了大肠的大部分,包括以下四个部分:
升结肠(从盲肠向上,位于腹部右下角)
横结肠(穿过腹部)
降结肠(向下到最后的结肠部分)
乙状结肠(结肠的最后部分)
升结肠
升结肠长约10厘米,起始于盲肠的肝曲。它沿着腹部右侧延伸,连接横结肠。
横结肠
横结肠长约50厘米,靠近胃、肝脏和胆囊。它连接到降结肠,即所谓的大肠远端或左侧部分的起点。
降结肠
降结肠长约10厘米,结肠在腺曲转折向下固定在腹后壁左侧的一段为降结肠。降结肠下降至左髂嵴处与乙状结肠相连。
乙状结肠
结肠的一部分,在左髂嵴平面与降结肠相连,至第三骶椎平面续于直肠,位于左腹下部及小骨盆内,呈“乙”状或“S”形弯曲,长约40cm。
•直肠
直肠是人体消化系统中的最后一部分,位于盆腔,连接着乙状结肠和肛门。
直肠的壁内有众多的褶皱和粘膜绒毛,这增加了其表面积,有助于吸收水分和电解质。这些特点有助于保持粪便的适当湿润,以便于排泄。
!
注意
人类结肠比小肠短得多(约1.5m)且宽。尽管也被单层柱状上皮细胞覆盖,但其表面完全由散布在表面上皮平坦区域之间的隐窝组成。产生粘液的杯状细胞在小肠上皮细胞中所占比例相对较小(≤10%),但在大肠上皮细胞中所占比例≥25%。
小肠与结肠中的主要细胞与结构
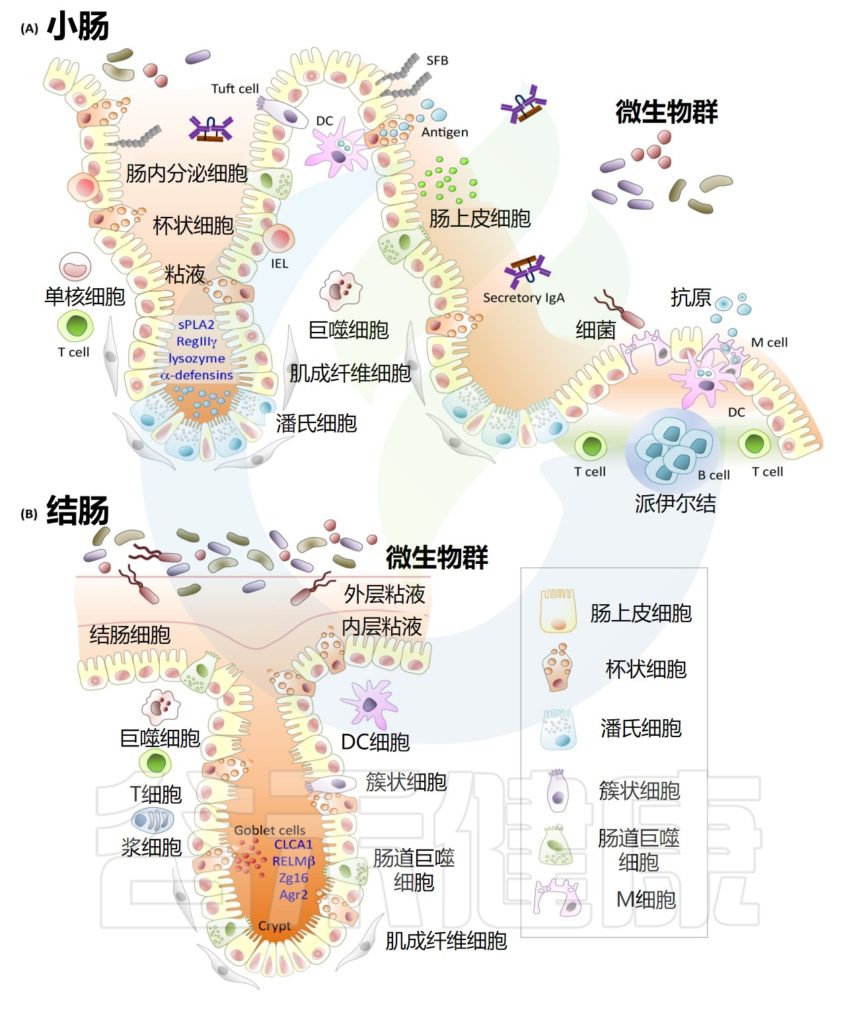
Allaire JM,et al.Trends Immunol.2018
尽管大部分膳食成分在小肠中被吸收,但每天仍有约1.5L的肠液通过回盲瓣。这种液体主要由水和电解质组成,以及半消化或未消化的食物,包括植物多糖和纤维。其中大部分被结肠上半部吸收,它们是常驻微生物群的重要营养来源。
所有结肠癌中约20%至25%起源于远端结肠,而25%至30%的癌症发生在长约15厘米的直肠。
✦与大肠相关的疾病
结肠癌
结肠癌可以始于升结肠。由于它并不总是会引起症状,因此通常直到癌症晚期才被发现。
注:当诊断出结肠癌时,病理学家将确定癌症已到达哪一层(例如最内层或最外层),以帮助确定癌症的分期。
结肠息肉
结肠息肉是发生在肠内壁上的茎状生长物。它们伸入结肠内部。如果它们变大,就会阻止废物通过。
注:息肉是癌症的先兆,在筛查结肠镜检查过程中,发现的任何息肉都将被切除,这样它们就不会继续生长并带来患癌症的风险。
憩室病
憩室病是一种在结肠内壁薄弱部分出现的向外囊状突起。它可能不会引起任何症状,除非被感染,这被称为憩室炎。憩室炎可能导致发烧、恶心、腹痛和腹泻。
注:对于40岁以上的人来说,结肠内壁出现这些小袋很常见,大多数50岁以上的人也有这种情况。
炎症性肠病
克罗恩病、溃疡性结肠炎和不确定性结肠炎是炎症性肠病的一种。炎症性肠病会引起消化道炎症和溃疡。这可能会导致消化道出血、腹痛和腹泻。
此外,炎症性肠病还可引起肠外并发症,包括疲劳、皮肤病、眼部问题和各种形式的关节炎。
▼
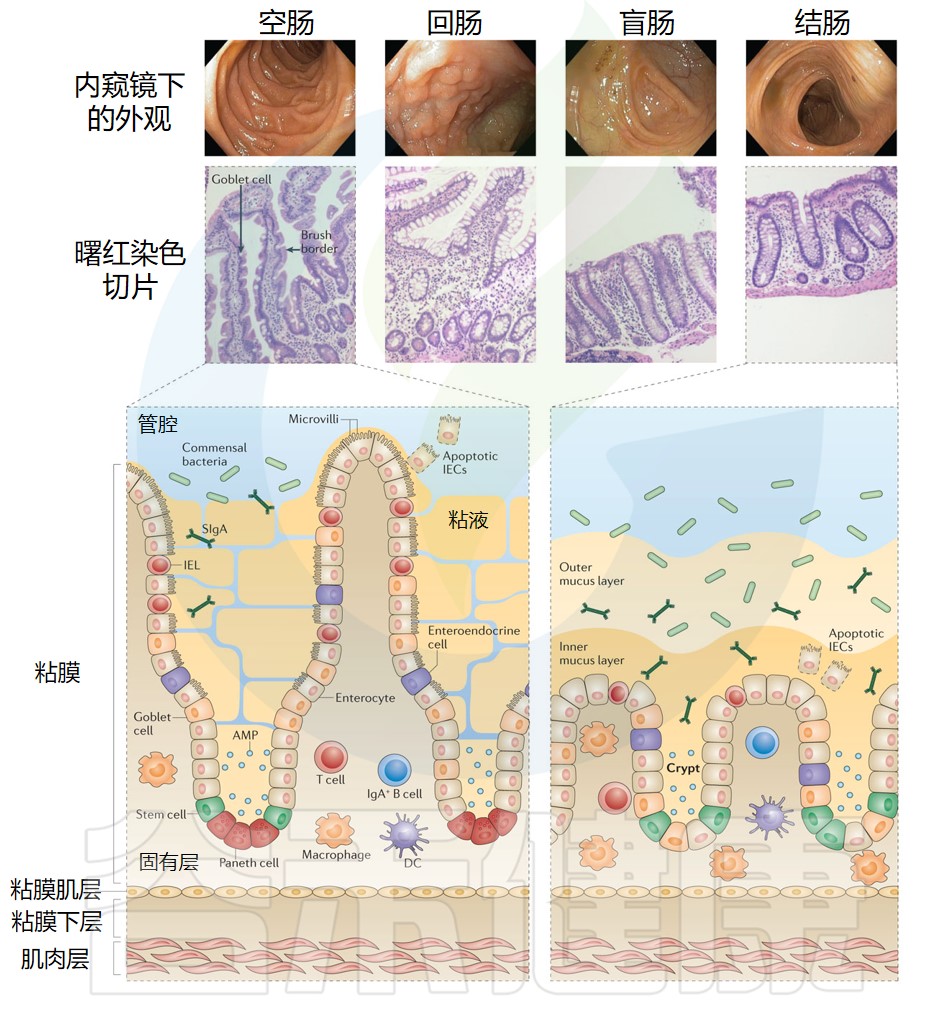
肠上皮细胞是位于肠道内壁的特殊细胞类型,它们构成了肠道黏膜的外层。肠上皮细胞具有多种重要功能,包括吸收营养物质、分泌消化酶、阻止有害物质进入身体内部以及促进肠道免疫响应等。
√大肠与小肠间肠上皮细胞差异较大
肠上皮是单层细胞,但是在小肠和大肠之间的结构和细胞组成上有很大差异。
在小肠中,上皮延伸到伸入管腔的结构,称为绒毛,从而增加粘膜表面积和营养吸收。结肠中没有绒毛,导致粘膜表面相对平坦,从而限制了半固体粪便通过大肠造成的潜在损害。
√肠上皮细胞内具有不同分化类型的细胞
上皮本身并不是一层直的细胞,而是由被称为“Lieberkühn 隐窝”的内陷组成。肠干细胞位于这些隐窝的底部,并产生短暂的增殖细胞,这些细胞在向上穿过过渡区时分化和成熟,肠上皮细胞最终脱落到隐窝顶端的管腔(或小肠中的绒毛)。
注:肠隐窝经历肠上皮细胞补充和更新的持续循环,在稳态条件下,估计整个隐窝每4-5天更换一次。
在肠道上皮细胞内发现了各种分化的细胞类型,每一种都执行独特和专门的功能。这些细胞类型包括:肠细胞,肠上皮细胞中负责营养和水分吸收的最主要细胞类型;各种分泌细胞,例如分泌粘蛋白的杯状细胞,分泌激素的肠内分泌细胞,以及释放抗菌因子以促进消化的潘氏细胞,保护小肠隐窝底部附近的干细胞。
还有在抵御蠕虫中起关键作用的化学感受簇细胞,以及对免疫系统摄取和最终呈递管腔抗原不可或缺的M细胞。
√肠上皮细胞在免疫中主要起屏障功能
肠上皮细胞对免疫的主要贡献是屏障功能,可防止肠腔物质与肠道内壁细胞之间的相互作用。除了不渗透性、快速更新和蠕动等因素外,上皮层首先受到杯状细胞产生的粘液的保护,其次受到无数抗菌分子的保护,例如隐菌素、溶菌酶、磷脂酶和趋化因子。
这些天然杀菌或抑菌化合物会损害多种微生物的增殖,从而阻止入侵粘膜之前大部分正在进行的定植。
尽管在大多数情况下先天免疫足以保护肠道,但适应性免疫系统效应淋巴细胞的记忆特性使病原体在再次遇到病原体时能够更特异、更有效地被消除。
因此,消化道是免疫记忆的主要部位,并且确实构成了人体B和T淋巴细胞的主要储存库。已有文献总结了在解剖学上不同的粘膜诱导位点和粘膜效应位点引发适应性免疫。抗原特异性反应的先决条件是通过粘膜上皮取样抗原并递送至下面的MALT或引流粘膜的淋巴结。
上皮细胞的能量代谢
上皮细胞的能量代谢,尤其是结肠中的能量代谢,在很大程度上取决于肠道微生物群。在生命早期,在适应性免疫系统成熟之前,未知的微生物衍生分子通过 STAT3 磷酸化以IL-23和IL-22依赖性方式激活上皮内淋巴细胞和先天淋巴细胞。
在缺乏适应性免疫的情况下,IL-23-IL-22可以控制肠道微生物群,但过度激活的IL-22产生会导致脂质代谢异常,并导致关键脂质转运蛋白的表达减少(例如, CD36、Fabp1/2),以及血清中甘油三酯和游离脂肪酸的减少。
自噬是由能量应激诱导的,以维持结肠细胞的稳态。结肠肠道微生物组产生的丁酸盐确实是结肠细胞的唯一碳源。转化为丁酰辅酶A后,它被动扩散到线粒体中,进行β-氧化,并供给三羧酸循环和氧化磷酸化以产生能量并抑制自噬激活。
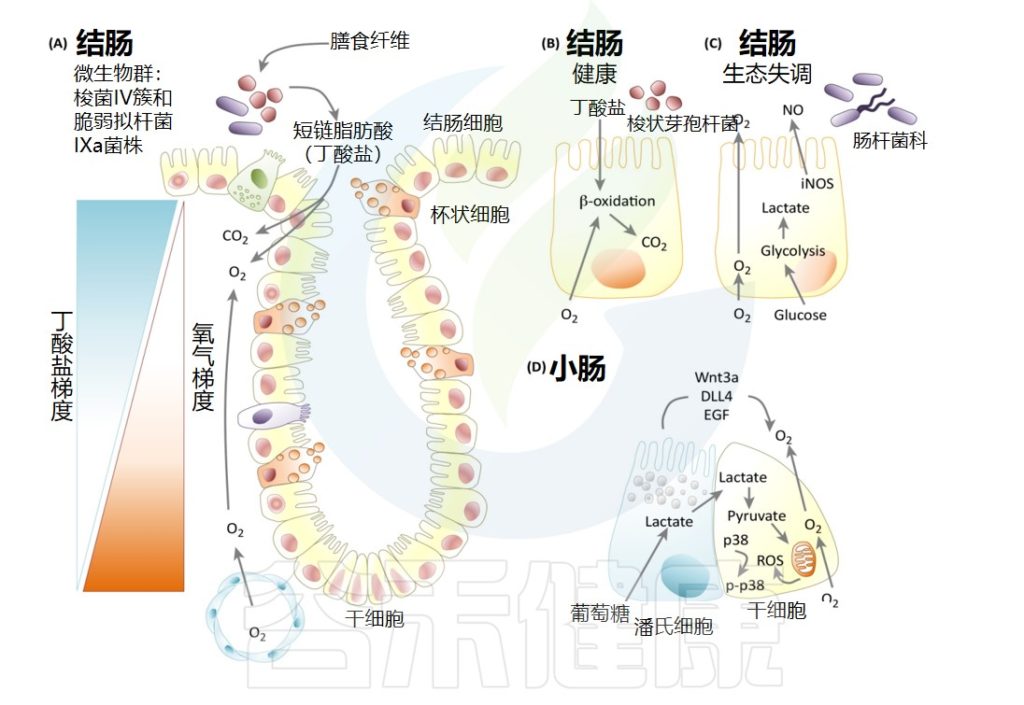
Allaire JM,et al.Trends Immunol.2018
肠上皮细胞大量暴露于肠道微生物,并产生粘液和抗菌肽以保持安全距离。丁酸盐还通过下调人类肠三批细胞中的IDO1表达和犬尿氨酸途径来促进肠道稳态。
IDO1在我们前面色氨酸代谢那篇文章中讲到过:
色氨酸通过犬尿氨酸通路的分解代谢,该通路占整体色氨酸降解的约95%,形成主要最终产物NAD+。
注:犬尿氨酸通路是炎症和免疫反应的重要参与者。
首先,色氨酸被转化为N-甲酰基-L-犬尿氨酸
该反应由三种限速酶之一催化:
TDO(色氨酸 2,3-双加氧酶) -肝内
IDO1(吲哚胺 2,3-双加氧酶 1) -肝外
IDO2(吲哚胺 2,3-双加氧酶 2) -肝外
注:这三种酶都是血红蛋白,并使用分子O2作为共底物,这也使它们能够利用活性氧 (ROS) 并调节细胞内的氧化还原平衡。
IDO和TDO酶在不同的组织中表达,暴露于不同的刺激物时被诱导,表明它们在健康和疾病中具有不同的功能。
TDO在基础条件下催化色氨酸分解,而在免疫调节中具有关键作用的IDO-1受到多种刺激物的诱导和调节,例如炎症信号 。
√肠道微生物群影响肠嗜铬细胞产生血清素
在不同的肠上皮细胞类型中,肠嗜铬细胞负责产生血清素,这对免疫细胞有重大影响。结肠中血清素的产生很大程度上受到肠道微生物群,特别是孢子形成细菌代谢物的调节。
其机制尚未完全阐明,但已表明,短链脂肪酸(丁酸和丙酸)和一些次级胆汁酸(例如通过微生物生物转化产生的脱氧胆酸)可以上调TpH1表达(血清素生产中的限速酶)。
即使需要进一步研究,这些数据表明,调节肠道微生物群组成或直接施用微生物代谢物可以从治疗角度控制血清素的产生。
▼
巨噬细胞是一种具有细胞吞噬能力的细胞,源自单核细胞,而单核细胞又来源于骨髓中的前体细胞。
√巨噬细胞在肠道稳态中发挥多种作用
巨噬细胞在维持肠道稳态方面发挥多种作用。与其他组织中的巨噬细胞一样,肠壁中的巨噬细胞也具有强烈的吞噬能力。然而,虽然具有高度杀菌作用,同时还不会导致明显的炎症反应 。
吞噬
肠固有层巨噬细胞具有高度吞噬能力,负责清除凋亡和衰老的上皮细胞。通过表达组织重塑金属蛋白酶和分泌刺激上皮干细胞更新的因子,例如前列腺素E2、肝细胞生长因子和Wnt配体,它们可促进上皮完整性。
转移
巨噬细胞可以将获得的抗原转移至迁移性树突状细胞,以呈递给引流肠系膜淋巴结中的T细胞。通过产生免疫调节细胞因子,例如IL10和TGFβ,它们维持并促进局部调节性T细胞的二次扩增。
以类似的方式,巨噬细胞通过产生IL1β来支持Th17细胞和ILC3,IL1β是通过暴露于微生物群或其衍生物而诱导的。巨噬细胞也存在于肠壁的更深层,包括粘膜下层和外肌层。
促炎或抗炎
巨噬细胞是免疫系统的关键介质,几乎遍布整个胃肠道。巨噬细胞对炎症状态具有重要影响,具有促炎和抗炎功能。它们的极化状态与许多代谢紊乱有关,例如炎症性肠病。
注:炎症M1细胞和抗炎M2细胞的表型和功能之间的平衡受细胞外和细胞内刺激的调节,从而决定疾病的进展情况。
√肠道微生物诱导巨噬细胞
现在有多种证据表明微生物群在影响粘膜巨噬细胞群动态方面发挥着关键作用。
首先,微生物定植后,结肠巨噬细胞区室发生了重大变化,特别是在断奶时,单核细胞通过单核细胞“瀑布”的分化已经建立。
其次,使用广谱抗生素可以减少巨噬细胞的周转,这进一步表明共生微生物群在控制巨噬细胞周转中的作用。
此外,与正常小鼠相比,无菌小鼠肠壁中发现的巨噬细胞较少。粘膜微环境实际上可能会对巨噬细胞进行招募。
如上所述,微生物群可能构成这种分化的一种刺激因素,其他可能性可能包括饮食代谢物或蠕动产生的持续机械应力。
注:有人建议机械应力可以解释心脏中胚胎来源的巨噬细胞的替换并且除了产生低度“炎症”外,它还可以简单地阻止巨噬细胞的长期停留。
巨噬细胞的能量代谢
巨噬细胞在免疫反应过程中处于第一线,但也感知微生物群并对其做出反应以控制微生物群,而不引发有害的炎症反应。
在致病反应期间,活化巨噬细胞的代谢特征根据情况而变化。在促炎性M1巨噬细胞中,三羧酸循环被破坏,导致衣康酸和琥珀酸的积累以及糖酵解的转变。
衣康酸是免疫代谢的主要参与者,具有免疫调节和抗菌作用。它还参与琥珀酸的积累,因为它通过阻断琥珀酸脱氢酶的活性来直接抑制其氧化。
琥珀酸通过氧化产生线粒体活性氧并导致IL-1β产生,从而表现出促炎作用。
Allaire JM,et al.Trends Immunol.2018
相反,M2巨噬细胞具有完整的三羧酸循环,并且主要依赖氧化磷酸化。肠道微生物群是通过短链脂肪酸来调节这些过程。丁酸盐(但不是乙酸盐或丙酸盐)将巨噬细胞代谢重新编程为氧化磷酸化和脂质代谢,从而导致抗炎M2表型。
▼
先天性淋巴细胞家族由自然杀伤细胞、ILC1、ILC2和ILC3组成,它们参与对病毒、细菌、寄生生物和转化细胞的免疫反应。
√先天淋巴细胞对感染和损伤做出免疫反应
有不同类型的先天淋巴细胞,其特征是表达特定的膜标记、转录因子和细胞因子特征。在激活过程中,先天淋巴细胞会深刻改变其能量代谢以适应其新功能。
先天淋巴细胞,通过产生细胞因子和分泌蛋白对感染或损伤作出反应,这些细胞因子和分泌蛋白指导和增强攻击前线的免疫反应。
√肠道微生物群影响先天淋巴细胞功能
肠道微生物群影响先天淋巴细胞功能,抗生素对ILC1、ILC2和ILC3转录组程序的巨大影响证明了这一点。
ILC3是胃肠道中存在的先天淋巴细胞的主要类型。这些细胞表达RORγt,可以产生IL-17和IL-22,是炎症、感染、微生物群组成和代谢的重要调节因子。
ILC3功能例如肠上皮防御的维持,依赖于昼夜节律调节器ARNTL介导的昼夜节律信号。但众所周知,肠道微生物群是昼夜节律的影响因素。
肠道微生物群衍生的丁酸盐也可调节ILC2功能,抑制其不受控制的激活,从而抑制其在肺部炎症和哮喘中的负面作用。
▼
哺乳动物的神经系统分为中枢神经系统和周围神经系统。中枢神经系统包括大脑和脊髓,周围神经系统包括神经节,神经节是神经细胞体的聚集体,头部、颈部和内脏中的神经束从神经细胞体中产生。
注:消化道同时受自主神经和管道壁上的肠神经丛的神经支配。
√交感神经直接从大脑连接到胃肠道
从中枢神经系统到肠神经系统的外部连接由交感神经纤维和副交感神经纤维组成。离开脑后,副交感神经和交感神经可以直接突触到胃肠道。
例如,副交感迷走神经,离开后脑后,沿着食道穿过隔膜,最终突触到胃肠道。交感神经起源于脊柱,并突触到交感内脏神经节,例如腹腔神经节、肠系膜上神经节和肠系膜下神经节。副交感神经和交感神经均可直接突触至肌间神经节、平滑肌和粘膜。
√盆腔神经支配远端结肠和直肠
此外,起源于脊髓并通过骶脊神经离开的盆腔神经支配远端结肠和直肠。盆腔神经传统上被认为是副交感神经。副交感神经对交感神经节的神经支配使胃肠道的神经连接变得更加复杂。
最后,内在的肠神经系统是沿着胃肠道的神经元和神经胶质细胞的扩展网络;这些可以自主发挥作用,但也可以通过肠神经系统与外在交感神经和副交感神经的连接进行调节。因此,中枢神经系统和肠神经系统之间的通信是双向的。
√肠神经元上的受体介导重要的胃肠道功能
肠神经元上的受体介导重要的胃肠道功能。机械感受器对粘膜磨损敏感;张力感受器对拉伸敏感;化学感受器对管腔内的各种化学刺激做出反应,例如pH值、渗透压和营养物质。此外,各种受体负责调节肠道内的液体交换。
2021年美国一项研究发现肠道神经细胞可以通过分泌细胞因子IL6来调控调节性T细胞的水平。
此外,调节性细胞诱导菌很有可能是通过降低肠道神经系统密度和神经细胞IL6水平的双重作用,来诱导调节性T细胞的。
▼
潘氏细胞于19世纪末首次描述为柱状上皮细胞,胞浆内有明显的嗜酸性颗粒。它们在维持肠道稳态和调节小肠及其相关微生物菌群的生理方面发挥着不可或缺的作用。
√潘氏细胞可调节肠道稳态
潘氏细胞产生的致密颗粒含有丰富的抗菌肽和免疫调节蛋白,它们的功能是调节肠道菌群的组成。
肠道中的潘氏细胞在小肠中比较丰富,与吸收性肠细胞不同,潘氏细胞寿命长,在从干细胞分化后向下迁移到隐窝的最底部。
√潘氏细胞具有重要的抗菌作用
它们具有重要的抗菌作用,可产生溶菌酶、防御素等抗菌肽并再生胰岛衍生蛋白IIIγ,以响应白细胞介素22或刺激Toll样受体、核苷酸结合寡聚化结构域2或胆碱能神经。
潘氏细胞是专业的分泌细胞,通常通过将抗菌因子分泌到管腔中来控制肠道细菌,从而在先天免疫系统中发挥作用。潘氏细胞能够感知来自腔内细菌的信号,并通过改变这些因子的产生来做出反应,以保护上皮屏障。潘氏细胞依靠自噬来调节其分泌能力和能力。
潘氏细胞还通过产生促表皮生长因子 配体来帮助维持正常的隐窝干细胞活性,它们对于肠道稳态至关重要。
小结
总之,肠潘氏细胞在肠道内稳态的许多方面都起着关键作用,从调节与上皮密切相关的微生物区系,到维持干细胞生态位的健康,再到帮助调节炎症水平。不仅保护自己不受外来入侵,而且促进肠道的生长和发育。
这些功能在早产儿的未成熟肠道中尤其重要,早产儿的肠道发育中伴随着一个非生物菌群。因此,潘氏细胞破坏与新生儿肠道坏死的发生有机械联系是合理的。对潘氏细胞生物学的进一步了解可能会为理解新生儿肠道坏死的发展提供一条重要的途径。
▼
我们的肠道和呼吸道中布满了杯状细胞,这些细胞因其杯状外观而得名,它们能够分泌具有保护作用的凝胶状粘液,以防止肠道内细菌直接侵入肠壁组织而引发炎症反应。
这些黏液的主要成分为肠壁上杯状细胞分泌的黏蛋白。黏蛋白则肠道黏液层的重要组成部分,凝胶形成黏蛋白和跨膜黏蛋白是目前已知的两种黏蛋白。
√杯状细胞沿着胃肠道逐渐增加
与潘氏细胞相反,产生粘液的杯状细胞的频率沿着胃肠道逐渐增加。杯状细胞在远端结肠中至少占所有上皮细胞的25%,而在小肠上段则为10%或更少。
与此同时,覆盖在粘膜上的粘液层——被称为糖萼,在结肠中最厚,它由两个不同的层组成:一个附着在上皮表面的致密内层和一个外层,类似于小肠中发现的松散层。
Kayama H,et al.Annu Rev Immunol.2020
√杯状细胞分泌的粘液具有抗菌作用
粘液具有抗菌作用,形成一种高度带电的凝胶,充当物理屏障,并且由对许多细菌有直接毒性的粘蛋白糖蛋白组成。
小肠较松散的糖萼也提供了抗体和抗菌肽可以粘附的基质。粘液合成的缺陷导致共生细菌更多地渗透到结肠的上皮表面,从而增加对结肠炎和结肠癌的易感性。
杯状细胞除了分泌肠道黏液层,形成肠道组织、免疫系统与肠腔微生物群、内容物分离的重要屏障,它其实还有其他意想不到的作用。
√避免一些蛋白引起免疫反应
杯状细胞还可以“护送”抗原到树突细胞。来自华盛顿大学圣路易斯医学院的研究人员在小鼠肠道中鉴定出一种“护送”食物中的抗原或蛋白到树突细胞的细胞,即杯状细胞,这样免疫系统就不会对它们发起免疫。这项发现给科学家提供一种潜在性靶标来治疗炎性肠道疾病、乳糜泻和食物过敏,这些疾病都是由于免疫系统过分活跃而导致的。
√杯状细胞与痛觉感受神经联系保护肠道
2022年,哈佛大学医学院的研究表明,肠道中的痛觉感受神经在正常情况下能够调节保护性粘液的存在,并在炎症状态下刺激肠道中的杯状细胞分泌更多粘液,从而直接发挥保护肠道的作用。
该研究详细描述了这种复杂信号级联的步骤,显示痛觉感受神经与肠道杯状细胞直接可以进行直接的串扰。
√杯状细胞增加粘液分泌阻止外来的细菌侵入
杯状细胞可以还“自杀”换取结肠的安全。
哥德堡大学的研究团队发现一类新的被称作杯状细胞的岗哨细胞能够冲洗走侵入到保护性的黏液屏障的细菌。一旦发现细菌存在于这种隐窝入口处的迹象,它们就启动一种级联反应,最终导致黏液增多,从而将细菌冲洗走。
这一发现可能为解释炎症性肠病(如溃疡性结肠炎)如何发生提供重要理论依据。
此前,该团队已证实黏液层作为第一道防线而存在。然后又证实在隐窝入口处存在一部分杯状细胞作为第二道防线阻止进入黏液层的细菌侵入结肠。
▼
T细胞在哺乳动物免疫反应中的核心作用是无可争议的。
根据所产生的细胞因子和效应细胞的生物功能特征,可将其分为辅助型T细胞1(Th1)、辅助型T细胞2(Th2)、调节性T细胞(Treg)和Th17等。
T细胞祖细胞源自骨髓中的造血干细胞。它们迁移到胸腺,在那里成熟为T细胞。随后,T细胞迁移通过次级淋巴器官,扫描呈递其同源抗原的抗原。
√肠道内含有大量的T细胞
肠道是体内最大的T细胞群之一的所在地。粘膜T细胞对于体内平衡和保护性免疫至关重要,但也在不受控制的(慢性)粘膜炎症中发挥作用,如乳糜泻或炎症性肠病,甚至可能在肿瘤的发展过程中。
胃肠道的上皮屏障是大量上皮内T细胞的家园。上皮内T细胞在功能上适应粘膜环境,是最早遇到微生物和饮食抗原的适应性免疫细胞之一。
√上皮内T细胞的免疫作用
上皮内T细胞在免疫监视中发挥重要作用,同时诱导对非病原性抗原的耐受性,从而保持单层上皮膜的完整性。
炎症性肠病和乳糜泻的特点是(复发性)胃肠道炎症占主导地位。上皮内T细胞在肠上皮屏障中的寿命长且大量存在,表明上皮内T细胞在这些疾病的复发和缓解过程以及持续的低度炎症中发挥作用。
作为组织特异性和潜在致病细胞,上皮内T细胞是治疗(非系统性)慢性组织特异性炎症性疾病(如 炎症性肠病)的理想靶点。
启动的T细胞会长期驻留在肠道中
在炎症或感染环境中,已启动的T细胞会暂时从循环中募集到肠道,但大量T细胞会永久驻留在肠道中,并且独立于淋巴和循环记忆群体而维持。
这些T细胞主要由T细胞受体αβ + CD8αβ +和TCRαβ + CD4组成+组织驻留记忆T细胞,但调节性T细胞 (TCRαβ + CD4 + FOXP3 + )、TCRγδ + T 细胞、自然杀伤T细胞和粘膜相关不变T细胞也建立长期驻留在肠道。
T细胞的能量代谢
T细胞代谢可塑性对于适应永久动态的免疫环境是必要的。肠道微生物群通过活性氧、短链脂肪酸和胆汁酸的产生以及氧化还原信号传导的修饰积极参与这一复杂过程。
效应T细胞和记忆T细胞具有不同的功能和需求,因此表现出不同的代谢。它在效应T细胞中由有氧糖酵解主导,在记忆T细胞中由脂肪酸氧化和氧化磷酸化 主导。记忆T细胞中存在融合的线粒体网络,效应T细胞中存在点状线粒体。
此外,线粒体是T细胞激活的关键组成部分,主要通过活性氧产生。通过CD3刺激T细胞会诱导钙内流,从而刺激丙酮酸脱氢酶和三羧酸循环酶的功能。
√短链脂肪酸可以增强T细胞功能
微生物群衍生的短链脂肪酸通过改变细胞代谢来增强CD8+ T细胞效应功能。肠道微生物群代谢膳食纤维产生的短链脂肪酸会刺激CD8+ T细胞中的氧化磷酸化和线粒体质量及其糖酵解能力。
此外,短链脂肪酸可以扩散到细胞质中并作为脂肪酸氧化的底物,导致乙酰辅酶A的产生,为三羧酸循环和氧化磷酸化提供燃料。
在活化的CD8+ T细胞中,短链脂肪酸,特别是丁酸,促进胆汁酸的摄取和氧化,导致三羧酸循环与糖酵解输入断开,并通过胆汁酸分解代谢和谷氨酰胺利用有利于氧化磷酸化。这种丁酸盐诱导的细胞代谢适应是分化为记忆T细胞所必需的。
短链脂肪酸对CD4+ T细胞也表现出显著影响,特别是在辅助性T(Th)17、Th1的生成方面和调节性T细胞。其机制涉及组蛋白去乙酰化酶的抑制和mTOR通路(细胞生长和代谢的主要调节因子)的调节。
最近发现了戊酸的这种联系,它可以通过为组蛋白乙酰转移酶提供额外的乙酰辅酶A并增强糖酵解和 mTOR 活性来刺激抗炎细胞因子IL-10的产生。
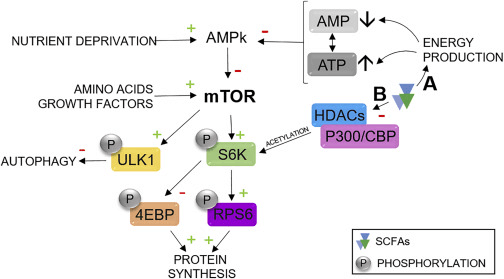
短链脂肪酸激活mTOR的机制
短链脂肪酸激活mTOR的机制有两种。短链脂肪酸通过对能量产生途径的作用,诱导ATP的产生和AMP的消耗,它们分别是AMP激活蛋白激酶 的抑制剂和激活剂。因此,AMPK对 mTOR的抑制剂活性被抑制,从而导致mTOR激活。
第二个潜在机制涉及短链脂肪酸的组蛋白去乙酰化酶抑制活性。短链脂肪酸与 P300/CBP(E1A 结合蛋白 p300/CREB 结合蛋白)结合,促进核糖体蛋白 S6 激酶 beta-1 (S6K1) 的乙酰化,这是mTOR 的下游靶标,从而更强有力地激活途径。
√胆汁酸对T细胞的影响
胆汁酸对T细胞也有重要影响。石胆酸的衍生物3-oxoLCA通过直接与转录因子RORγt相互作用来抑制Th17细胞的分化。
相反,石胆酸的另一种衍生物isoalloLCA促进调节性T细胞的分化。该机制涉及刺激OXPHOS和mROS的产生,从而通过增加Foxp3启动子中组蛋白 (H3K27) 乙酰化的水平来增加FOXP3的表达。
具体来说,在结肠中,胆汁酸通过胆汁酸受体 Breg 调节 RORγ+ Treg 细胞的功能,这些细胞在维持结肠稳态中发挥着重要作用。
▼
B细胞是一种在适应性免疫过程中发挥重要功能的淋巴细胞,B细胞及其所生产的抗体,对于免疫系统的正常运行是十分重要的。
√B细胞通过产生抗体阻止外来病原体
B细胞是可以产生抗体的白细胞。这些抗体或免疫球蛋白可以与有害的外来颗粒(如病毒或致病细菌)结合,阻止它们入侵和感染人体细胞。
每个B细胞携带一个单独的B细胞受体,该受体决定它可以结合哪些病原体,就像每个锁接受不同的钥匙一样。
人类肠道中B细胞谱系的两个主要区室是肠道相关淋巴组织和弥漫性淋巴组织中高度组织化的B细胞的聚集地,它们占据了大肠和小肠的广泛肠固有层 。
肠道相关淋巴组织可以以滤泡簇的形式出现,通常被称为派尔氏淋巴结或孤立的淋巴滤泡,分布在整个胃肠道中,是对从肠腔采样的抗原做出长期反应的免疫诱导位点。
研究人员使用专门设计的计算机程序来处理数百万个基因序列,这些序列会比较来自B细胞的抗体库,这取决于这些微生物是留在肠道里,还是进入血液。在这两种情况下,抗体库都改变了,但改变的方式不同,取决于暴露发生的方式。
√肠壁与血液中的抗体不同
与血液中的抗体(IgM和IgG)相比,肠壁(IgA)中存在不同种类的抗体。通过强大的基因分析,研究人员发现,肠道产生的不同抗体的范围远小于身体中央组织产生的抗体。
这意味着一旦微生物进入人体,免疫系统就有更多的可能去中和和消灭它们,而肠道中的抗体主要结合在任何时候都能看到的细菌分子。
B细胞有时会影响受损肠道的愈合
B细胞有时也会好心办坏事。来自瑞典研究组发表研究结果发现,B细胞的数量在肠道损伤后大幅增加,并且阻碍了损伤组织的愈合。这一研究结果对炎症性肠病的治疗具有重要意义。
√短链脂肪酸有助于B细胞的能量代谢
B细胞分化为浆细胞和产生抗体需要大量能量和涉及细胞代谢的整体变化。肠道微生物群衍生的短链脂肪酸有助于在不同水平上为这些过程的细胞能量引擎提供燃料,并促进抗体的产生。
短链脂肪酸转化为乙酰辅酶A,并整合到线粒体三羧酸循环循环中,从而产生ATP。短链脂肪酸还通过mTOR激活刺激B细胞中的糖酵解。短链脂肪酸衍生的乙酰辅酶A也是胆汁酸(特别是棕榈酸)合成的底物,这对于浆细胞分化并刺激抗体产生至关重要。
注:最近研究表明,异丁酸或异戊酸等分支短链脂肪酸也可以调节B细胞功能。受操纵的小鼠中缺乏分支短链脂肪酸产生导致小肠中 IgA+ 浆细胞的频率增加,以及与先天免疫细胞(如中性粒细胞、巨噬细胞和树突状细胞)表面结合的IgA水平增加。
此外,B细胞通过产生免疫球蛋白和产生IL-10的调节性B细胞的作用,在对肠道微生物群的耐受性中发挥关键作用。
肠道微生物群与免疫系统之间存在着紧密的相互作用,这种作用对于维持免疫系统的正常功能和免疫平衡至关重要。
肠道微生物群维持上皮屏障的完整性并塑造粘膜免疫系统,通过微生物代谢物、成分和对宿主细胞的附着来平衡宿主免疫防御和口服耐受性。
为了避免异常的免疫反应,上皮细胞通过构建化学和物理屏障将肠道微生物群与免疫细胞隔离开来,从而建立宿主共生互惠关系。
这是为什么人们努力表征肠道微生物群在健康和疾病中的功能。许多研究报告和我们检测实践报告了众多慢病的临床前和临床模型中微生物群的改变。尽管这些改变可能部分是由疾病状态驱动的,但最近的研究强调,改变的微生物群也可以在驱动疾病本身方面发挥核心作用。
▼
我们肠道内的良性微生物数量与我们体内的细胞数量大致相同。这些细菌大多停留在肠道内,而不是渗透到人体组织中。
不幸的是,有些渗透是不可避免的,因为肠道只有一层细胞,把我们吸收食物所需的血管和管子内部分开。
✦细菌的数量沿着胃肠道逐渐增加
细菌的数量通常会沿着胃肠道增加,从胃的高酸性环境中的每毫升100–1,000个到小肠上段的每毫升约105个,在结肠中高达每毫升1010-1011个。
然而,回肠末端可能含有比结肠中更多的细菌。这些细菌使用复杂的多糖和粘液的其他成分以及未消化的纤维作为能量来源,产生生物素、短链脂肪酸 和维生素等必需的代谢物。
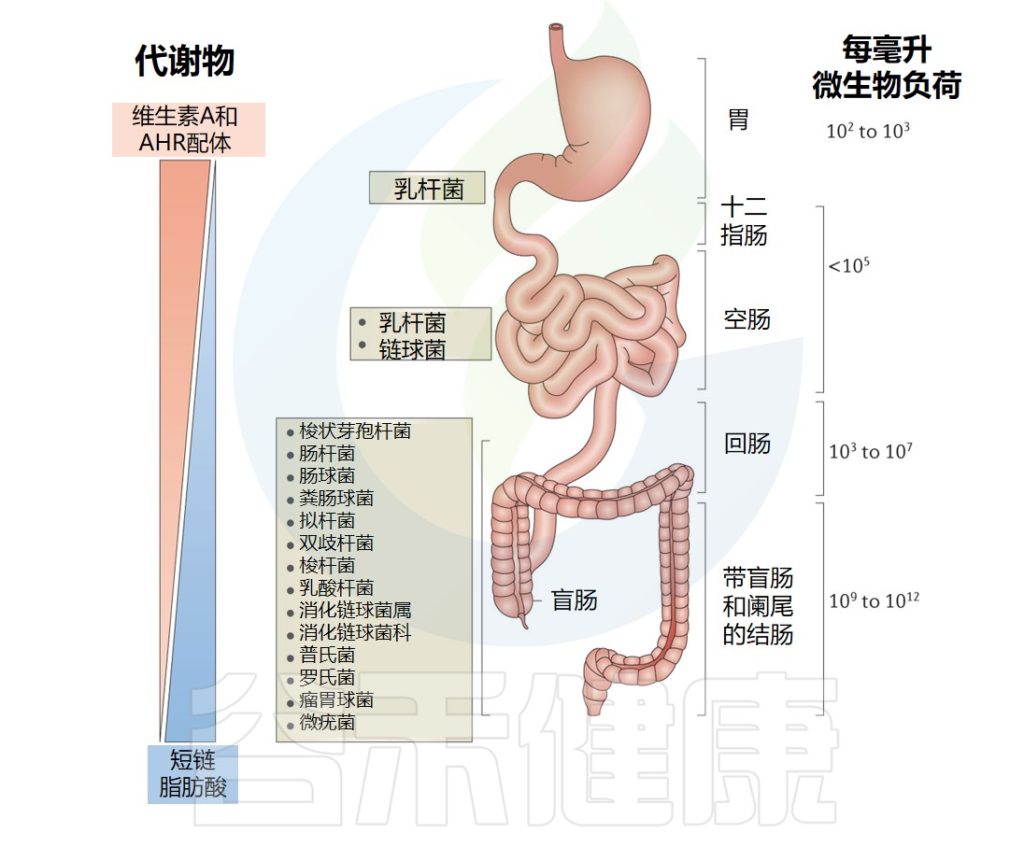
Mowat AM,et al.Nat Rev Immunol.2014
由于结肠大多数物种是专性厌氧菌,难以在体外培养,还有部分种群的组成和分布尚未最终确定。然而,分子技术揭示了几个主要门类中大约200-2000 个物种;它们的相对数量因人而异,但厚壁菌门、拟杆菌门、变形菌门和放线菌门最为普遍。
也包括少部分古细菌,真菌和病毒。好氧菌在小肠上段普遍存在,而厌氧菌在结肠中占主导地位。
✦不同的疾病会导致肠道菌群分布改变
构成肠道微生物群的物种分布的改变(生态失调)与许多不同的疾病有关,从精神疾病到代谢性疾病、过敏和自身免疫病。其中盲肠中细菌密度相较于小肠较高,该部位肿瘤发生率较高的原因。
肠道不同区域接触各种饮食成分可影响免疫功能的程度也存在显著差异。例如,维生素A只存在于饮食中,也可以通过胆汁输送到小肠,类黄酮和芳基烃受体的其他配体在小肠中的含量也较高。
▼
肠道微生物通过多种机制与肠外器官中的免疫细胞进行交流。
微生物能够通过直接与免疫细胞作用,如粘附到细胞表面或被吞噬细胞摄取,来调节免疫细胞的功能和活性。
其次,微生物通过与肠道黏膜上皮细胞和巨噬细胞表面的受体结合,激活免疫细胞,使它们产生免疫应答和促炎信号,并进一步调节免疫细胞的活性。
微生物还通过产生代谢产物,如短链脂肪酸和其他微生物分子,与免疫细胞进行信号传导。这些代谢产物可以直接调节免疫细胞的活性和功能,影响它们的增殖、分化和效应物质的产生。
✦肠道微生物群影响免疫系统的发育
首先,肠道微生物群对于免疫系统的发育起着重要的影响。在婴幼儿期,肠道微生物群通过与免疫系统的交互作用,促进免疫系统的成熟和功能发展。特别是在胎儿期和早期生活中,良好的肠道微生物组成对于建立免疫耐受、预防过敏反应和自身免疫性疾病的发生非常重要。
微生物群刺激对于产生免疫球蛋白 (IgA) 的B细胞的发育和成熟、通过促进IgA类别转换的记忆浆细胞以及生发中心的发育至关重要。
此外,通过介导浆细胞样树突状细胞产生I型干扰素,微生物群增强了抗原特异性T细胞反应。
✦肠道微生物可以直接抵抗病原体
肠道微生物群赋予宿主免疫保护的最直接例子是通过对粘膜病原体(如艰难梭菌)的定植抵抗。
多样化的肠道微生物群通过各种机制帮助宿主抵抗病原体定植,包括微生物间对营养的竞争、代谢竞争和直接拮抗以及粘膜屏障完整性的强化和局部粘膜免疫防御。
通过这种方式,肠道共生微生物保护宿主免受粘膜感染,以及由于肠道病原体过度生长和转移到循环系统和远端器官中而可能发生的继发性全身感染。
此外,之前也有报道称,病原体入侵是人类代谢失调的一个特征。这些研究进一步说明了在粘膜表面需要良好的宿主/微生物群相互作用。
// 双歧杆菌对免疫细胞的影响
本文以双歧杆菌(Bifidobacterium)为例:在小鼠肥胖模型中,给予假链状双歧杆菌通过恢复调节性T细胞和B淋巴细胞的平衡状态来减少全身炎症。
类似的免疫稳态特性已归因于双歧杆菌菌株,其通过体外刺激树突状细胞,诱导Th17谱并增强调节性T细胞从幼稚淋巴细胞的分化。
双歧双歧杆菌DSM 20082裂解物刺激外周T细胞可增加CD8+ T细胞的细胞毒活性,而对CD4+ T细胞活性没有任何影响。
✦肠道微生物通过其代谢物影响免疫细胞
肠道微生物产生的代谢产物能够刺激肠道上皮细胞和免疫细胞,如树突状细胞和巨噬细胞,促进免疫细胞的活化和增殖。
肠道微生物通过膳食分子的修饰(脂肪酸、色氨酸代谢物,氧化三甲胺)、宿主胆汁酸(次级胆汁酸)以及从头细菌代谢物(多胺、维生素、支链氨基酸),可以促进全身免疫反应的调节。
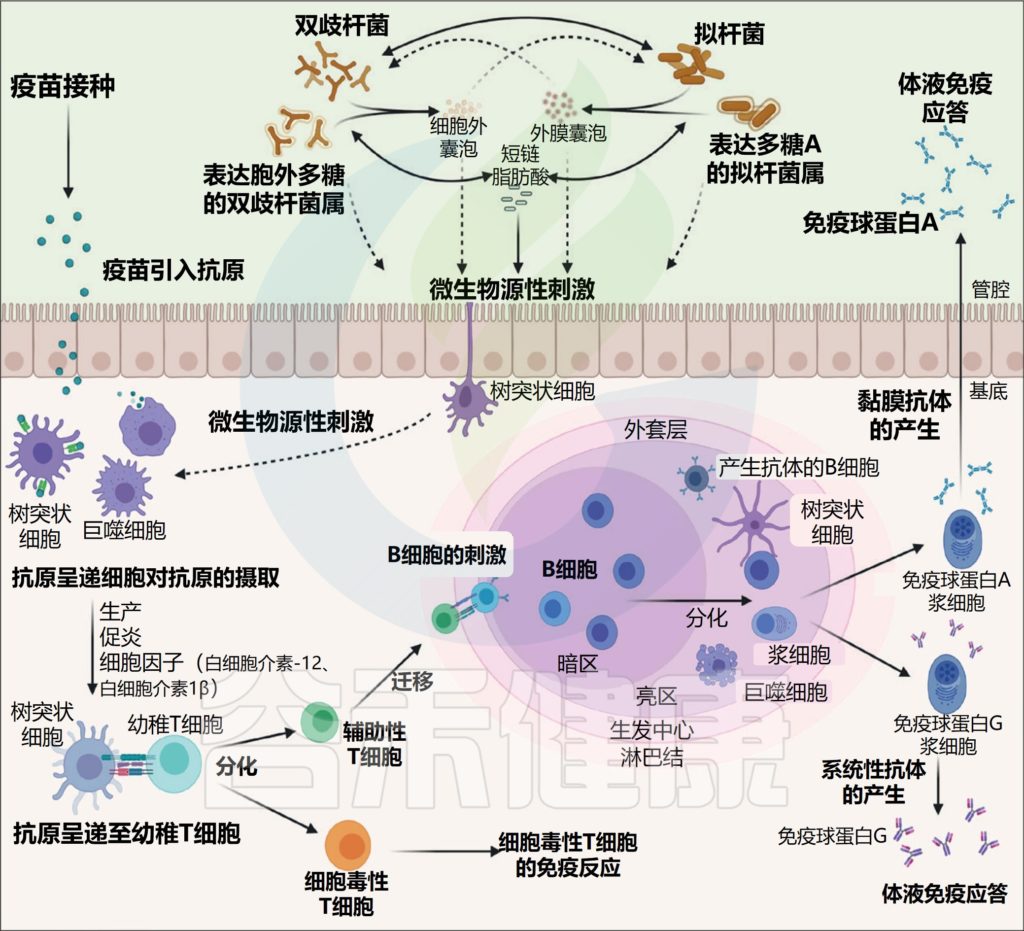
肠道微生物群的免疫调节
Jordan A,et al.Lancet Microbe.2022
// 乙酸盐可以激活免疫细胞
乙酸盐可以激活B细胞、T细胞亚群、中性粒细胞、巨噬细胞、树突细胞和肠上皮细胞上表达的G蛋白偶联受体43(GPR43),导致固有层调节性T细胞增殖,和调节自身抗体的产生和边缘区B细胞。
// 丙酸盐影响巨噬细胞浓度
丙酸盐诱导小鼠调节性T细胞的分化和增殖以及白细胞介素10的表达。丙酸盐激活GPR15和GPR43还通过降低组蛋白去乙酰化酶6和9的表达以及抑制 NF-κB信号传导来增加结肠调节性T细胞的浓度。
丙酸盐激活GPR41会改变骨髓造血功能,导致巨噬细胞和树突状细胞前体浓度升高,并扭曲Th2分化。
// 细菌胞外多糖影响免疫反应
脆弱拟杆菌的表面多糖A可激活巨噬细胞上的Toll样受体2,并诱导调节性T细胞的扩增和抗炎白细胞介素10的产生,从而在病毒感染期间促进强烈的抗炎反应。
来自长双歧杆菌BCRC14634的胞外多糖被证明可增强巨噬细胞产生白细胞介素,与同基因的胞外多糖阴性突变体相比,来自B.longum亚种longum 35624的胞外多糖显示通过抑制促炎细胞因子产生来抑制促炎性Th17细胞的扩增。
✦肠道微生物调节神经免疫信号
肠道和肠外器官之间的信号传导也可以通过肠道微生物调节的神经免疫机制来完成。肠道细菌可以合成γ-氨基丁酸等神经递质,并调节各种影响免疫细胞功能的神经递质。
此外,肠道细菌刺激肠内分泌细胞释放神经内分泌肽激素,这些细胞可以进入循环系统并作用于全身。
▼
肠道微生物群通过多种途径调节免疫系统的功能,包括维持免疫平衡、促进免疫耐受、保护黏膜屏障等。
➤ 自身免疫性疾病
肠道微生物群对免疫耐受的维持至关重要
自身免疫性疾病分为肠道相关疾病、非肠道相关疾病和免疫特异性疾病。
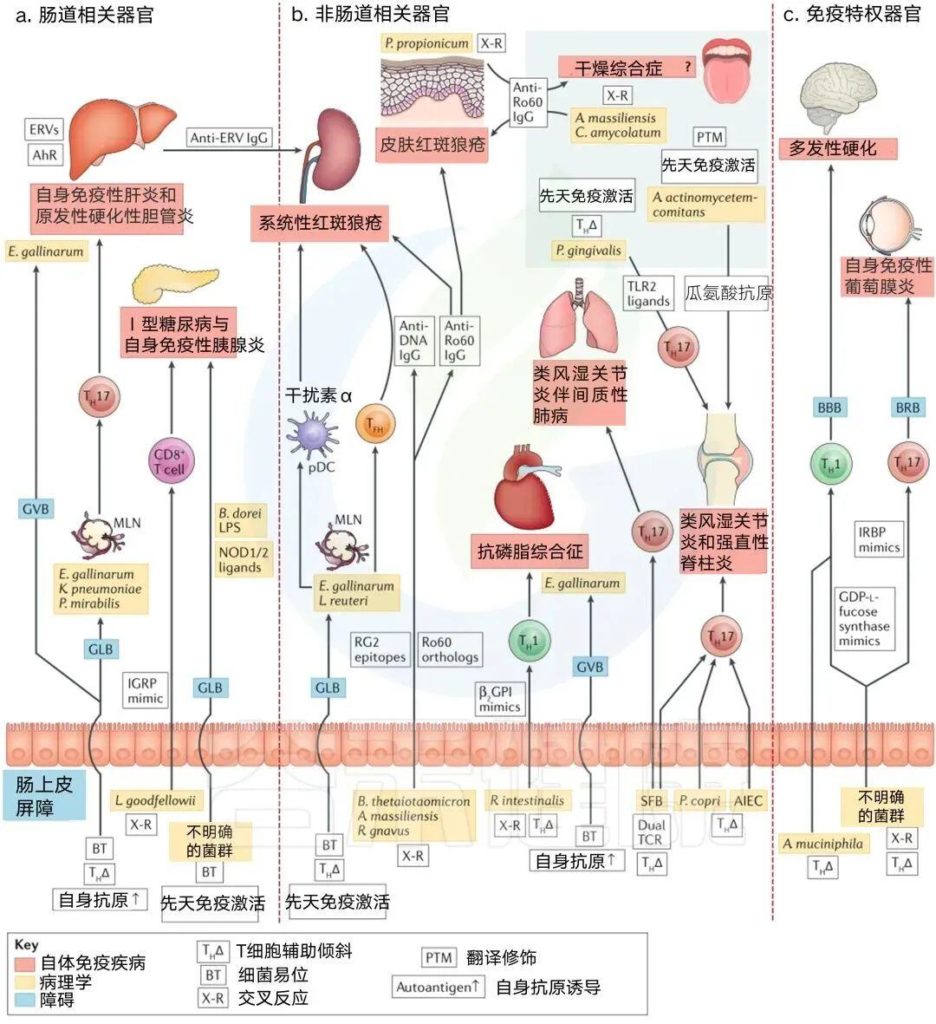
a | 肠相关器官。鸡肠球菌(Enterococcus gallinarum)向肝脏的转移促进了自身免疫性肝炎,并与肺炎克雷伯菌和奇异变形杆菌一起,通过包括TH17分化在内的多种机制导致原发性硬化性胆管炎。鸡肠球菌诱导的肝芳香烃受体(AhR)和内源性逆转录病毒(ERV)也参与了小鼠狼疮性肾炎(一种非肠道相关器官)的系统性抗ERV反应。
Leptotrichia goodfellowii 能与小鼠Ⅰ型糖尿病自身抗原胰岛特异性葡萄糖-6-磷酸酶催化亚单位相关蛋白(IGRP)发生交叉反应。在小鼠Ⅰ型糖尿病中,未定义的微生物群易位到胰腺淋巴结并激活NOD2,而来自微生物群的NOD1配体的易位驱动自身免疫性胰腺炎。
b | 非肠道相关器官。鸡肠球菌和乳杆菌在系统性红斑狼疮模型中向肠系膜淋巴结(MLNs)、肝脏和脾脏转移,从而激活先天性(浆细胞样树突状细胞(pDC))途径。
鸡肠球菌还刺激狼疮患者滤泡辅助因子(TFH)和自身抗体的产生。Ro60直系表达细菌在狼疮患者中引起Ro60自身免疫反应;肠道中的类泰奥托米克龙杆菌、口腔或肠道中的大量放线菌和阿米巴棒状杆菌,除了狼疮外,还可能导致Sjógren综合征,丙酸假丙酸杆菌与亚急性皮肤狼疮皮损通过交叉反应。
Ruminococcus gnavus在狼疮性肾炎时扩张,可与狼疮双链DNA发生交叉反应。抗磷脂综合征是一种凝血性自身免疫性疾病,鸡肠球菌传染性支气管炎也加重了这种疾病,它诱导肝脏中的自身抗原β2GPI。
口腔牙龈卟啉单胞菌、普雷沃氏菌和小肠节段丝状菌(SFB)的TH17偏斜均与类风湿关节炎的发病有关。同样,粘附性侵袭性大肠杆菌(AIEC)可导致克罗恩病相关性脊柱炎。在关节炎模型(未显示)中,SFB还使CD4+T细胞向TFH细胞倾斜,并通过携带双TCRs的TH17细胞在同一模型中引发间质性肺病。口腔中的放线菌群通过一种也能激活中性粒细胞的毒素,瓜氨酸化类风湿关节炎中的系统自身抗原。
c | 免疫特权器官。Akkermansia muciniphila在多发性硬化症中增强TH1反应,并且未定义的微生物群与GDP-l-岩藻糖合酶(一种神经炎症疾病的自身抗原)交叉反应。未定义的小鼠肠道微生物群与TH17细胞交叉反应,识别自身免疫性葡萄膜炎的自身抗原——视黄酸结合蛋白(IRBP)。
扩展阅读:
➤ 过敏性疾病
肠道微生物群可以影响免疫系统的平衡
肠道微生物群的失调可能导致免疫系统的异常激活,从而增加过敏性疾病的风险。肠道微生物群可以通过多种途径影响免疫系统,包括调节免疫应答的平衡、维持免疫耐受等。
以过敏性鼻炎为例,典型的过敏性鼻炎肠道菌群促进细胞因子的不利变化,从而促进过敏性鼻炎发展中涉及的 Th1/Th2 失衡。
过敏性鼻炎患者的肠道菌群失调及其对 Th1/Th2 平衡的影响
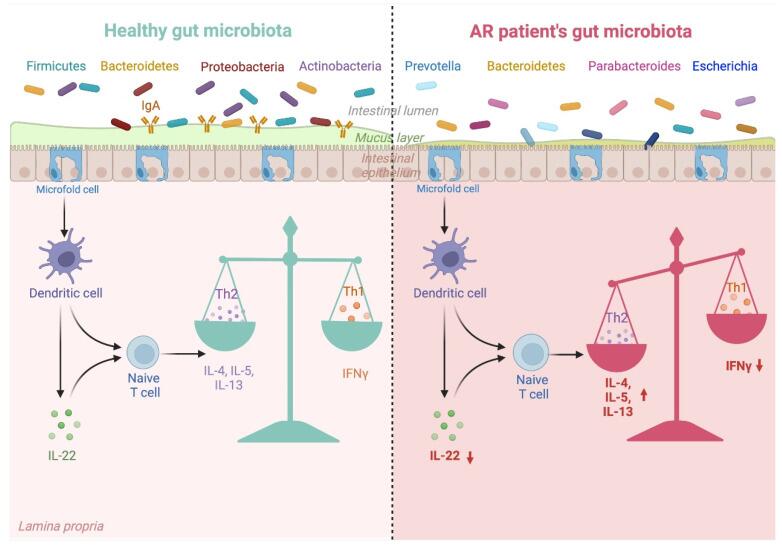
Kaczynska A, et al., Nutrients. 2022
肠道菌群的每一次紊乱都会对免疫系统产生负面影响,并破坏先天性和适应性反应。上图解释了改变的肠道微生物群组成与其对过敏性鼻炎患者免疫系统的影响之间的联系。
肠道微生物群失调影响过敏性鼻炎
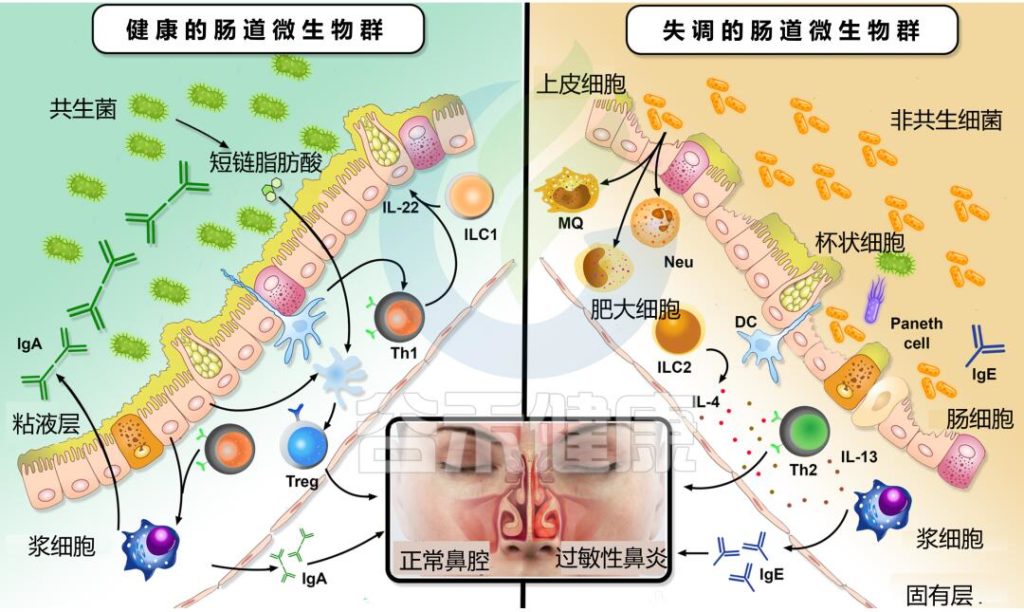
Li J, et al., Eur Arch Otorhinolaryngol. 2022
肠道微生物群影响ILC2通过肠-肺轴从肠道向肺的迁移。例如,变形菌门显著促进了上述迁移,并促进了IL-33的产生。此外,呼吸道过敏的发展与Ruminococcus gnavus的丰度增加有关联。
扩展阅读:
➤ 肠道炎症性疾病
肠道微生物群对保护黏膜屏障功能至关重要
与溃疡性结肠炎患者或健康受试者相比,克罗恩病患者的变形菌门显著增加。在炎症克罗恩病组织中,埃希氏菌(Escherichia)和志贺氏菌(Shigella)更为丰富,而在非炎症克罗恩病组织里,Pseudomonas更为广泛。
大肠杆菌和梭杆菌能够粘附粘膜,侵入肠上皮细胞,并可能加剧炎症。具核梭杆菌通过破坏上皮屏障和诱发异常炎症而加重结肠炎。
相对于炎症性溃疡性结肠炎,拟杆菌门在克罗恩病炎症粘膜中更常见。克罗恩病粘膜活检中最常见的是普通拟杆菌(Bacteroides vulgatus),克罗恩病患者中脆弱拟杆菌(Bacteroides fragilis)的数量明显高于溃疡性结肠炎患者或健康受试者。脆弱拟杆菌与产生IL-10的调节性T细胞的增加有关,这限制了促炎机制,从而有助于减少炎症。
肠道微生物群在调节宿主免疫以建立和维持肠道稳态方面起着至关重要的作用。
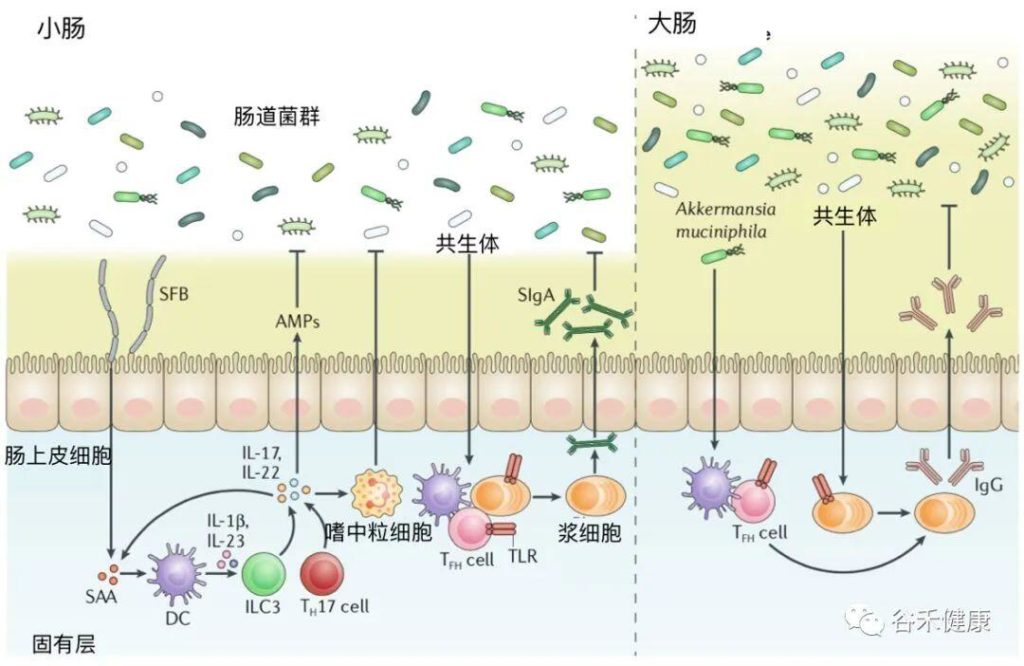
doi: 10.1038/s41577-019-0268-7
失调可能分为早期和晚期两个阶段进行。
在早期失调期间,IBD相关的遗传和环境因素可能导致致病菌的积累,这可能先于临床疾病的发展。
在晚期失调期间,肠道炎症推动了菌群的进一步变化,包括变形菌的增殖。鉴于肠道微生物群的不同菌群对宿主免疫系统和肠道屏障有有益的影响,某些菌群的缺失可能导致肠道炎症的加剧或消退。特定细菌的大量繁殖,如粘附性和侵袭性大肠杆菌,积聚在IBD患者的炎症粘膜中,可进一步促进炎症反应。
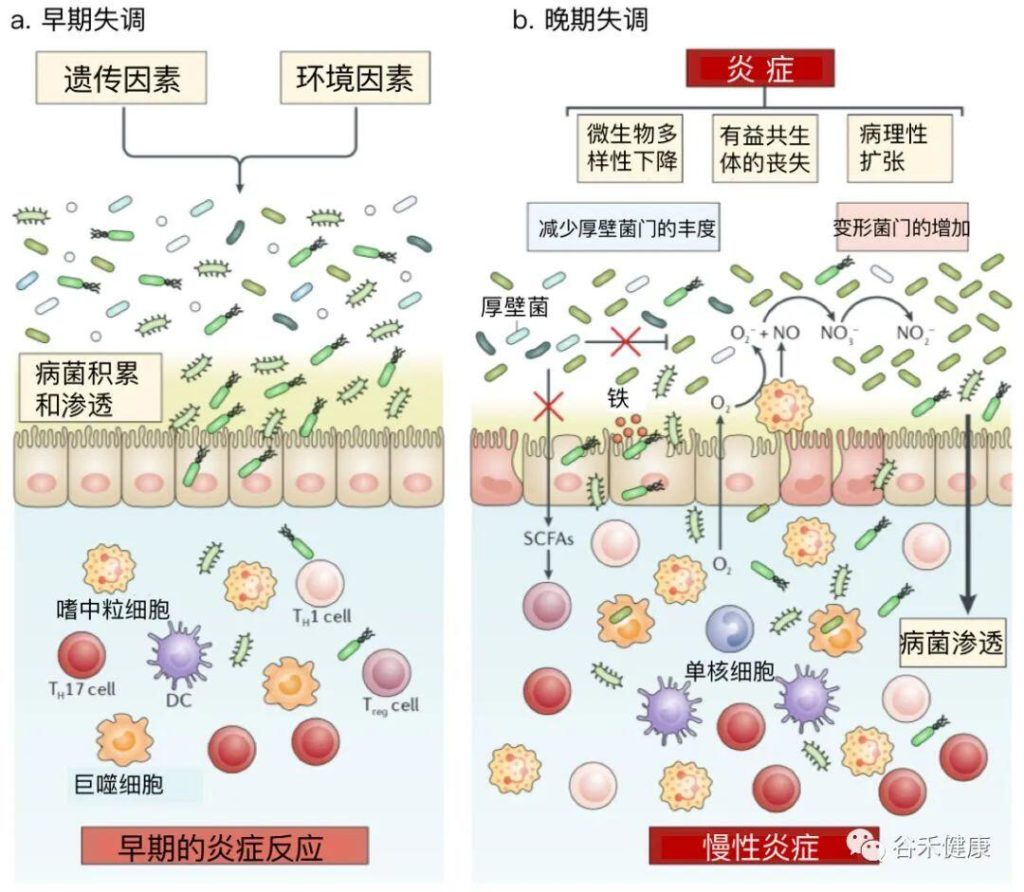
doi: 10.1038/s41577-019-0268-7
扩展阅读:
小结
综上所述,肠道微生物群与免疫系统之间的相互作用对于维持免疫系统的正常功能和免疫平衡至关重要。
良好的肠道微生物组成能够促进免疫系统的发育和成熟,调节免疫应答的程度和方向,预防免疫相关疾病的发生。
肠道内的免疫细胞在整个机体中扮演着至关重要的角色。它们作为守卫者,保护着我们免受外界病原体的入侵,并维持着肠道的免疫平衡。
肠道免疫细胞与肠道中的微生物密切合作,形成了一种复杂的共生关系。这些微生物通过调节免疫细胞的活性和功能,帮助我们抵御病原体,促进免疫平衡和整体健康。
此外,肠道免疫细胞还参与到免疫耐受的调节中,即对自身组织的免疫反应进行限制,避免自体免疫疾病的发生。肠道免疫细胞的失调可能会导致免疫炎症反应过度激活,引发炎症性肠病和其他免疫相关疾病。因此,保持肠道免疫系统的平衡和稳定对于整体健康至关重要。
了解和关注肠道免疫系统的功能及其与微生物群的相互作用对于维持整体健康具有重要意义。
主要参考文献
Mowat AM, Agace WW. Regional specialization within the intestinal immune system. Nat Rev Immunol. 2014 Oct;14(10):667-85.
Kayama H, Okumura R, Takeda K. Interaction Between the Microbiota, Epithelia, and Immune Cells in the Intestine. Annu Rev Immunol. 2020 Apr 26;38:23-48.
Allaire JM, Crowley SM, Law HT, Chang SY, Ko HJ, Vallance BA. The Intestinal Epithelium: Central Coordinator of Mucosal Immunity. Trends Immunol. 2018 Sep;39(9):677-696.
Yan Y, Ramanan D, Rozenberg M, McGovern K, Rastelli D, Vijaykumar B, Yaghi O, Voisin T, Mosaheb M, Chiu I, Itzkovitz S, Rao M, Mathis D, Benoist C. Interleukin-6 produced by enteric neurons regulates the number and phenotype of microbe-responsive regulatory T cells in the gut. Immunity. 2021 Mar 9;54(3):499-513.e5.
Cray P, Sheahan BJ, Dekaney CM. Secretory Sorcery: Paneth Cell Control of Intestinal Repair and Homeostasis. Cell Mol Gastroenterol Hepatol. 2021;12(4):1239-1250.
Yang D, Jacobson A, Meerschaert KA, Sifakis JJ, Wu M, Chen X, Yang T, Zhou Y, Anekal PV, Rucker RA, Sharma D, Sontheimer-Phelps A, Wu GS, Deng L, Anderson MD, Choi S, Neel D, Lee N, Kasper DL, Jabri B, Huh JR, Johansson M, Thiagarajah JR, Riesenfeld SJ, Chiu IM. Nociceptor neurons direct goblet cells via a CGRP-RAMP1 axis to drive mucus production and gut barrier protection. Cell. 2022 Oct 27;185(22):4190-4205.e25.

谷禾健康
便秘是世界范围内高度流行的功能性胃肠道疾病,患病率为 2-20%.
根据罗马 IV 标准,慢性便秘的典型症状是排便困难、不频繁或不完全排便。便秘的症状总是连续不断、反复出现,严重影响患者的身心健康和生活质量。


慢性便秘与多种因素有关,如肠道神经细胞的变化、菌群改变、肌病、神经递质和生态失调。此外,部分原因是其久坐的生活方式,情绪、饮食方式和液体摄入偏少,用药物(例如磷酸盐结合剂)和多种合并症(例如糖尿病,甲状腺疾病等),
而长期便秘会使人心情烦躁、容貌衰老,还可能造成肛肠疾患、胃肠道功能紊乱、心血管疾病和性生活障碍等,同时还会增加溃疡性结肠炎、患结肠癌等疾病的患病率。
最近的流行病学研究表明,便秘与许多不良临床结局相关,例如结直肠癌、肝性脑病、乳腺疾病、阿尔茨海默病、终末期肾脏疾病(ESRD),心血管(CV)疾病和死亡率等。
便秘严重影响生活质量,耗费大量金钱与医疗资源,但目前临床治疗效果却不尽如人意,往往停药便复发,因此寻找新的治疗方法或改进现有的治疗方法,可改善患者生活质量与减少社会医疗负担。
临床上认为一周小于3次的排便就叫便秘,尽管很多患者想力求每天一次才舒适。此外,判别便秘的其他特征和症状还包括:
