 国家高新企业 | ISO9001认证
国家高新企业 | ISO9001认证 二级病原微生物安全实验室
二级病原微生物安全实验室- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
国家高新企业 | ISO9001认证 二级病原微生物安全实验室

谷禾健康
http://v.qq.com/s/videoplus/457452244皮肤菌群如何导致痤疮、湿疹?如何维持皮肤健康?_腾讯视频




相关阅读:

谷禾健康
生物系统——组成
生物系统很复杂,具有许多调节功能,例如DNA,mRNA,蛋白质,代谢物,以及表观遗传功能(例如DNA甲基化和组蛋白翻译后修饰(PTM))。 这些特征中的每一个都可能受到疾病的影响,并引起细胞信号传导级联和表型的改变。 除了宿主对疾病的反应调节机制外,微生物组还可以改变宿主特征的表达,例如它们的基因,蛋白质和/或PTM。
生物系统——疾病
为了深入了解疾病的机制,我们需要研究这些特征及其相互作用。例如,黑色素瘤、肺癌和甲状腺癌等癌症是由BRAF癌基因驱动的。然而,当患者接受抑制BRAF的治疗时,往往会产生耐药性。最近的多组学研究揭示了肿瘤特征的异质性和复杂性,如基因突变、转录组、蛋白质和信号通路。现在人们认识到肿瘤可以绕开治疗而产生耐药性。
生物系统——技术
随着下一代测序和质谱技术的发展,人们越来越需要融合生物特征的能力来研究整个系统。转录组、甲基组、蛋白质组、组蛋白翻译后修饰和微生物组等特征都影响宿主对各种疾病和癌症的反应。由于样品制备步骤、测序所需的材料量和测序深度要求,每个平台都有技术限制。近年来,数据集成方法的发展受到了推动。每种方法都使用诸如概念整合、统计整合、基于模型的整合、网络和路径数据整合等方法来具体整合组学数据的子集。
生物系统——多组学
多组学方法的整合使得对疾病病因学有了更深入的了解,例如:揭示微生物组在减轻或增加疾病风险方面发挥作用的各种方式。双酚A(BPA)是一种大规模生产的化学品,广泛应用于食品包装、塑料和树脂中,双酚A的不完全分解就是一个例子。由于双酚a是一种内分泌干扰物,双酚A已成为日益增长的公共卫生问题。因此,利用微生物手段快速、完全降解双酚A等化合物的研究具有重要意义。
本文讨论每个数据特征的研究设计考虑,基因和蛋白质丰度及其表达率的限制,当前的数据整合方法,以及微生物对基因和蛋白质表达的影响。在开发整合多组学数据的新算法时应考虑的因素。
不同生物基因数量
生物系统是具有多种调控功能的复杂生物。 例如,人类基因组由大约32亿个核苷酸组成,可产生20 000至25 000个蛋白质编码基因,并且通过选择性剪接事件可产生超过100万种蛋白质(下图)。
不同的生物有不同数量的基因和蛋白质。例如,在大肠杆菌、酿酒酵母和智人基因组中分别有大约4300、6000和25 000个基因。这导致大肠杆菌、酿酒酵母和智人的每个细胞中分别有大约2400到7800、15 000和300 000个mRNA分子。线粒体转录物约占多聚腺苷酸化RNA的20%。其他高丰度的转录物包括编码核糖体蛋白质和参与能量代谢的蛋白质的转录物。下图概述了人类DNA、DNA甲基化、组蛋白翻译后修饰、mRNA和蛋白质的复杂性。

Graw et al., 2020 Molecular Omics
染色质结构和基因/蛋白质调控的概述。 DNA通路受DNA甲基化和组蛋白翻译后修饰(PTM)的调控。调节的每一层也可以通过环境和宿主生物中存在的微生物进行修饰。 可以通过使用各种核苷酸和蛋白质/肽测序技术对生物调节的每个水平进行测序。
细胞中蛋白质含量
一个细胞中蛋白质的估计数量约为2.36×106(在大肠杆菌中),约为2.3×109(在晚期智人细胞中)。在一个细胞的全部蛋白质总数中,最丰富的蛋白质可占蛋白质含量的5-10%,由核糖体蛋白、酰基载体蛋白(ACP)(在脂肪酸生物合成中的功能)组成,分子伴侣和折叠催化剂、糖酵解蛋白质(能量和碳代谢的主干)和肌动蛋白等结构蛋白质。
转录因子是一种低丰度的蛋白质,在细菌中每个细胞的拷贝数为1-103,在哺乳动物细胞中为103-106。
最丰富的蛋白质通常在细菌中有数千个拷贝,在哺乳动物细胞中有数百万个拷贝。由转录因子调控的基因数量取决于其浓度。蛋白质含量取决于生长条件和基因诱导。最后,考虑到微生物与宿主细胞数量的比例(取决于宿主细胞类型)和其他因素,这可能会变得更加复杂
mRNA 和蛋白质寿命以及差异
由于仪器检测,动态范围和分子寿命表达的限制,用于各种组学平台的测序技术只能捕获某一时刻某个细胞群体中发生的情况的快照。 例如,mRNA转录本和蛋白质的终生表达差异很大。 在大肠杆菌中,mRNA的中位寿命为5分钟,在发芽酵母中为20分钟,而对于人参则为600分钟。然而,蛋白质的寿命约为1-2天。
转录和翻译的速率因生物体的不同而不同(大肠杆菌:每秒10-100个核苷酸(nt)和10-20个氨基酸(aa)/ s。智人:6-70 nt / s和2 aa per s;分别为转录和翻译速率)。
对于大肠杆菌来说,一个单一的mRNA转录本在被降解之前可以产生10-100个蛋白质。鉴于这一信息,我们可以看到,将我们对组学平台的选择和由此产生的对细胞过程的解释相结合,检测具有更长寿命的蛋白质的机会将增加。
在考虑数据整合研究设计、开发新算法和解释结果时,认识生物体的生物复杂性、分子的动态范围、测序限制以及这些分子的表达寿命非常重要。
近年来,微生物组学在宿主健康中的重要性已得到公认。全生物和全基因组的概念对我们如何看待微生物组有着深远的影响,尤其是在治疗方面。这种微生物-宿主相互作用的密切关系可以更明确地称为“微生物群-营养代谢-宿主表观遗传轴”。微生物与宿主相互作用的紧密关系可以更明确地称为“微生物群-营养代谢-宿主表观遗传轴”。
微生物群及其代谢产物可以通过直接修饰组蛋白,改变DNA甲基化谱图和影响而影响宿主表观遗传。 非编码RNA的性质(上图)。 例如,可以通过改变组蛋白修饰酶的活性和酶底物的水平,通过微生物群来修饰组蛋白。
微生物影响药效
微生物群也可以影响药物的治疗性质。 许多前药,即必须进行代谢转化才能在药理上有用的药物,可能会保持无活性(即不存在介导前药向其活性形式转化的微生物群),或者该药物/前药可能无法生物利用。此外,服用NSAIDs(非甾体类抗炎药)的患者可能会促进抗生素耐药菌的优势,因为24%的非处方非处方NSAIDs被抑制。
这些代谢组学效应引起人们对旨在用于人类和农业系统的治疗药物或其他饮食和治疗方案的潜在副作用的担忧。 例如,抗生素可以消除产生组蛋白脱乙酰基酶(HDAC)抑制剂的微生物。 这些微生物(如果存在)可以增强调节性T(Treg)细胞,从而有助于抗炎过程。
微生物代谢途径的多样性及其对药物药代动力学和药效学的影响可能部分解释了个体和人群之间药物反应的变化。 因此,涉及微生物组的治疗方法可能必须因地制宜。组蛋白可以同时进行变体置换和翻译后修饰(PTM),这些共同构成了“组蛋白密码”。 这些局部排列可以影响染色质结构,从而导致转录活性的激活或抑制。
通过饮食,微生物有能力改变宿主的甲基化和PTM谱,并且还可以通过膳食碳水化合物的发酵影响短链脂肪酸(SCFA)的生成。丁酸盐和乙酸盐等SCFAs可抑制脱乙酰酶水平。这意味着由于乙酰化促进转录活性的增加,染色质结构变得越来越松弛。事实上,已经证明微生物可以以位点特异性和组合方式影响宿主组织乙酰化和甲基化染色质状态,甚至影响宿主发育和代谢表型。
微生物参与干预
未来关注健康医疗策略时,越来越多地考虑对微生物组的发展及其相应的宿主个体发育变化进行建模。考虑到宿主免疫系统不仅必须能够识别“自身”抗原,而且还必须能够识别共生微生物的抗原,这些变化可以通过宿主免疫成熟来证明。
微生物如何影响主要组织相容性复合体(MHC)的表达,或者宿主杂合度如何通过MHC影响微生物群的多样性,这在很大程度上是未知的,也是一个活跃的研究领域。微生物在癌症和免疫治疗中的作用正日益成为治疗策略发展的目标。蛋白质组学与其他组学策略相结合已被用于研究疾病过程。如果我们不考虑微生物群的影响,那么我们可能会错过开发潜在治疗方法的有意义的见解。尤其是那些与代谢紊乱(如肥胖)或代谢物(如胆汁酸)对器官系统的全身影响有关的疾病。
微生物生态学的历史围绕着适当的系统发生标记基因的测序和比对。 WoSes and Fox(1977)首先将16S rRNA基因用作标记基因,是迄今为止最常用的标记基因,其大规模数据库包含从环境和培养来源(例如SILVA,RDP,Greengenes)获得的全长基因分离株 )(表1)。 新的微生物分类数据库,例如基因组分类数据库(GTDB),不仅建立了16S rRNA基因参考数据库,而且还利用系统基因组学信息提供了一个一致的框架,用于确定从元基因组获得的系统发育背景部分或完整基因组 。
大数据集的可用资源列表

选择合适的引物和平台
除了选择标记基因和合适的数据库外,研究人员还可以在测序方法和平台之间进行选择。 由于Illumina和Ion Torrent等短读平台的局限性,研究人员必须在〜1500 bp的16S rRNA基因的可变区之间进行选择。 取决于微生物群落组成,每个可变区提供不同水平的敏感性和特异性。 然后选择在研究中最能区分普通分类群的引物组和扩增子区域的组合。
目前针对扩增子测序可选择的测序平台和方案很多,不同平台的读长和适用的测序区段以及优势各有不同。16s测序主要的测序区段包括v4、v3v4,v1v2,v6,此外还有全长等不同的区段选择,不同可变区或全长由于引物的不同以及不同种属相应区段内的变异多样性差异,对菌属的丰度评估会有一定的差异。
从长度来看,全长16s长度为1.5kb左右,单菌落的16s全长sanger一代测序仍然是菌种鉴定的主要手段,纳米孔和pacbio的三代测序可以高通量的获得全长序列,对于希望更高分辨率的分析菌种的研究有一定优势。三代的测序准确度目前逐渐改进,直接测序准确度可以在90%以上,纠错后可以提高到97~99%以上,已足够提供高精度的分类。三代目前主要问题在于建库成本相对较高,通过使用barcode可以降低部分但仍然偏高,此外普遍测序深度相对于二代测序要低许多。
目前最主要的可变区选择是v4区和v3v4区,v4区长度为256bp左右,加上两侧引物长度为290bp左右,使用双端2x250bp或2x150bp可以测通,此外如454、life、illumina的测序平台读长也可以主要涵盖该区段读长。例如采用illumina Novaseq测序平台对该项目进行双端测序(paired-end),测序得到了fastq格式的原始数据(样本对应一对序列s_1.fastq和s_2.fastq)。再配对拼接成单条序列。其引物通用性相对是所有可变区中最高的,大量的大规模菌群调查研究都采用v4区作为检测区域,包括人体菌群研究如:hmp,肠道菌群如美国肠道计划agp,欧洲的fgfp等,以及全球土壤菌群调查,目前仍然是国际研究中使用最广泛和认可的检测区域。
illumina的miseq提供了长达2x300bp以及hiseq2500和最近的novoseq提供有2x250bp的测序方案,为进一步利用读长,目前有相当一部分研究选择v3v4区,该区段长度在460bp左右,相较于v4度多出了v3区段约100bp左右的片段,在少部分菌属中可以增加一定分辨率。经过对比,v3v4区的检测结果和v4区在绝大部分菌属中的丰度一致,但由于引物不同,在少量菌属中丰度会有不同偏向,v3v4从otu层面上并未发现较v4区有明显增加。引物的选择和提取、储存方法是影响菌群检测丰度构成的主要因素,不同研究之间的比较需要考虑到实验方案的一致,相同的方案可以直接比较。
当前的宏基因组分析技术已使研究人员能够从环境/宿主来源的样品中获得足够的序列覆盖率,从而获得部分和完整的基因组草图。 该覆盖因子高度取决于物种的均匀度和丰富度。 还可以通过拼接组装元基因组组。 但是,由于难以组装和正确分装高度保守的基因(如核糖体亚基基因),因此它们通常必须使用浓缩的通用蛋白将这些基因组置于系统发育背景中。
组合的通用标记基因被用来构建由环境和寄主衍生序列组成的基因组以及来自培养物收集的少数常见微生物基因组的系统发育。微生物基因组测序的热潮使得有必要构建易于使用的软件包以及分析工具,以帮助生物学家学习如何对其全部或部分的元基因组数据进行分析。这类工具的例子如,QIIME 2、metaWRAP、 Sunbeam、SqueezeMeta、metAMOS、 mg RAST、IMG/M、 Anvi’o、MicrobiomeAnalyst、以及biobakery集合中的各种工具(例如MetaPhlan2、PhyloPhlan、HUMAnN、LEfSe)等。
此外,如果深入研究,还需要望整合疾病指标、宿主蛋白质组学和微生物多样性多组学的联合分析。
根据生物学问题的不同,有许多类型的组学技术,针对DNA、总RNA、mRNA、miRNA、DNA甲基化、蛋白质、蛋白质修饰、组蛋白翻译后修饰、宏基因组学、宏蛋白质组学,测序平台经过多年的改进,现在可以在几天内从少量材料中对大型复杂人体样本进行测序(表2)。已经开发了几种工作流程来对整个基因组、整个外显子组(DNA的蛋白质编码部分)和转录组(mRNA)进行排序,并对特定的癌症或免疫相关基因进行排列。此外,还可以利用亚硫酸氢盐全基因组测序或Illumina的甲基化珠芯片阵列分析修饰,如DNA甲基化。 还可以通过牛津纳米孔技术(ONT)MinION平台和PacBio仪器对长读的DNA和RNA进行直接测序来确定此类修饰的检测。
推荐覆盖率和读数

基因组测序
DNA测序技术的错误率和读取长度各不相同。Illumina短读测序(即Hiseq、Miniseq等)通常具有非常低的错误率,约为每碱基0.25%,但对低多样性文库敏感,如16S宏基因组学和靶向基因方法等应用。长读取技术的错误率较高,PacBio为13–15%,Oxford Nanopore instruments为5–20%。Illumina平台的读取长度最大为600个碱基,但长读取技术通常一次读取可达到10–30 kb。最佳读取长度也取决于应用程序。
大多数测序实验可以收集150-300碱基对读取长度的合适信息,但也有例外。对于全基因组测序(WGS),最长的读取可能是最佳的,但是对于长读取技术,错误率随着长度的增加而增加。有许多研究者把“短读”和“长读”结合起来。由于最近长读取排序技术的出现,关于WGS以外应用程序的最佳长读取长度的信息非常缺乏,但Illumina short read sequencing提供了丰富的最佳读取长度建议。
蛋白质测序
在过去5-10年中,质谱仪通过增加测序深度能力也得到了改进。这项技术已经从使用旧的LTQ质谱仪在细胞系实验中对大约3000个蛋白质进行测序,发展到使用新的Orbitrap Lumos和Orbitrap Eclipse质谱仪对8000-10000个蛋白质进行常规测序。大多数蛋白质组学实验都是采用数据相关采集(DDA)模式进行的。在该方法中,选择从液相色谱(LC)柱洗脱的MS1扫描中最丰富的前20个肽在orbitrap中进行裂解,以产生肽序列MS2扫描。样品混合物的复杂性极大地影响了测序深度和将鉴定多少蛋白质。了解样品的蛋白质丰度和组成是至关重要的。如果转录因子是目标分子,那么在质谱分析之前去除高丰度蛋白质的方法可能是必要的。这对于含有大量分子(如白蛋白和血红蛋白)的血清和血浆样品尤其重要。否则,质谱仪将测序数千个白蛋白分子,并错过最有趣的低丰度蛋白质。
最新的质谱技术利用数据独立采集(DIA)来对MS1扫描中所有肽从LC色谱柱洗脱时的序列进行测序,这与仅对最丰富的峰进行测序的DDA方法相反。
对于复杂的混合物,例如上面的血清示例,DIA方法优于DDA。 这种方法有助于克服受高丰度蛋白质高度影响的复杂混合物。
除了对宿主基因和/或蛋白质进行鸟枪法测序外,我们还可以对微生物组利用鸟枪法测序。
当测序深度很浅时,弹枪宏基因组学/元代谢组学只能采样优势菌群。shot弹枪对微生物组测序的主要挑战是由于采样不足而难以组装基因组片段,将肽组装在一起以进行可靠的蛋白质和生物分类鉴定也同样困难。
尽管存在这些潜在问题,但从各种人体部位和疾病(如唾液、肠道/粪便、颈阴道疾病或慢性肾脏疾病)中对微生物蛋白质组进行深度取样是可能的。然而,每个研究必须考虑的研究/取样设计和分析方法可能有很大差异。
从差速离心到双过滤差速分离,几种样品制备方法已被证明能富集微生物生物量。这些方法通常遵循各种优化的微生物裂解方案,通常涉及机械破坏(如打珠、超声波),辅以酶(如胰蛋白酶)和洗涤剂。在成功溶解后,同样重要的是去除残留的酶、洗涤剂和盐。
元蛋白质组学实验的另一个复杂性是由于同一生物体内的蛋白质具有共享的肽序列这一事实。 为了对蛋白质鉴定有信心,应以高可信度鉴定蛋白质的独特肽段匹配。 当将肽序列映射到数百个具有保守蛋白序列的不同物种时,这变得更加复杂。 质谱法不对蛋白质进行测序,而是测量肽的电荷,并依靠与蛋白质序列数据库匹配的质谱进行蛋白质鉴定。
精心挑选的数据库对于正确分析从这些各种测序平台生成的核苷酸和蛋白质测序数据至关重要。 使读数与参考基因组比对的能力仅与参考基因组中存在的序列和注释信息一样好。 有几种资源可以不断地整理和更新核苷酸序列信息和注释,包括加利福尼亚大学圣克鲁斯分校(UCSC)基因组学研究所基因组,美国国家生物技术信息中心(NCBI)GenBank和RefSeq,DNA元素百科全书(ENCODE)和Ensembl 仅举几例。 通用蛋白质资源(UniProt)包含Swiss-Prot(手动注释和审阅)和TrEMBL(自动注释且未审阅)数据库,以获取蛋白质序列信息。
已经开发了几种数据集成方法来集成某些类型的组学数据。 另外,已经创建了大数据存储库来存储来自各种疾病的测序实验的数据。 这些资源提供了有价值的构建基块和大量生物样本,可用于推动数据集成方法的发展。 当前,数据集成工具实现了多种方法,但通常分为两类:多阶段分析和元维度分析。
多阶段集成模型仅使用数据的两个数字或分类特征构建。 例如,将来自RNA-seq实验的基因计数与来自质谱运行的蛋白质信息相结合。元维度分析试图通过级联或转换将所有感兴趣的数据类型合并到可以同时分析的同时矩阵或“元数据”集中。
后一种方法具有更大的统计能力,但在尝试合并来自不同类型数据集的数据时可能会具有挑战性。 但是,研究人员如何确定最合适的工具或方法?
如上所述,生物学问题是选择的分析方法类型的驱动力,诸如采样,平台类型和数据质量等因素很重要。 样品如何收集和准备?
如果测序深度或质量较低,是否可以有效分析数据? 数据类型兼容吗?
归一化和滤波后损失了多少信号?
这些都是在选择适当工具之前应考虑的所有问题。
不幸的是,数据集成和分析非常复杂,并且对于具有有限生物信息学背景的研究人员而言,目前还没有许多用户友好的工具。 许多工具使用统计语言R,除了强大的生物统计知识外,它还需要专业编程知识。 例如,将蛋白质组学,转录组学和途径分析结合到两个数据集上的R包积分学使用了相关分析和偏最小二乘回归。R包mixOmics使用多元分析进行数据探索,降维和可视化。 通过途径分析,iClusterplus和LRACluster进行的miRNA和基因表达使用聚类来整合甲基化和基因表达数据。
多组学数据集成工具



对于多状态和多维方法,都使用了许多不同的算法,但最常见的算法是聚类,网络分析,数据约简(PCA)和贝叶斯分析。Ray等2014年使用贝叶斯分析,使用从癌症基因组图谱项目收集的数据分析卵巢癌中的基因表达和甲基化数据,并检测到一个基因SPON1,该基因似乎受其CpG位点的甲基化调控。当缺乏生化相互作用的先验知识时,基于相关性的分析是有用的。无论采用何种方法,适当的规范化和数据过滤是非常重要的,因为数据来自多个来源。
还有一些基于网络的工具,如Paintomics,试图使数据分析更容易,但对于缺乏经验的用户来说仍然很困难,研究人员必须对他们的数据有很好的工作知识。此外,还有一些数据库常用于综合组学分析,如癌症细胞系百科全书(CCLE)、癌症基因组图谱计划(TCGA)、与基因组学驱动治疗相关的肿瘤改变(TARGET)和组学发现指数(OmicsDI)。CCLE和TCGA已经描述了数以千计的癌症数据集,可以用于数据挖掘和可视化。TARGET利用临床信息并在其网站上提供分析工具的资源。omicdi提供了一个平台,用于搜索各种生物的公共和受保护数据。
对于任何高质量的研究,进行多组学研究应该首先确定研究的范围和限制。仔细的计划和执行将提高研究的稳健性和可重复性,在多组学研究中尤其重要,因为它们涉及大量的比较、定制的统计分析、大量的财力,时间和精力。一旦研究假设被明确定义,选择一个合适的研究设计,最好地解决研究假设。因此,有几个问题需要评估,
例如:是否有一个或多个干预组与对照组(或其自身)进行比较,或者是否在干预前后对同一样本的效果进行评估?
干预效应是在一段时间内产生的,还是在几个不同的时间点测量样本?
生物样本是否会被单独收集或分析?它的科学依据是什么?
哪些类型的组学平台将提供最有价值的以及如何整合多组学数据?
来自同一生物来源的样本是否可用于所有感兴趣的多组学平台?
理想情况下,所有omic平台的样本将从同一来源收集。
然而,由于样品的特殊限制或材料的可及性和数量,这并不总是可能的,从福尔马林固定石蜡包埋(FFPE)组织生成多组学数据对于某些组学平台可能是不可能的。虽然在选择实验设计时有许多问题需要考虑,但选择研究设计的决定因素通常是其可行性和经费限制。
样品和数据的收集应以数据分析为指导,以减少混淆和技术因素,例如批量效应。这些效应可以在样品收集,制备和存储的步骤中引入。
由于与多组学研究相关的数据的复杂性和大量数据,因此针对特定的研究项目量身定制统计分析至关重要。已提出了多种集成多组学数据的方法,并将其归类为受监督的, 半监督或非监督; 以及基于概念,统计,相关性,网络和模型的集成。
一项研究的统计能力取决于几个因素(下图),其中一些因素可以控制,而另一些因素由于研究及其设计而固定。首先,选择了用于分析的统计方法。虽然有些测试比其他测试更强大,但重要的是验证和满足他们的假设。
影响研究统计能力的另一个因素是单个组学平台测量的变量数量,通常由组学平台决定。例如,基因组学通常测量数百万个变体,转录组学量化了成千上万个分子,和蛋白质组学和代谢组学分析了数千个分子。此外,统计效力受表型或处理效应的大小和差异程度(效应大小)的影响。效果有多明显?组间的信号差异有多大?有多少被测变量受到影响?关于效应大小的信息可以从以前的文献或专家知识中获得,但通常是未知的。
这种情况下,初步研究可以帮助估计效应大小,但由于不稳定,这些估计需要谨慎处理。另一个效力影响因素是测量值的均匀性,描述了样品的自然方差、测量仪器的精度和检测限。随着方差的增大,统计效力将减小。样本的方差可能是多方面的结果,例如样本群体的选择、组织类型的选择或混杂因素。
除了样本方差膨胀外,混杂因素也会在数据中引入偏差,因此,收集样本元数据以减轻某些混淆的影响是很重要的。由于影响研究统计能力的大多数因素是固定的或由研究设计决定的,因此最常用于调整研究统计能力的因素是样本量。

多组学研究中影响统计功效的因素
Graw et al., 2020 Molecular Omics
研究的首要考虑正在调查的疾病或研究问题的背景,以及整合在一起时,什么类型的数据将提供有价值的见解。根据生物学问题、材料类型(新鲜组织、FFPE组织、血清/血浆和细胞系)、DNA/RNA/蛋白质的数量、生物复制的数量以及研究中混杂效应的数量,这些因素将决定数据采集所需的最佳样品制备和测序方法。
样品制备方法,包括每个样品制备的日期、提取的DNA、RNA和/或蛋白质的类型、基因组学的文库生成、质谱分析的蛋白质消化和肽标记方法以及测序平台/仪器,都是研究设计和最终结果解释的关键因素结果。
如果一个样本是在不同的日期制备的,而不是其他生物复制品,这将引入方差和/或偏差,并降低分析的统计能力。如果蛋白质组样品使用多个TMT-10plex批次进行复合,这将在整个测序过程中引入批次效应。这些因素应在样品制备前进行讨论。
同样重要的是要知道什么样的调控特征被捕获用于测序和整合。例如,如果在进行质谱分析之前在样品制备过程中膜蛋白没有溶解,那么膜结合蛋白就不能与基因表达数据整合。质谱数据的一个警告是,缺失值并不一定意味着蛋白质没有表达,只是蛋白质低于质谱仪的检测限。生物学问题应该成为多组学数据整合方法的驱动力。
在大多数情况下,当前的工具利用聚类、网络、数据简化和贝叶斯分析。随着数据获取量的不断增加,产生了大量的数据集,使得机器学习对于有效的分析和数据挖掘变得越来越必要。有必要使用易于获取和记录良好的方法、工具和算法。
机器学习在允许科学家集成多组学数据集方面发挥了越来越重要的作用。通过利用机器在大量生物数据中比较和识别模式的能力,可以用更加准确和有效的方法来阐明复杂的细胞机制,在某些情况下还可以预测临床结果。这是通过计算机独特的能力来实现的,它可以同时观察多个层次的组学数据,从而提供一个更全面的系统视图。
尽管多组学数据集可以为个体提供更深入的理解,但这并非没有成本。组学研究通常依赖于大量的比较、正确的数据类型、适当的统计分析以及大量的时间、技术人员和金钱投入。在构建一个实验时,人们必须清楚什么类型的组学数据可以而且应该被整合以获得对所研究系统的最大理解。
高通量的组学平台并不总是回答研究问题所必需的。传统技术:如酶联免疫吸附试验(ELISA)、免疫组织化学(IHC)和定量聚合酶链反应(qPCR),也是验证特定生物学机制所必需的。事实上,为了验证从组学数据中鉴定出的重要分子是一个真正的阳性结果,通常需要这些技术来验证一个更大的组学研究的结果。
但是每种方法都受到其统计能力、样本量、技术变量、批次效应、测序深度、样本制备和许多其他因素的限制。在设计、进行和分析研究以及解释研究结果时,必须牢记这些因素。因此,如果允许,建议研究设计一开始就让生物统计学家/生物信息学家参与进来。
参考文献:
Graw S, Chappell K, Washam CL, Gies A, Bird J, Robeson MS 2nd, Byrum SD. Multi-omics data integration considerations and study design for biological systems and disease. Mol Omics. 2020 Dec 21. doi: 10.1039/d0mo00041h. Epub ahead of print. PMID: 33347526.
A. Zaman , W. Wu and T. G. Bivona , Targeting Oncogenic BRAF: Past, Present, and Future, Cancers, 2019, 11 , 1197
A. Alvarez-Arenas et al., Interplay of Darwinian Selection, Lamarckian Induction and Microvesicle Transfer on Drug Resistance in Cancer, Sci. Rep., 2019, 9 , 9332 .
K. Yu et al., An integrated meta-omics approach reveals substrates involved in synergistic interactions in a bisphenol A (BPA)-degrading microbial community, Microbiome, 2019, 7 , 16.
G. D. Poore et al., Microbiome analyses of blood and tissues suggest cancer diagnostic approach, Nature, 2020, 579 , 567 —574 .
A. Gonzalez et al., Characterizing microbial communities through space and time, Curr. Opin. Biotechnol., 2012, 23 , 431 —436 Search PubMed .
D. Gurwitz The Gut Microbiome: Insights for Personalized Medicine, Drug Dev. Res., 2013, 74 , 341 —343 .
N. Issa Isaac et al., Metaproteomics of the human gut microbiota: Challenges and contributions to other OMICS, Clin. Mass Spectrom., 2019, 14 , 18 —30

谷禾健康

感到疲惫?乏力?食欲不振?容易流血?皮肤发痒?腿肿?体重减轻?…这些有可能是慢性肝病的信号,慢性肝病是一个主要的健康问题,它可以发展为肝硬化和肝癌。
肝病按照病因划分,常见的有如下几大类:
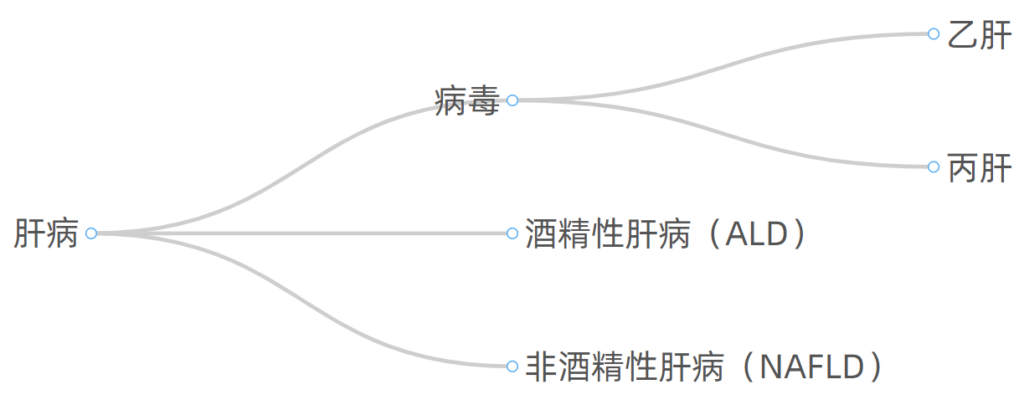
无论病因如何,肝病如果没有得到有效管理控制,进展的常见终末阶段是肝硬化。
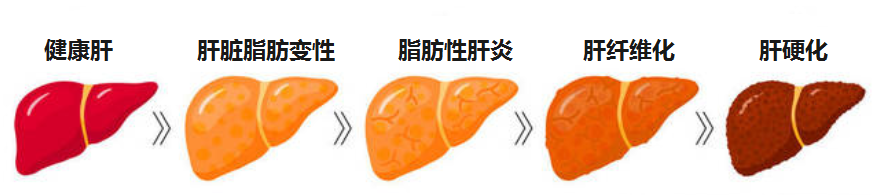

肝硬化可导致失代偿和肝细胞癌的发展。
发病率
目前由于疫苗接种计划和有效抗病毒治疗不断发展,乙肝和丙肝肝硬化的发病率正在下降,但由于以下两个原因导致另两种肝病发病率不断上升:
1、肥胖,导致非酒精性脂肪肝的发病率增加
2、饮酒,导致酒精性肝病的发病率增加。
本文带你详细了解肝病与肠道菌群:
肝脏也是胆汁产生的场所,在肠肝循环中具有重要作用。
肝脏,作为第一个接触微生物产物进入门静脉循环的器官,可能受到肠道微生物群及其变化的多种影响。肠道微生物群的成分或代谢物通过各种机制与肝脏相互作用。
肝脏中产生的胆汁酸(如胆酸和鹅去氧胆酸)与葡萄糖醛酸酶结合。一旦分泌到十二指肠肠腔,肠道菌群代谢和解共轭,产生尿胆原(排泄)和初级胆汁酸被吸收和循环。胆汁酸还通过内在的微生物调节活动来控制和影响菌群。
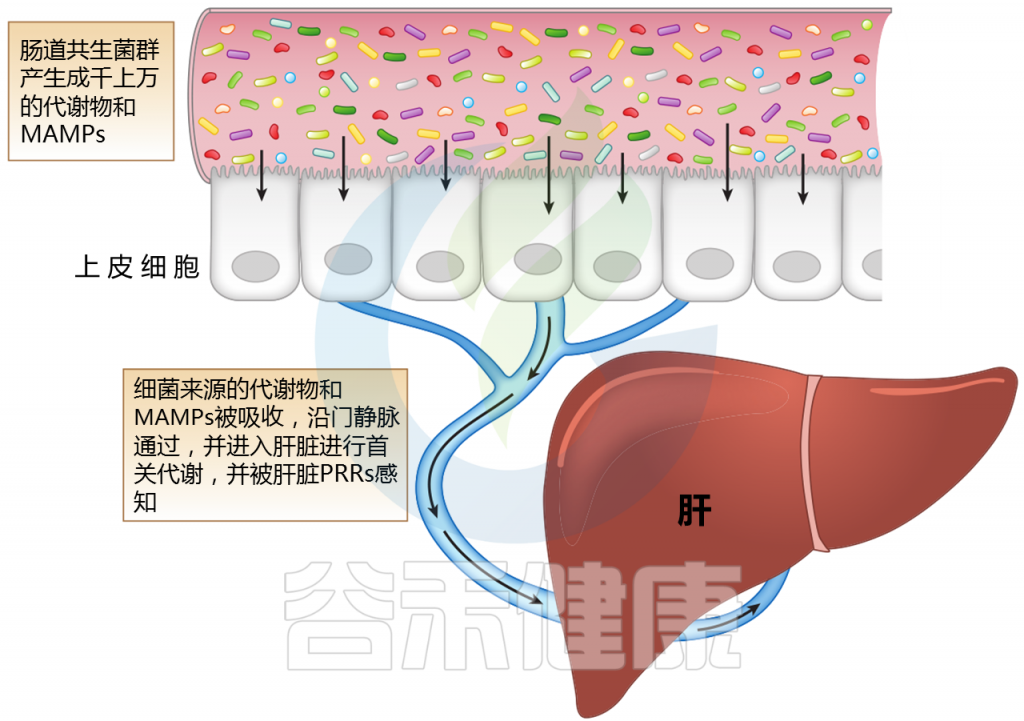

Jones R M,et al.,Annual Review,2020
注:MAMPs——微生物相关分子模式;PRR——模式识别受体
肝脏位于体循环和来自肠道吸收活动的大量外源性分子和微生物相关分子模式(MAMPs)之间的界面。
微生物产生的代谢物会随着微生物群的组成而变化;它们沿着门静脉分流并引发多种信号通路的激活。MAMP暴露过多会导致炎症。
肝硬化前期肠道菌群总体变化:
整体多样性减少,厚壁菌门减少,拟杆菌门和变形菌门增加。
在三种类型的肝炎中,肝硬化前期微生物参与非酒精性脂肪肝病和酒精相关性肝病的证据相对更为有力。
—— 非酒精性脂肪肝病
非酒精性脂肪性肝病(NAFLD)和非酒精性脂肪性肝炎(NASH)是一种以肝细胞内脂质(主要是甘油三酯)的微泡或大泡状积聚为特征的肝病,并伴有不同程度的炎症浸润。
在非酒精性脂肪性肝病中,研究人员发现粪便、血液和肝活检中的致病菌(如肠杆菌科和大肠杆菌)的相对丰度增加。随着病原菌数量的增加和厚壁菌门数量的减少,菌群失调。
—— 酒精性肝病
长期摄入酒精可导致人类和动物模型中菌群结构的变化,肠道通透性增加,随着持续的酒精滥用,致病菌的多样性进一步减少,相对丰度增加,如肠杆菌科和肠球菌科。
在小鼠中,乙醇可以降低肠道屏障功能,并允许增加局部炎症。从机理上讲,乙醇也能抑制抗微生物凝集素的表达,在慢性乙醇消耗模型中,无Reg3β和Reg3γ的小鼠表现出增加的细菌移位和NASH的发生。这些发现表明乙醇可能介导了肝脏潜在的促炎性刺激,有多个长期后遗症。
尽管肝硬化的病因可能有所不同,但在肝病的末期,微生物群与肝脏的相互作用在很大程度上与病因无关。也就是说无论哪种都可能发展为肝硬化。
前面我们知道,肝炎如果不能得到有效控制,久而久之会对肝造成永久性损伤,也就是肝硬化。肝硬化又分为两种,一种是代偿性肝硬化,一种是失代偿肝硬化。
代偿性肝硬化虽然肝功能减退,但机体仍能正常运行。
失代偿性肝硬化指肝硬化发展到一定程度,出现肝功能衰竭,可能出现严重并发症,如肾功能衰竭、静脉曲张出血和肝性脑病等。
肝硬化患者最初没有症状,等到了肝硬化失代偿(定义为腹水、肝性脑病、黄疸和/或胃肠道出血),就是肝硬化过程中的转折点。
——肝硬化失代偿期会发生什么?
代偿性肝硬化患者可能会保持多年稳定,死亡率非常低,每年< 1%。然而,在第一次住院治疗后,35%的患者将在28天内出现后续的失代偿,需要频繁入院。肝硬化急性失代偿可导致慢加急性肝衰竭(ACLF)。
注:慢加急性肝衰竭是一种综合征,以一个或多个主要器官或系统(肝、肾、脑、凝血、循环或呼吸)衰竭为特征。
慢加急性肝衰竭是失代偿性肝硬化的主要死亡原因。他们的慢加急性肝衰竭(ACLF)的中位时间为2-3年,死亡率每年约为10%。


Trebicka Jonelet al, Nat Rev Gastroenterol Hepatol,2020
每次失代偿后,慢加急性肝衰竭在28天内的发展约为30%,在此阶段死亡率在28天内攀升至40%。这些器官衰竭患者通常在重症监护室接受治疗,肝移植是唯一的治疗方法。
肠道菌群如何影响肝硬化失代偿?
肝硬化中微生物群-肠道-肝脏相互作用
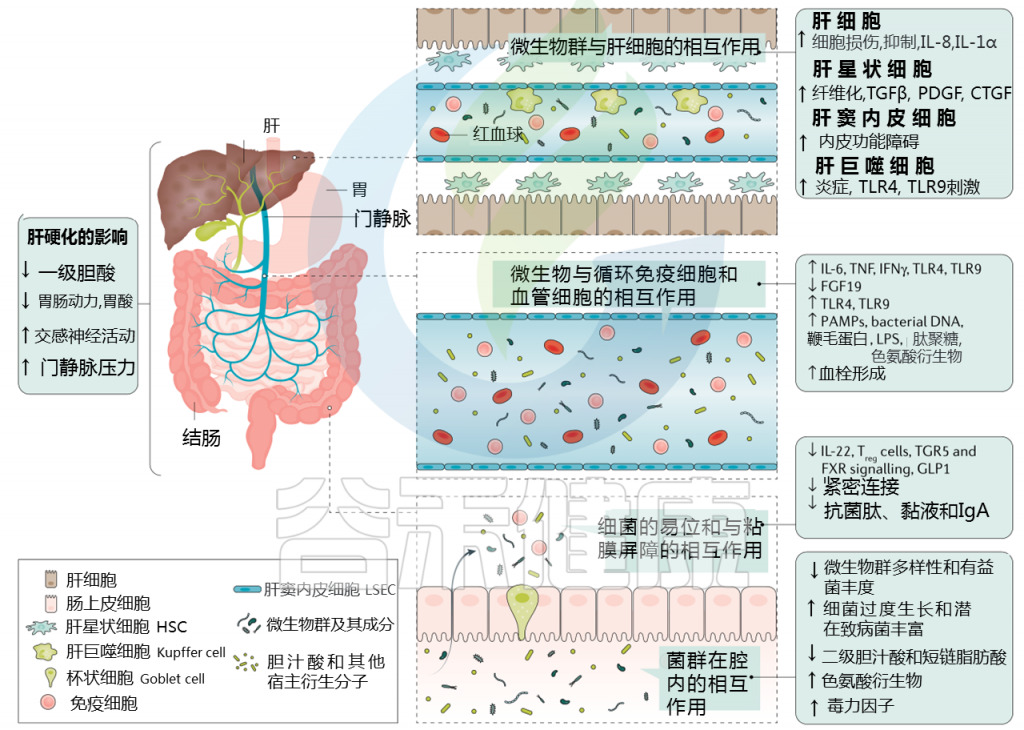

Trebicka Jonelet al, Nat Rev Gastroenterol Hepatol,2020
肝硬化发展过程中肠道微生物群的变化在很大程度上归因于肠道微生物群组成及其产物,产物可能具有致病作用。短链脂肪酸(SCFAs)、次级胆汁酸和色氨酸衍生物增加,而粘液蛋白的变化、功能失调的紧密连接和抗菌肽的减少促进了微生物组成分和/或其代谢物(病原体相关分子模式(PAMPs))跨肠屏障的移位。
接下来,就肠道菌群与肠道屏障,病变肝脏的相互作用,菌群代谢产物与转运,门静脉高压和酒精的作用这4个方面展开讨论。
1. 与肠道屏障相互作用
肠漏:
一个不正常的微生物群可以影响肠道上皮屏障,并导致内容物从肠道到肝脏和其他地方难以控制的转移。这种肠上皮屏障的损伤被称为漏肠。
在肝脏疾病期间,由于肝肠循环、肠道炎症和门脉高压的改变,宿主生理和肠道完整性发生重大变化。与此同时,肠道微生物组和肝脏之间的串扰也发生了重大变化:
肠道中的宿主粘膜蛋白和通路(例如FXR信号)被肠道微生物群代谢物(例如短链脂肪酸)改变,并可能导致肝脏损伤;
肝脏疾病进展引起的肠道内化变化可能会影响肠道屏障,促进肠道炎症,降低抑菌肽;
而肠道相关淋巴组织可能通过驱动肠道炎症,从而增加肠屏障的通透性参与肠屏障的功能障碍
由于穿过肠道上皮屏障的细菌成分会引发免疫反应,因此肠道渗漏被认为会增加慢性全身炎症。例如,在肝病患者,尤其是肝硬化患者中,细菌细胞壁脂多糖(LPS)水平升高,也被称为内毒素血症。
2018年的一项研究表明,从失代偿期肝硬化患者的循环血和其他“无菌”隔室中分离出的细菌是可行的,这表明即使是活的细菌也可以在失代偿期肝硬化期间通过肠屏障转位。
2. 与病变肝脏的相互作用
从失代偿到慢加急性肝衰竭的进展与全身炎症的广泛激活有关,影响许多细胞因子和炎症系统。
释放危险相关的分子模式来促进失代偿和炎症
慢加急性肝衰竭的发展,不同衰竭器官的死亡细胞和细胞外基质的重塑,特别是在肝脏,都通过释放危险相关的分子模式来促进失代偿和炎症,最终可能是慢加急性肝衰竭发展的先决条件。
这种联系在乙肝病毒引起的肝硬化和/或慢加急性肝衰竭患者中尤其常见。慢性乙肝病毒的再激活可能导致肝损伤,从而释放危险相关的分子模式,进而导致器官衰竭和慢加急性肝衰竭。
在慢性肝病和肝硬化患者中,甲肝病毒和戊肝病毒重叠感染也可能诱发慢加急性肝衰竭。
对病原体相关分子模式的免疫反应导致器官功能障碍
另一方面,这种炎症的爆发是由微生物或其成分的转移引起的,也称为病原体相关分子模式。对病原体相关分子模式的免疫反应可能在免疫病理学过程中导致器官功能障碍,在这种过程中,免疫反应会导致感染造成损害。
炎症标志物水平升高
血浆中全身性炎症标志物(如IL-8或IL-6)水平升高,无论是否有明显的细菌感染,都与肝硬化和慢加急性肝衰竭的急性失代偿有关。新的数据强化了改变后的肠道微生物群和慢加急性肝衰竭之间的联系。
血浆代谢物特征
除了肠道菌群衍生代谢物与慢加急性肝衰竭发展之间的关联之外,血浆代谢物特征(可能也来自肠道菌群)也与全身炎症和慢加急性肝衰竭密切相关。
这些发现可能解释了为什么肠道微生物群的改变和细菌易位的增加可能为不同器官的免疫病理发展为器官衰竭,加重全身炎症和诱导ACLF的环境。
酒精性肝病
2019年研究表明,乙肝病毒相关ACLF患者的循环细菌DNA显著增加,并与炎症标记物(如CXCL10,一种已知的趋化因子)相关,在非乙肝病毒病因中也与ACLF相关,尤其是酒精性肝硬化。
另外,在急性酒精性肝炎(ACLF的一个亚型)中,研究表明,与没有这种特殊菌株的患者相比,具有一种产生细胞溶血素的肠球菌菌株的患者具有极高的死亡率(180天内的死亡率分别为89%和3.8%)。
我们知道,急性酒精性肝炎与微生物变化密切相关,这可能决定患者的临床特征及其结果。在几乎一半的急性酒精性肝炎患者中,失代偿和慢加急性肝衰竭都会发生,但没有任何可识别的促发事件,这表明这与内源性机制有关(例如,门静脉高压症或细菌易位)。
3. 门静脉高压症和酒精的作用
即使肝硬化的门静脉高压症得到有效治疗,例如通过放置经颈静脉肝内门体分流术(TIPS)支架,许多患者仍出现进一步失代偿和ACLF(2年内约47%),尤其是由于全身炎症和随后的器官衰竭。
此外,与肝静脉、右心房和外周静脉血相比,腹水的存在决定了门静脉循环微生物群的组成(腹水:一种失代偿的指标),以及与炎症标记物和ACLF的发生相关的特定循环微生物群。
然而,应该指出的是,TIPS支架本身可能会加重高动力循环,从而导致内毒素水平的增加。从长远来看,TIPS支架降低了失代偿率,可能主要是由于这些肝硬化患者腹水的发生率降低。
失代偿性肝硬化和ACLF先于酒精性肝硬化发生。这一发现可能很重要,不仅因为酒精是失代偿期肝硬化最常见的病因,而且可能是由于酒精本身对肠道微生物群和肠屏障功能的直接影响。
在健康个体和动物模型中,已经证明急性酗酒会增加循环中的内毒素和细菌DNA水平。
酒精显著改变肠道微生物群的组成,降低拟杆菌的相对丰度,增加变形菌。
它的代谢物,特别是乙醛,可以破坏肠道上皮细胞的紧密连接,导致肠道渗漏,促进细菌和真菌的易位,这两者都可能与肝硬化的进展有关。
在不同病因的肝硬化(例如,非酒精性脂肪性肝炎或乙型肝炎引起的肝硬化)中也观察到微生物群组成的改变以及肠道的渗漏。
4. 微生物代谢产物和转运
肝硬化发展过程中肠道微生物群的变化在很大程度上不仅归因于肠道微生物群的组成,还归因于其产物,这些产物可能具有致病作用。
一项研究表明,在肝硬化向失代偿和ACLF发展的过程中,血液代谢物特征明显改变。
针对903人的大规模研究为某些强预测代谢物模式的微生物起源带来了重要的见解,但很难对这些发现作出有力的陈述。
另一项对602名患者进行的研究表明,微生物群衍生代谢物与ACLF的存在有关。其他微生物成分,如次级胆汁酸、短链脂肪酸和色氨酸代谢物也在肝硬化中发生较大变化,与疾病的进展有关。尤其是,胆汁酸通过下调FXR受体来改变肠道屏障功能,这为临床FXR激动剂已经可用提供了新的治疗机会。
间接证据表明肠道微生物群可能会影响肝硬化的预后。
首先,研究表明,使用非选择性β受体阻滞剂(一种预防肝硬化失代偿的常用方法)治疗可改善肠道屏障,减少肠道转运时间,从而降低肝硬化患者细菌转移的可能性。
第二,在静脉曲张破裂出血期间或作为失代偿期肝硬化的长期预防措施,抗生素治疗是一种推荐的治疗方法,因为它可以改善结果,其机制可能与减少细菌移位有关。
尽管这一观点尚未得到确凿的证实,但我们知道,一种特定的微生物组表型(所谓的肠型)会导致肝功能的病理改变,并且考虑到微生物群中存在着广泛的成员,如真菌或病毒,微生物组标志物可能是非常有用的临床工具,以确定有失代偿和ACLF风险的患者。
肠道微生物群可以作为疾病进展、严重程度和治疗反应的生物标志物。
不同肝病的菌群特征
肝硬化:
瘤胃菌科、毛螺菌科与其他菌(包括肠杆菌科和拟杆菌科)进行比较,菌群比率失调。随着ACLF的发展,肝硬化菌群失调比率恶化。
该研究表明,肝硬化患者的菌群比率失调
对129名患者的研究显示,毛螺菌科减少,巴斯德氏菌科(Pasteurellaceae)的丰度可以预测死亡率。
其他研究也提供了证据,表明特定的微生物组群特征可以区分肝硬化和健康对照组,并且微生物组的变化可能能够监测病情的严重程度和进展情况。
一组45例肝移植受者中,肝移植改变了肠道菌群,使微生物多样性增加,原生细菌增加(如毛螺菌科),潜在致病菌减少(如肠杆菌科)。
该研究强调了患病的肝脏会影响肠道菌群。
肝硬化和慢加急性肝衰竭:
慢加急性肝衰竭和肝硬化的肠道微生物群特征


Trebicka Jonelet al, Nat Rev Gastroenterol Hepatol,2020
中国肝硬化患者宏基因组全关联研究(MWAS)显示,与健康个体相比,肝硬化肠道菌群组成有大于34个显著差异的菌群。肝硬化患者中韦荣球菌属和链球菌增多,而产丁酸共生菌包括Faecalibacterium prausnitzii 和Coprococcus comes减少。(下表)
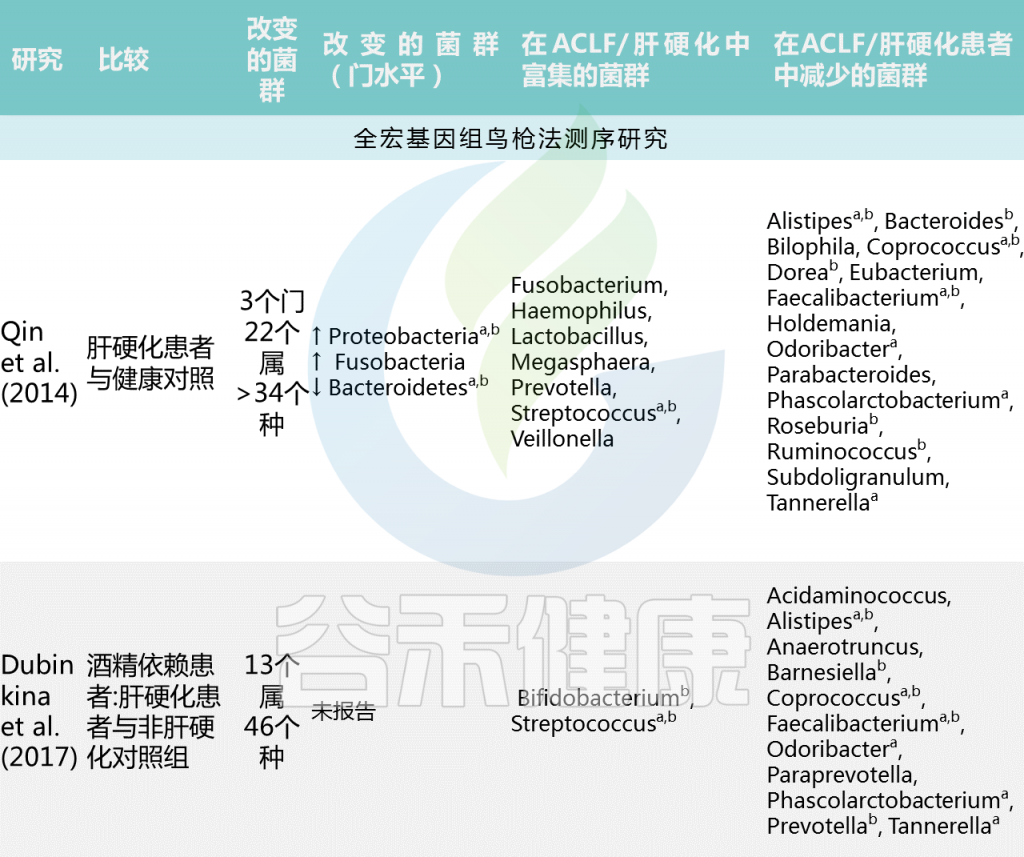

Trebicka Jonelet al, Nat Rev Gastroenterol Hepatol,2020
肝细胞癌(HCC)
持续的肝损伤和再生促进了肝细胞癌的发生,它是全球癌症死亡的第三大原因。
肝细胞癌HCC与肠道大肠杆菌过度生长有关,患者微生物群多样性增加,与产丁酸菌属(如Alistipes)减少有关,而致病性产脂多糖菌(如克雷伯氏菌)增加。
在与非酒精性脂肪性肝病相关的HCC,一项来自意大利的研究注意到拟杆菌和瘤胃球菌科增加,双歧杆菌减少。
不同部位菌群功能
肠道菌群:
从门诊病人的角度来看,肠道菌群可以证明是预测入院,MHE和晚期纤维化患者的有用工具。
一项针对NAFLD肝硬化的研究发现,粪便微生物分析中的27种细菌特征以及年龄,性别和体重指数(BMI)可预测NAFLD肝硬化。
晚期纤维化是NAFLD死亡率的重要预测指标,可以从粪便菌群中推断出纤维化,以帮助识别高危人群。
鉴于失代偿性肝硬化中存在预测的失调模式,因此将CDR(肝硬化失调率-毛螺菌科+瘤胃菌科+韦荣氏菌科/肠杆菌科+拟杆菌科)视为预测失代偿风险的工具(适用于所有常见的CLD病因)较低的分数预示结果较差,并且具有较强的预测价值。
粪便菌群也可以用来预测谁会在90天内再次入院和急性肝性脑病复发,因为急性肝性脑病和其他失代偿的粪便菌群有所不同。
唾液菌群:
唾液菌群失调比率是一种简单实用的临床工具(Lachnospiraceae + Ruminococcaceae + Veillonellaceae/ Streptococcaceae),可以预测有和无肝性脑病的失代谢性肝硬化患者90天的入院情况。
其他部位菌群:
一些研究表明,粪便微生物群和结肠黏膜相关微生物群是不同的。事实上,回肠、空肠和十二指肠粘膜样本或抽吸物的微生物群是不同的,它们可能对肝硬化具有重要的预后能力,主要表现为肝性脑病。然而,这些样本并不容易获得,而且很难将其作为生物标志物进行常规的护理点采集。
微生物工具的潜力
需要进一步的研究才能使人信服结论:患者在不同的条件和疾病下,它们的肠道微生物群既表现出疾病特异性的变化,也表现出非特异性的共同反应。因此,在其作为生物标志物的潜力得以实现之前,确定强有力的疾病特异性肠道菌群特征至关重要。
生物标志物和基于微生物的工具的潜力

Trebicka Jonelet al, Nat Rev Gastroenterol Hepatol,2020
要更准确地估计疾病对肠道微生物群的影响程度,就需要从世界各地获得强有力的大规模纵向数据,同时考虑各种混杂因素。理解和解释其他相关的混杂因素将使我们更接近以微生物组为基础的肝硬化生物标志物。
微生物组的整体观点,包括微生物基因表达和微生物衍生的代谢物或蛋白质,可能导致更准确和全面的生物标志物,类似于整合宿主遗传学与基因表达和表观遗传学时的强大疾病关联。这种包含宿主-微生物全生物的整体观点也可能支持诊断、分层和治疗的个体化,并可能开创全生物群关联研究的新时代,扩大宏基因组全关联研究领域。
我们知道,即使在没有肝病的情况下,微生物组也在调节情绪和潜在的认知中发挥作用。慢性肝病患者的脑功能改变是肠-肝-脑轴的结果,甚至在肝硬化之前就可以影响病程。
酒精使用障碍(AUD)
一项关于酒精使用障碍(AUD)患者肠-肝-脑轴的重要研究发现,抑郁、焦虑和酒精渴求与肠通透性增加相关,即使在戒酒后,肠通透性高的患者仍然存在抑郁、焦虑和酗酒。
最近发表的一篇关于AUD中这一轴的操纵的文章显示,对于AUD患者,在粪菌移植后,这些渴望、消费和长期的与AUD相关的住院治疗可能会降低。
肝性脑病
肝硬化中普遍存在的认知功能障碍是一种记忆障碍型,称为轻度或隐性肝性脑病。这可以发展为明显的肝性脑病,表现为嗜睡,迷失方向,晕眩和昏迷。
肝性脑病是肠肝轴中与微生物组相关的经典并发症,是全身性内毒素血症和炎症的结果,最终加重了神经炎症,而神经炎症是支撑肝性脑病发病机理的重要因素。
多年来,氨被认为是肝性脑病发病的关键,但后来发现全身炎症对于氨发挥其神经毒性作用是必不可少的。越来越多地描述了特定微生物群在肝性脑病和全身性炎症(包括性别影响)进程中的作用。在小鼠中,发现肠道微生物组是肝性脑病所特有的全身性炎症和神经炎症所必需的。
轻度肝性脑病和PTSD
在临床上,轻度肝性脑病需要专门的认知测试,例如心理计量学肝性脑病评分(PHES),抑制性对照测试,脑卒中诊断等。但这些测试最好在专门的环境中进行,并且需要专业知识。
轻度肝性脑病(MHE)和创伤后应激障碍(PTSD)肠道菌群之间也存在协同作用,与其他人相比,PTSD患者的菌群失调更为严重。PTSD,酒精滥用和肝硬化常常并存。微生物变化可能有助于将MHE与PTSD区别开来,并且可以帮助我们定义较新的疗法。
老年肠道-肝-脑轴改变
利用肠脑轴的主要挑战是老年人群(慢性肝病患者)。老年人患者的致病菌往往更具致病性,随着合并症,住院和抗生素暴露于更大的革兰氏阴性病原体中,病情恶化。
与类似的老年非肝硬化对照组相比,年龄较大(> 65岁)的肝硬化患者往往会改变肠道-肝-脑轴,从而增加炎症并损害记忆的认知能力。在相关网络分析的研究中,更重要的是,在老年肝硬化和非肝硬化组中也发现了类似的模式,这表明老年患者已经具有潜在的肠脑轴改变。
这对肝移植具有重要意义,因为尽管进行了肝移植,但与年龄相关的肠脑轴变化仍可能持续。人口老龄化往往会导致多种其他合并症和精神疾病,需要额外的照顾以确保最佳的移植后效果。
针对肝硬化肠道微生物群的策略
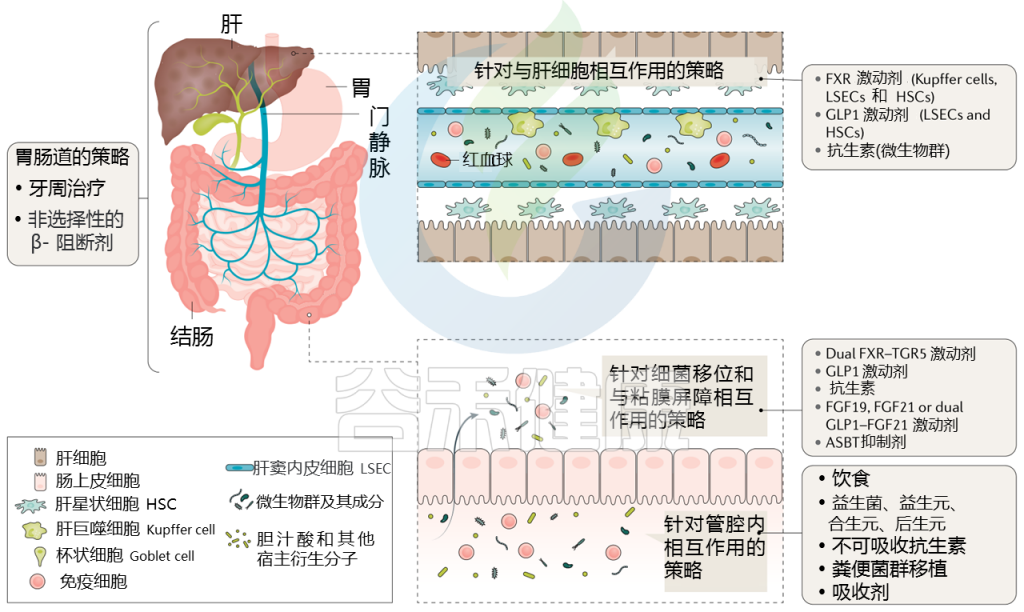

Trebicka Jonelet al, Nat Rev Gastroenterol Hepatol,2020
通过治疗(部分成立和部分试验)调节肠道微生物群可能改善肝硬化患者的预后,而微生物组生物标志物可能反映对这些治疗的反应。下表显示了不同的策略。
针对肠道微生物群的肝硬化干预措施

Trebicka Jonelet al, Nat Rev Gastroenterol Hepatol,2020
接下来主要介绍一些具体疗法:饮食、益生菌、抗生素、白蛋白、牙周、粪菌移植等,有些目前正在临床试验中。
饮 食
除其他因素外,饮食对肠道微生物群有重要影响,这一点已在各种疾病和条件下得到研究。一项针对国际肝硬化人群的研究报告称,咖啡、茶、蔬菜和酸奶的摄入都与微生物群落多样性的增加和住院率的降低有关,这为调节肠道微生物群落以促进肝脏健康提供了前景广阔的途径。(具体的饮食方面建议见文末附录部分)
益 生 菌
益生菌已经成为肝硬化临床研究的主题,特别是它们对脑功能的影响和肝性脑病的风险。
益生菌在肝硬化前可以改善NAFLD的组织学,在肝硬化中可以减少失调,对逆转OHE的二级预防有效,减少失代偿事件的住院率,但可能或不能改善认知能力(MHE)。
大多数制剂都含有乳酸菌,可产生短链脂肪酸,有利于结肠健康。一项短期(5天)口服两歧双歧杆菌和植物乳杆菌8PA3的小型试点研究表明,与单独标准治疗相比,口服两歧双歧杆菌和植物乳杆菌8PA3能恢复肠道菌群,并能更好地改善酒精性肝损伤。
以上是关于益生菌的前沿研究,如果想要尝试的话,最好是经过肠道菌群检测或者在医生的指导下服用更可靠。
抗 生 素
抗生素,也被用作失代偿期肝硬化的预防性治疗。
腹水患者发生自发性细菌性腹膜炎(SBP)的风险增加,接受抗生素预防来预防自发性细菌性腹膜炎的发展。这种SBP的一级预防已经被证明可以提高某些患者的生存率。
在一项多中心、随机对照试验中,对严重肝硬化患者使用抗生素,总体生存率没有任何益处,但是,同样,腹水中白蛋白浓度低的患者在接受抗生素治疗时,生存率得到了提高。
预防性抗生素治疗也同样有效建议用于其他临床情况,如静脉曲张破裂出血、既往SBP和复发性显性肝性脑病。然而,尽管对最脆弱的患者的短期效果可能是有益的,但在其他许多患者中,这些治疗不能阻止进一步的失代偿和ACLF。也许这种情况的部分原因是抗生素引起的微生物群落多样性减少。
因此,需要开发一种更具针对性的肠道微生物群来改善肝脏健康。
牙周卫生
也许你不知道,牙周卫生也会与肝硬化有关。已发表的人类MWAS表明,在肝硬化期间,肠道微生物组向口腔微生物组转变。
减少胃部充当天然屏障的酸性环境可能促成口腔微生物群转移到肠道,在肠道上皮屏障适应不同的微生物组分。如前所述,这些变化可能促进肠道微生物群通过肠道屏障的转移,进而导致并发症。因此,防止口腔微生物不必要的转移到肠道,可以成为未来肝硬化治疗的基础。
一项研究表明,与20名非肝硬化对照组相比,30名肝硬化患者的牙周卫生干预措施改变了肠道微生物群,改善了肝性脑病。
白蛋白作为潜在疗法
当肠道屏障被削弱时,它会双向泄漏,这也会导致重要的宿主分子从血液室泄漏到肠腔。例如,在酒精性肝病动物模型中,肠道通透性增加导致粪便白蛋白浓度升高。
随着肝硬化的进展,肝脏中的白蛋白合成受到损害:一方面,肝损伤导致肝细胞功能质量的丧失;
另一方面,肝脏僵硬程度的增加,白蛋白合成减少。不仅白蛋白减少,而且它能结合毒素和其他有害物质。这些观察结果将白蛋白水平定位为肝病严重程度的关键参数。
白蛋白还可通过前列腺素E2改善免疫B细胞功能,减少肝硬化患者的免疫反应,从而阻止与ACLF和死亡密切相关的全身炎症爆发。
长期白蛋白治疗肝损害和非自发性细菌性腹膜炎感染患者分别通过提高生存率和解决ACLF改善了临床结果。因此,白蛋白泄漏到肠腔可能会影响肠道微生物群并被其代谢,从而可能改变微生物组分,间接改变宿主。
粪菌移植(FMT)
失代偿期肝硬化中菌群移植的初步研究正在出现,其安全性也很高,为更大规模的研究铺平了道路。
菌群移植后变形菌减少,放线杆菌增多,肝脏疾病严重程度改善,严重酒精性肝炎患者3个月时的死亡率降低,1年生存率提高。
慢性乙型肝炎FMT组出现微生物改变,HBeAg降低。FMT减少了短期酒精渴求和消费,以及AUD相关的住院治疗。
一项包括20名复发性肝性脑病患者在内的开放性随机试验观察到,使用FMT灌肠剂减少了住院人数,改善了认知和菌群失调。
另一项研究也证明了这一点。使用口服胶囊制剂的FMT的新应用也显示出类似的安全性和对肝硬化和肝性脑病的治疗效果。然而,由于报告了一些由耐yao菌转移导致的FMT相关死亡病例,包括一些肝硬化患者,因此需要谨慎。因此,更好地理解和全面描述这些研究可能带来明确的微生物调节干预治疗失代偿期肝硬化。
加深菌群影响机制的理解
未来的研究应该探索和加深对肠道微生物组变化影响肝硬化进展和失代偿发展的动力学和机制的理解。
诊断、治疗
由于肝硬化表现为肠道微生物群与宿主的大量重要相互作用,微生物组诊断和治疗几乎是治疗疾病进展和失代偿发展的必经之路。需要进一步倡导基于微生物组检测和靶向疗法。
改善预后
针对不同水平微生物群与宿主相互作用的策略改善患者的预后。
产学研合作
使之更容易获得和更具成本效益,微生物检测需要更广泛地适应,因为它相对容易收集且信息量较大。可以帮助诊断,预测和潜在地个性化治疗。
多学科综合
多学科例如微生物学,宿主遗传学,基因组学,表观遗传学,代谢组学,营养学等相结合,更好地利用微生物帮助我们有效的进行慢病管理,预防为主,将指导落实到饮食、生活方式等具体应用上,用全新的视角帮助我们认识健康,保持健康。
饮 食 方 式
尽可能选择对肝脏友好的食物,帮你保护肝脏。以下是一些饮食技巧:
选择全麦谷物,面包和谷物。
多吃各种颜色的水果和蔬菜。
选择低脂乳制品。
选择瘦肉蛋白质。
选择植物油,例如橄榄油,而不是黄油。
零食坚果和种子食物。
减少钠摄入量。
每天喝8-10杯水。
避免含有反式脂肪的加工食品。
避免添加糖。
选择可以长期生活并遵循的饮食计划。
吃八分饱腹感
少吃多餐,每三到四个小时吃一顿小餐或吃零食,以补充精力。
限 制 饮 酒 量
肝脏是负责代谢营养物质和其他摄入物质的主要器官。如果体内酒精过多,肝酶可能没有足够的能力对其进行处理。过量的酒精会在身体的其余部分循环,产生负面影响。
定期减少饮酒量很重要。医生甚至可能建议你完全戒酒。
增 加 运 动 量
运动的好处不仅限于减肥和控制体重,还可以帮助减少肝脏周围的脂肪。定期运动也可以改善你的情绪,助你维持在最佳的健康状态。
开始时要循序渐进,比如可以每周进行150分钟运动开始,尽量选择你喜欢的运动,例如,跑步和步行的组合,多人参与的活动增加趣味性(如各种球类运动),健身房的器械训练等。
睡 觉
充足的睡眠对于保持健康状态非常重要(包括慢性肝病治疗期间睡眠很重要)。如果你觉得入睡困难,开始慢慢尝试练习良好的睡眠习惯,例如:
每天都在同一时间上床睡觉和起床,作息规律。
避免咖啡因,烟草和其他刺激物。
尽量保持卧室环境清爽舒适。
运动尽量安排在清晨或午后,不要在睡前剧烈运动。
相关阅读:
膳食胆固醇通过调节肠道微生物群和代谢产物驱动脂肪肝相关的肝癌
参考文献:
Grat M, Wronka KM, Krasnodebski M, et al. Profile of Gut Microbiota Associated With the Presence of Hepatocellular Cancer in Patients With Liver Cirrhosis. Transplant Proc 2016;48:1687-91.
Jones R M, Neish A S. Gut Microbiota in Intestinal and Liver Disease[J]. Annual Review of Pathology:Mechanisms of Disease, 2020, 16.
Acharya Chathur,Bajaj Jasmohan S,Chronic Liver Diseases and the Microbiome: Translating Our Knowledge of Gut Microbiota to Management of Chronic Liver Disease.[J] .Gastroenterology, 2020
Bajaj JS, Sharma A, Dudeja PK. Targeting Gut Microbiome Interactions in Service-related Gastrointestinal and Liver Diseases of Veterans: Meeting Summary.Gastroenterology 2019.
Liu R, Kang JD, Sartor RB, et al. Neuroinflammation in Murine Cirrhosis Is Dependent on the Gut Microbiome and Is Attenuated by Fecal Transplant. Hepatology 2020;71:611-626
Ren Z, Li A, Jiang J, et al. Gut microbiome analysis as a tool towards targeted non-invasive biomarkers for early hepatocellular carcinoma. Gut 2019;68:1014-1023.
Trebicka Jonel,Bork Peer,Krag Aleksander et al. Utilizing the gut microbiome in decompensated cirrhosis and acute-on-chronic liver failure.[J] .Nat Rev Gastroenterol Hepatol, 2020
Lang S, Fairfied B, Gao B, et al. Changes in the fecal bacterial microbiota associated with disease severity in alcoholic hepatitis patients. Gut Microbes 2020;12:1785251.
Ponziani FR, Bhoori S, Castelli C, et al. Hepatocellular Carcinoma Is Associated With Gut Microbiota Profile and Inflammation in Nonalcoholic Fatty Liver Disease. Hepatology 2019;69:107-120.
Gao B, Duan Y, Lang S, et al. Functional Microbiomics Reveals Alterations of the Gut Microbiome and Host Co-Metabolism in Patients With Alcoholic Hepatitis. Hepatol Commun 2020;4:1168-1182.
Duan, Y . et al. Bacteriophage targeting of gut bacterium attenuates alcoholic liver disease. Nature 575, 505–51 1 (2019)
谷禾健康
随着研究微生物组的需求越来越旺盛,用于微生物组数据分析的平台也越来越多。分享两款最新的微生物组数据分析平台。
能够处理来自NGS平台的数据,包括Illumina MiSeq,Illumina HiSeq或Illumina NovaSeq,还可以处理以前的454焦磷酸测序技术。CoMA着重于短读而非长读的数据处理。
处理过程包括数据预处理、质量检查、对操作分类单元(OTUs)的聚类、分类、数据后续处理、数据可视化和统计评估。输出结果包含可供发表的图形和标准化格式的文件(例如,制表符分隔的OTU表、biom、Newick tree)。
比起qiime,它的下载安装没那么复杂,兼容linux、mac、windows系统,并且交互式的用户界面更方便小白上手。
性能评估方面,使用了模拟数据和真实的土壤数据与现流行的Mothur、QIIME和QIIME2软件进行了比较,其结果是一致的。
Coma的交互界面
$ coma
如同回答问题般的操作
“Do you want to start a new project?” → Yes
“Do you want to assign your files and choose the number of CPUs?” → Yes
“Are you using paired-end reads?” → Yes
工作流程
使用了各种开源的第三方工具,并以Bash脚本的形式将它们组合到了线性分析工作流程中,从原始输入文件(以FASTQ格式)开始。
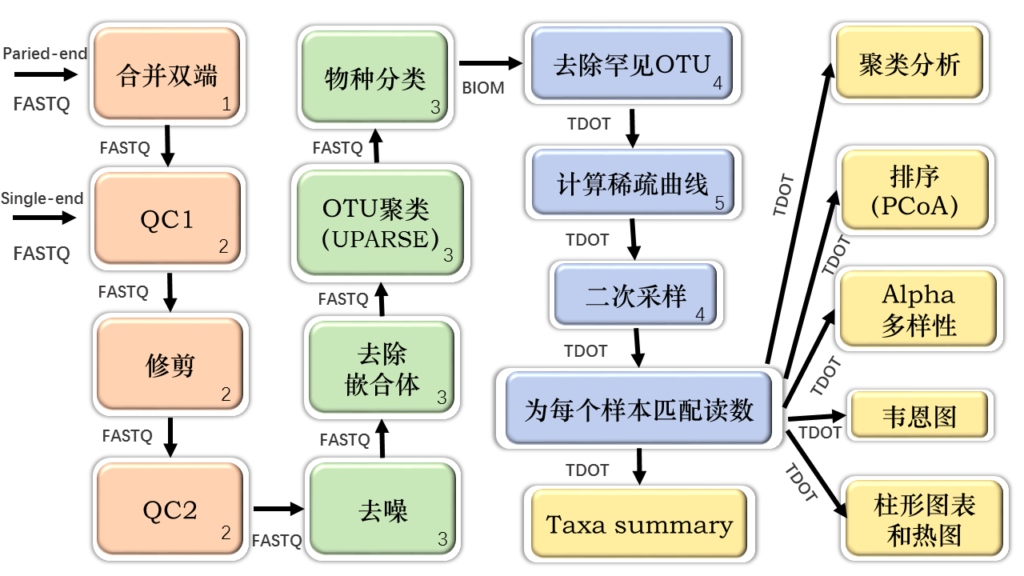
不同的颜色代表CoMA工作流程的四个子部分:数据预处理和质量检查(橙色),OTU的聚类和物种分类(绿色),数据后处理(蓝色)以及数据可视化和统计分析(黄色)。
箭头指示发生的顺序,并指定了每个步骤的输入所需的文件类型。通过BLAST、LAMBDA或RDP使用任一可用数据库完成物种分类(例如Sliva或用户自定义数据库)。
数字表示用于特定CoMA步骤的第三方工具:1 = PANDAseq,2 = PRINSEQ,3 = LotuS / sdm,4 = QIIME,5 = Mothur。TDOT =制表符分隔的OTU表
模拟社区群落数据测试
来自公共模拟微生物库。选择了3个数据集分别为MOCK-13、MOCK-16、MOCK-26.。Mock-13包含21个细菌菌株(18个属;三个重复),MOCK-16包括古细菌,共59个菌株(46属;三个重复),Mock-26包含来自11个真菌菌株(11属)的ITS数据。
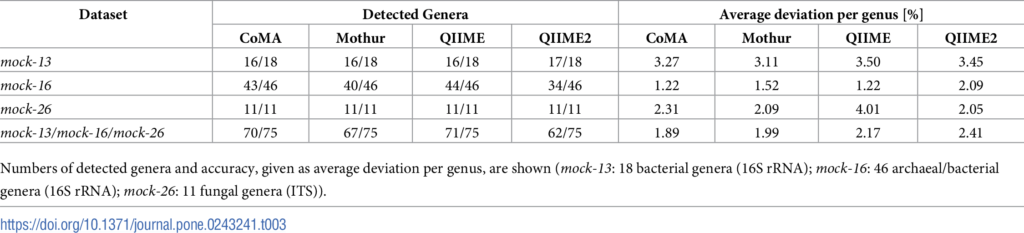
表1展示了四个不同分析平台的基准测试结果:使用CoMA进行的数据分析揭示了18个属中的16个,与Mothur和QIIME数量一致。

综合所有模拟社区,CoMA表现出总体最高的准确性,每个分类单元的平均偏差为1.89%(表1),而Mothur(1.99%),QIIME(2.17%)和QIIME2(2.41%)。从层次聚类分析来看,CoMA和QIIME2显示最小的余弦距离(0.08)。Mothur的距离为0.11,QIIME的距离为0.12。所有四个管线之间的相互关系比估计值更紧密(总余弦距离:0.18)。
土壤微生物数据集测试
(森林F、草地GR、沼泽S)

Shannon-Wiener多样性。字母表示不同栖息地使用的分析工具之间的显著差异。三种分析工具都显示出了不同水平的Shannon-Wiener多样性,Mothur>QIIME>CoMA


主成分分析结果表示CoMA和QIIME分析得到的微生物群落具有更高的相似性。
韦恩图也显示出相似的结论,无论是哪个分类水平,Mothur分类得到的OTU数目都是最少的,CoMA和QIIME分类得到的一样的OTU的数目最高。

分析了三个土壤类型的关键科水平物种(即丰度>5%的序列)。在表格中被列举出来的关键科水平物种中,无论土壤类型为哪个,三种分析工具都确定了6个关键科。
另一方面,Mothur确定了在CoMA和QIIME中都找不到的4个关键科,CoMA和QIIME再次确定了相同的分类单元(18个中的14个)

在森林、草原和土壤中的每个分类水平的未分类读数。CoMA和QIIME在所有分类水平上的表现都相似。但是,在分析草地时,CoMA会发现更多的Order和Family(分别为5%和+ 3%; p <0.001)。
总体而言,CoMA的表现与Mothur和QIIME相当,在微生物多样性分析和分类学分类上导致的差异,文章中也做出了相应解释,这通常与算法、参考数据库、参数选择有关。
CoMA的主要优点是图形用户界面支持的直观且用户友好的操作。它使入门级用户无需进行费时费力的培训即可执行扩增子测序数据分析并获得可靠的结果。CoMA还提供了随时停止未完成的分析以稍后再继续或重新计算部分工作流的可能性。通过简单地调整决定性的输入参数,而无需进行完整的重新计算。未来,CoMA将会支持ASV结果输出。
一款用于研究微生物群与宿主表型数据关系的分析软件,可以本地安装也可在线使用。可以是16SrRNA测序数据,也可以是宏基因组数据。
软件自建了一个微生物组合表型数据的综合数据库,用户能够共享或存储自己的数据,通过数据库和相关分析脚本该可以清楚地展示微生物组和表型数据(例如生活方式等)之间的相关性。依旧是交互式界面,但这个更直观,可以本地安装也可以在线使用。
用户界面

详细手册:https://mizuguchilab.org/manta/manta-tutorial.html
操作类似于MicrobiomeAnalyst这种平台,非常好上手。具体怎么操作就不赘述了,主要看看这款软件在表型数据的分析中表现怎么样。
案例研究
脂肪摄入量与微生物群之间的相关性
数据来源于NIBIOHN队列数据,20名健康成年志愿者(21-41岁,男性)
首先,使用基于OTU的Bray-Curtis距离进行PCoA分析,以评估志愿者之间的相似性。可以通过勾选感兴趣的表型数据进行分组分析,比如图中的abc分别勾选了“食用油”、“脂肪”和“ω-6多不饱和脂肪酸”,然后计算Spearman相关系数,搜索与“脂肪”相关的肠道菌群。


这项分析表明,Lachnospiraceae与脂肪摄入量呈正相关。Lachnospiraceae由丁酸产生菌组成,据研究,低碳水化合物摄入量的高脂肪饮食与Lachnospiraceae和Ruminococcacea的丰度比有关。根据这些结果,推测Lachnospiraceae和Ruminococcacea的比例主要受饮食,特别是脂肪摄入量的影响。
然后反过来,探讨与Lachnospiraceae菌相关的其它表型数据。观察到:
(a)与单不饱和脂肪酸或饱和脂肪酸摄入量呈正相关;
(b)与体育活动时间相关的参数呈负相关;
与Ruminococcaceae菌相关的其它表型数据,观察到:
(c)与ω-6多不饱和脂肪酸摄入量呈正相关;
(d)与体重、总能量消耗、碳水化合物摄入(如煮熟的大米和谷物)等身体成分相关的参数呈负相关。
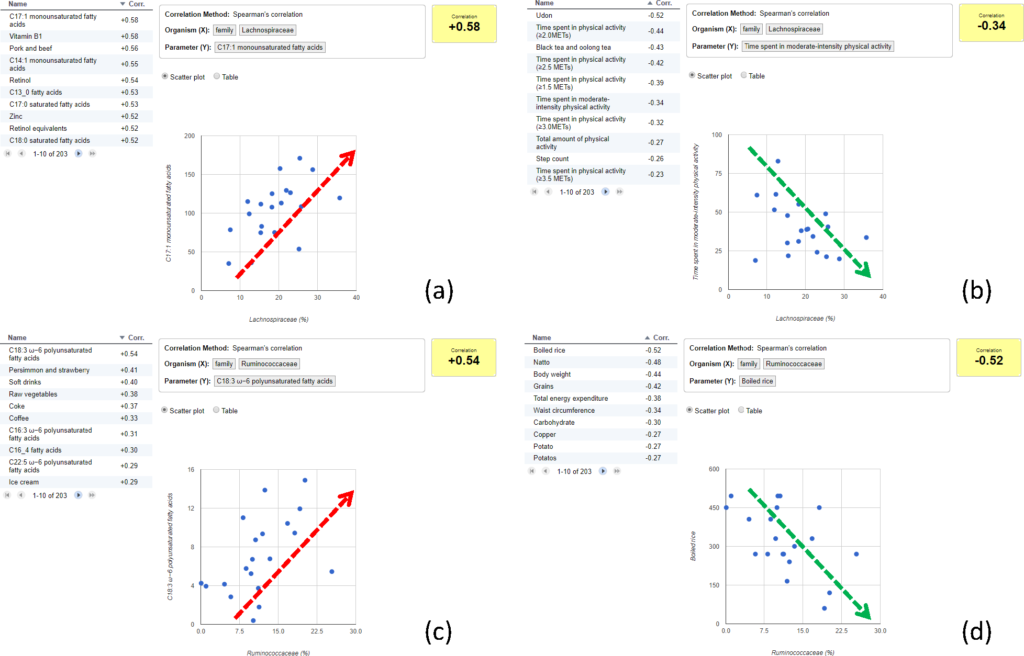
使用MANTA分析出的结果与案例中的结果一致,不同类型的脂肪酸会分别影响Lachnospiraceae和Ruminococcacea。这表示MANTA成功的假设了微生物群和生活方式之间可能存在的联系。
基于以上,我认为MANTA是一个不错的辅助分析工具, MANTA还提供了一个更加简易的版本MANTA BASIC, 输入的文件也比较简单,OUT表和含表型数据的制表符格式文件。
延 伸
软件开发拥有一套复杂的流程。从提出问题,编写方法、开发,然后是一系列的测试,通常包含基准测试、模拟数据的测试、真实的生物数据的测试。每一步都需要经过严格的控制,包括要使用哪些数据,要应用哪种性能指标以及要采用哪种基准。最后整理成文档,将方法放置在软件包或插件中,开放存取,持续的测试更新和版本控制等。复杂但也有逻辑可寻。如果你在开发方面感兴趣,可以阅读这篇文章“Measuring the microbiome: Best practices for developing and benchmarking microbiomics methods”,它详细的介绍了开发用于微生物研究的软件或方法的一套流程,可以说是一篇指南。
工具的选择应该基于解决什么样的问题。对于CoMA,如果你觉得用qiime太困难了,可以试试CoMA,这种非命令式的交互界面还是很好理解的,从案例研究的结果来看,与qiime的结果几乎一致。虽然是新开发的软件,但较为内核的算法借鉴或直接使用了老牌的工具。
MANTA是个研究表型数据与微生物相关性的方便好用的软件,比起自己写代码和调试,这个可以直接上手,很方便。
相关阅读:

谷禾健康
感觉疲惫?沮丧?容易出汗?经常感冒?免疫力下降?骨痛?脱发?…
这些都可能与维生素D缺乏有关。
维生素D对人体健康很重要。然而流行病学研究表明,全世界近十亿人口受维生素D缺乏症的影响。
一项针对北京和上海50-70岁人群调查显示:

注:VD水平缺乏(低于20ng/ml);VD水平不足(低于30ng/ml)
首先我们来看下维生素D在人体中扮演怎样的角色。
维生素D
· 维生素D是脂溶性维生素
· 参与钙和磷的吸收,骨骼健康以及肌肉的生长和发育(生理过程)
· 刺激肌肉细胞的增殖和分化(影响肌肉系统)
· 其充足的供应可预防呼吸道感染,间接参与具有抗生素特性的化合物的生产(免疫系统)
· 其最佳含量对皮肤的状况和健康以及男性和女性生殖过程的调节都有积极的作用(皮肤)
为什么会有这么多人出现维生素D缺乏?
1. 饮食摄入和/或吸收减少
某些吸收不良综合症,如腹腔疾病,短肠综合症,胃搭桥,炎性肠病,慢性胰腺功能不全和囊性纤维化,可能导致维生素D缺乏。老年人中口服维生素D摄入量较低的情况更为普遍。
2. 减少阳光照射
大约50%-90%的维生素D通过阳光通过皮肤吸收,其余的来自饮食。为了防止维生素D缺乏,每天需要20分钟的阳光照射,皮肤暴露在40%以上。维生素D的皮肤合成随着年龄的增长而下降。黑皮肤的人皮肤维生素D的合成较少。在住院或长期住院的患者中减少的日光照射也会导致维生素D缺乏。持续使用防晒霜的人的有效日晒量降低。
3.内源性合成减少
患有慢性肝病(例如肝硬化)的个体可能具有缺陷的25-羟化作用,导致活性维生素D缺乏。1-α25-羟化作用的缺陷可见于甲状旁腺功能亢进,肾衰竭和1-α羟化酶缺乏症。
4.肝分解代谢增加
某些yao物会诱导肝p450酶,从而激活维生素D的降解。
5.维生素D 抵抗
在遗传性抗维生素D佝偻病中可以看到维生素D受体抵抗。
维生素D缺乏症的大多数患者无症状。
轻度的慢性维生素D缺乏症,也可能会导致慢性低钙血症和甲状旁腺功能亢进,特别是在老年人群中,可能导致骨质疏松,跌倒和骨折的风险。
长期严重维生素D缺乏症患者会出现与继发性甲状旁腺功能亢进有关的症状,包括骨痛,关节痛,肌痛,疲劳,肌肉抽搐(束缚)和无力。脆性骨折可能是由于长期缺乏维生素D导致骨质疏松所致。
在儿童中可能产生的症状:烦躁,嗜睡,发育迟缓,骨骼变化或骨折等。
其他一些症状可能会被忽视,其实也与维生素D缺乏有关,例如:
感到疲惫,免疫力下降,反复感染,容易出汗,脱发,伤口愈合慢,头晕,心脏问题,超重或肥胖,情绪低落,认知功能障碍等。
25(OH)D是血中维生素D的主要存在形式, 而且血清或血浆中25(OH)D受机体调节的影响较小, 可以在较长时间内维持稳定,可以体现膳食摄入维生素D和阳光维生素D的总量,这是评估维生素D水平是否正常的首选临床措施。
临床上,血清25(OH)D水平标准如下:
足够: 大于20–30 ng/ml 或50–75 nmol/L
安全上限: 不超过60 ng / ml 或 150 nmol/L
有毒: 高于150 ng/mL 或 375 nmol/L
毒 副 作 用
高维生素D水平是由于过量摄入而不是由于过度日光照射引起的。
急性中毒可导致急性高钙血症,可引起混乱,厌食,呕吐,多尿,多饮和肌肉无力。
慢性中毒可导致肾钙化和骨痛。
维生素D缺乏与许多非传染性疾病的发生有关,如结肠癌,乳腺癌,卵巢癌,肾癌,胰腺癌,精神分裂症,阿尔茨海默,多发性硬化,骨软化症,佝偻病,骨质疏松,糖尿病等。
4.1 呼吸道疾病
一些研究表明,维生素D补充剂可以增强免疫反应并预防呼吸道感染。
一项来自14个国家/地区的11,321人的研究表明,补充维生素D可以使维生素D缺乏和适当水平的人降低急性呼吸道感染(ARI)的风险。
来自三项随机对照试验的Meta分析研究中,发现补充维生素D可以降低维生素D水平低于25 nmol / L的患者慢性阻塞性肺疾病恶化率。
4.2 新 冠 肺 炎
与呼吸道疾病相关的,例如COVID-19。
最近的一项研究表明,血液中25(OH)D的浓度至少为30 ng / mL有助于住院的COVID-19患者减少临床预后不良和死亡的可能性。
研究分析了235例COVID-19患者的住院资料。与维生素D缺乏症患者相比,在40岁以上的患者中,维生素D含量适当的患者发生不良后果(包括失去知觉,缺氧和死亡)的可能性降低51.5%。
在疫情期间,我们可能常听到这样的词:“细胞因子风暴”。而维生素D缺乏会增强细胞因子风暴的过程。
注:细胞因子是蛋白质,是免疫系统不可或缺的一部分。它们可以同时具有促炎和抗炎作用,并发挥重要作用,有助于预防感染和疾病。但是,在某些情况下,细胞因子也会引起组织损伤。
细胞因子风暴是指响应感染或其他因素而发生的促炎性细胞因子的失控释放。细胞因子的这种失调和过度释放会导致严重的组织损伤,并增强疾病的进展和严重程度。
细胞因子风暴是导致多器官功能衰竭和急性呼吸窘迫综合征(ARDS)的主要原因,也是导致COVID-19进程和严重程度的重要因素。例如,已显示严重病例COVID-19的患者释放大量细胞因子,尤其是IL-1和IL-6。
维生素D缺乏与免疫功能降低有关,并可能增强细胞因子风暴。当然该领域还在研究中。
需要特别注意,单独服用补充维生素D并不能让你免受COVID-19的侵害。不过,缺乏维生素D可能会损害免疫功能,从而增加感染风险和对疾病的敏感性。
4.3 自 闭 症
自闭症是遗传和环境因素共同作用导致的神经发育异常,自闭症人群中维生素D的缺乏较常见,而维生素D能够促进儿童的大脑神经发育,因此可能在自闭症的病因学上起着重要作用。
采用随机双盲对照临床试验(RCT)设计,纳入109名自闭症儿童(85名男孩和24名女孩;年龄为3-10岁)。将自闭症患儿随机分组,分别接受维生素D3或安慰剂治疗4个月。结果发现,自闭症儿童可很好地耐受高剂量的维生素D3,且临床疗效较好。
这项研究是第一个证明维生素D3补充剂有益于自闭症患者的随机双盲对照试验,但是仍需更大规模的随机对照试验来严格验证维生素D对自闭症患者的疗效。
4.4 癌 症
对包括57,000多名受试者的18项随机对照试验(RCT)进行的荟萃分析发现,每天摄入维生素D补充剂可降低总死亡率。补充钙和维生素D降低了罹患总癌症,乳腺癌和结直肠癌的风险。
一项RCT显示,钙和维生素D可以大大降低绝经后妇女的所有癌症风险。
近几年来的人群研究还发现,机体维生素D水平与肠道菌群之间也存在密切联系。
生命早期生长发育与维生素D有关
我们知道,婴儿在出生6 个月内身体生长发育迅速,所以需要充足的营养。维生素D 缺乏是婴儿早期营养不足的主要原因之一,维生素D 补充不足会严重影响婴儿的骨骼发育。
有研究显示,婴儿体重与维生素D 缺乏具有明显关联性。婴幼儿缺乏维生素D 也会影响肠道菌群的定植能力和数量平衡,导致消化系统功能失调造成吸收功能障碍。

婴儿期肠道菌群数量的改变受多种因素影响。研究显示,维生素D可通过信号传导来增强肠道结构屏障的完整性和提高肠道免疫反应能力,进而稳定菌群在肠道的定植和数量平衡。
婴儿在1 岁以内是肠道内菌群定植、演替和达到平衡的主要阶段,特别是肠道内双歧杆菌、大肠杆菌、乳杆菌等主要菌体在婴儿出生半年内完成定植、演替和数量的平衡,对增强机体免疫力和促进消化吸收起到了重要作用。
在最近对3-6 个月大的婴儿肠道微生物群的研究中,研究人员发现脐带血维生素D 水平与乳球菌减少有关。

正常成年人补充维生素D 后,普氏菌属(Prevotella)的丰度随维生素D 的摄入量增加而增加,嗜血杆菌(Haemophilus)和韦荣氏球菌(Veillonella)的丰度随维生素D 的摄入量增加而降低,粪球菌属(Coprococcus)和双歧杆菌属(Bifdobacterium)的丰度与血清25-OH-D 水平呈负相关。
上述资料表明从婴儿期到成人期,维生素D 对肠道菌群的组成具有调节作用。
维生素D减少与肥胖有关
有研究显示,饮食中类固醇类物质的缺乏会影响肠道菌群的定植和平衡状态,使菌群数量明显增加,可能与肥胖的发生均有一定关系,但仍需进一步研究证实。
维生素D 还可促进脂肪细胞内Ca2 + 的吸收,使脂肪酸合成酶的活性增强进而减少脂肪的分解,增强脂肪细胞的储脂能力。所以体内维生素D 含量减少时,体内脂质代谢会明显增强和细胞内储脂能力降低,导致肥胖发生。
国内学者发现,超重和肥胖患者血清中维生素D 水平明显低于正常水平,且维生素D 与BMI 和腰围指标呈独立负相关性。
维生素D 缺乏的婴儿体重和BMI 指标明显高于维生素补充充足的婴儿。肠道菌群数量的增加与肥胖具有密切联系,特别是乳酸杆菌和双歧杆菌以及肠球菌能增加体重和引起肥胖。
该研究发现,观察组婴儿补充维生素D 后乳酸杆菌、双歧杆菌、肠球菌含量明显低于对照组。
活性维生素D代谢产物
多项研究表明,肠道菌群会改变肠道维生素D的代谢,而益生菌补充剂会影响循环中的维生素D水平。维生素D缺乏会降低防御素的生成,防御素是一种对于维持健康肠道菌群非常重要的抗菌分子。正如研究人员期待的那样,口服合成防御素分子能够恢复肠道菌群平衡,降低血糖水平,并改善脂肪肝症状。

最近Nature Communications上发表的一项横断面研究,对567名老年男性的血清维生素D代谢产物及粪便菌群进行深入分析后发现,8种特定肠道细菌分类群与维生素D活性形式——1,25(OH)2D的水平相关,而更高的1,25(OH)2D水平与更健康的肠道菌群(更多的丁酸盐产生菌)相关。同时,1,25(OH)2D水平较高的男性具有更大的多样性。
这些结果提供了强有力的证据,表明宿主维生素D信号和老年人肠道菌群健康之间存在重要的相互作用。肠道中CYP27B1-和CYP24A1-24-羟化酶的共定位表达可能会被存在的微生物群增强或抑制。
无论是通过饮食干预和/或在临床上给合适的人群补充维生素D,发现与更有利的肠道微生物多样性相关的活性维生素D代谢产物(包括已知的丁酸盐生产者的特定微生物)可能会为临床提供潜在的干预目标。
自闭症儿童维生素D水平较低现象
大量研究证实,自闭症儿童的维生素D 水平低于同龄健康人群,并且足量、规范地补充维生素D 可改善其自闭症核心症。
动物实验研究表明,发育早期维生素D 缺乏会导致永久性的大脑发育异常,如神经生长因子(NGF)表达减少、侧脑室增大、神经结构或神经递质相关基因(如MAP2, NF-L,GABA-Aa4)表达降低、脑组织氨酰胺水平普遍降低、去甲肾上腺素和丝氨酸水平升高、多巴胺水平升高、5-HT 水平下降等。
维生素D——神经保护剂
而维生素 D的存在,可以改善由于脑部炎症和神经毒性引起的脑损伤,促进NGF表达,延缓神经元细胞死亡。因此,维生素D 可以作为一种神经保护剂保护大脑皮层神经元避免兴奋性毒性。
自闭症儿童肠道患病率更高
自闭症儿童存在免疫系统功能紊乱,胃肠道紊乱患病率高于正常发育儿童。一项超过14,000 例自闭症患者的多中心研究显示,自闭症患者炎症性肠道疾病患病率为0.83%,而全院患者患病率为0.54%;自闭症患者其他胃肠道紊乱疾病患病率为11.74%,而全院患者患病率为4.5%,表明自闭症肠道患病率更高。
Wang L 等研究发现,在自闭症患者和正常人群中发现鲍氏梭状芽孢杆菌存在明显差异。自闭症患者肠道中的脱硫菌属比正常人群明显增多,且普通拟杆菌属含量丰富,自闭症患者与对照组正常人群肠道中的放线菌及变形菌属含量也有差异。
还有研究观察到自闭症患者肠道中有更高水平的萨特菌和瘤胃球菌属。国外通过自闭症小鼠模型肠道菌群宏基因组的研究发现,其脆弱拟杆菌属、梭菌属比例较对照组明显异常,口服脆弱拟杆菌可改善自闭症样行为症状。
维生素D通过影响Th17/Treg细胞平衡改善菌群紊乱
维生素D 可调节Th 细胞以及Treg 细胞功能,可抑制Th17 细胞,并促进Treg 细胞的增殖。Th17/Treg细胞平衡可能是维持正常的机体整体免疫功能和肠道免疫屏障功能中关键的因素。
维生素D 通过影响Th17/Treg细胞平衡调节自闭症肠道的菌群紊乱,进而通过脑肠轴作用改善神经系统的发育。
这些研究提示,自闭症存在肠道微生物生态失衡,肠道微生物失衡的改善很有可能作为自闭症治疗新的发展方向。
VD与肠道菌群相互影响,从而影响肠道疾病
结直肠癌
2009 年Volker Mai 团队研究了非裔美国人结直肠癌高于白种美国人,研究发现非裔美国人饮食中的环杂胺明显增高,维生素D 摄入量明显减少,两组人群的粪便微生物群组成也存在差异,推测环杂胺、维生素D 与肠道菌群的组成有相互影响,从而影响肠道环境促进结直肠癌的发生。
克罗恩病
维生素D 缺乏与克罗恩病(Crohn’s disease,CD)的发病有关,研究表明补充维生素D3 会改变CD 患者肠道细菌组成,参与者口服维生素D3,从第1 天至第3天每天服用20000 IU,然后每隔一天服用一次,共4 周,CD 患者口服一周后,Alistipes 菌、Barnesiella 菌、紫单胞菌科(Porphyromonadaceae)、Roseburia 菌、Anaerotruncus 菌、Subdoligranulum 菌和Ruminococaceae(均为厚壁菌门)的丰度显著增加。
炎症性肠病
1,25-OH2-D3 治疗还可在炎症性肠病中改变肠道菌群的组成,从而对小鼠有保护作用。
在肠道性疾病中肠道细菌组成多发生改变,补充维生素D 则可调节已经发生改变的肠道菌群组成。通过补充维生素D可以改变肠道内不同菌种的含量,调节肠道菌群的组成,增加双歧杆菌,拟杆菌,降低大肠杆菌和肠球菌的含量和比例。
肠炎
此外,益生菌治疗沙门氏菌肠炎的研究显示益生菌在维生素D 受体(VDR)缺陷小鼠中没有显示出保护作用,而在野生型小鼠中可抑制沙门氏菌诱导的炎症和损伤。
VDR基因消融降低肠道中DEFA5和MMP7表达,肠道通透性增加,肠道内环境紊乱,维生素D参与ß-防御素-2的表达,通过ß-防御素-2的表达与肠道菌群相互作用,改善肠道环境以及影响肠道炎症,并且存在剂量关系。溃疡性结肠炎急性期用维生素D能够缓解急性期症状,改善肠道粘膜和肠道菌群。此外,推测益生菌对个体起效可能依赖于VDR 基因。
以上,我们看到在各种肠道疾病的条件下,补充维生素D在一定程度上能改善肠道菌群。那么在缺乏维生素D的健康个体中,补充维生素D对肠道菌群是否有益?
近日,一项研究对80位健康维生素D缺乏的女性进行补充维生素D的治疗。

该研究主要是解决两个问题。
1. 补充维生素D对受试者的肠道菌群的影响
补充维生素D可显着增加肠道微生物多样性。具体而言,拟杆菌/厚壁菌的比例增加,益生菌类Akkermansia和双歧杆菌的丰度增加。拟杆菌属和普氏杆菌属显著变化,表明补充后肠型发生了变化。
2. 评估受试者菌群是否与他们的无反应状态有关。
注:无反应状态指一部分人群口服高剂量的维生素D也无效。
已有研究表明,Bacteroides acidifaciens可以预防肥胖和提高胰岛素敏感性,也是大肠中促进IgA抗体产生的主要共生菌之一。在该研究中,无反应者中较低的Bacteroides acidifaciens以及补充维生素D后的额外消耗表明,这种细菌可能与补充维生素D的反应有关。
因此,研究人员有这样一个设想,维生素D的补充促进有益菌生长,以维持免疫微生物的体内平衡。当然,需要开展更大队列的实验研究,以充分代表有反应者/无反应者,来证实该研究的发现。
6.1 鱼 类
鲑鱼
鲑鱼是一种受欢迎的脂肪鱼,也是维生素D的重要来源。

根据USDA食物成分数据库显示,一份3.5盎司(100克)大西洋鲑鱼每份含526 IU维生素D。
一些研究发现野生鲑鱼的含量更高,每份高达1300 IU。而鲑鱼养殖平均含量为250 IU。
鲱鱼
鲱鱼也是维生素D的最佳来源之一。每100克新鲜大西洋鲱鱼可提供216 IU,如果不是新鲜的鱼,腌鲱鱼也是维生素D的良好来源,每100克提供112 IU。

其他鱼类包括沙丁鱼,大比目鱼和鲭鱼等,都是维生素D的良好来源。
金枪鱼罐头
每包100克可含多达268 IU的维生素D,然而金枪鱼罐头中一般含有甲基汞(甲基汞是许多鱼类中发现的一种毒素)。可以选择淡金枪鱼(通常比白金枪鱼更好),每周不超过170克,可以防止甲基汞过多。

鱼肝油
鱼肝油每匙(4.9毫升)含维生素D约448 IU。同时也富含维生素A,omega-3脂肪酸等,但是不要服用过量。
6.2 蛋 黄
不吃鱼的人应该知道,海鲜不是维生素D的唯一来源。全蛋是另一种良好的来源,也是一种营养丰富的食品。
鸡蛋中的蛋白质大多在蛋清中,而脂肪、维生素和矿物质主要在蛋黄中。一颗标准的蛋黄含有37 IU的维生素D。

蛋黄中的维生素D含量取决于阳光照射和鸡饲料中维生素D的含量。当使用相同的饲料时,在阳光下漫游的牧场饲养的鸡所产卵的水平要高3-4倍。
从饲养的鸡中选择鸡蛋,或者从市场上购买富含维生素D的鸡蛋,可以满足日常需求。
6.3 蘑 菇
蘑菇是维生素D的唯一良好植物来源。蘑菇有点像人类,蘑菇在暴露于紫外线时也可以合成维生素D。不同的是,蘑菇产生维生素D2,而动物产生维生素D3。
某些野生蘑菇每100克包装含维生素D高达2300 IU。总的来说,只有野生蘑菇或经紫外线照射的蘑菇才是维生素D的良好来源。

6.4 强化食品
注:强化食品指向食品中添加一定量的食品营养强化剂,以提高其营养价值。
牛奶
有些牛奶中富含维生素D。每杯牛奶(237毫升)通常含有115-130 IU。
豆浆
通常会含维生素D以及牛奶中常见的其他维生素和矿物质。一杯(237毫升)通常含有107-117 IU的维生素D。
橙汁
一杯(237毫升)强化橙汁,包括高达100 IU的维生素D,当作早餐开启一天的活力。

燕麦片
半杯(78克)可提供维生素D量为54–136 IU。
注意以上食物中维生素D计算是强化食品中的含量。
最后,要提醒大家的是,“过犹不及”。维生素D虽然好,但不能补充过量。一旦维生素D过量,就会对身体带来伤害。
大剂量的维生素D服用一开始没有症状,但过几个月或几年就开始显现了,严重的情况下甚至可能导致肾脏衰竭和动脉钙化。
每天摄入1000-4000IU(25-100微克)的维生素D应该足以确保大多数人的最佳血液水平。
附 录 : 特殊人群维生素D补充剂量
维生素D缺乏引起的骨软化需要起始每日使用3000-40000IU的维生素D,随后使用每日400IU的维持剂量。
肠道吸收不良或肝脏疾病引起的维生素D缺乏通常需要每日使用40000IU(成人)治疗,每日10000-25000IU(儿童)。
甲状旁腺功能减退引起的低钙血症需要使用每日最多100000IU维生素D
——参考自:《DK家庭用药指南》
相关阅读:
参考文献:
Sizar O, Khare S, Goyal A, et al. Vitamin D Deficiency. [Updated 2020 Jul 21]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2020 Jan
Nair R, Maseeh A. Vitamin D: The “sunshine” vitamin. J Pharmacol Pharmacother. 2012 Apr;3(2):118-26.
Holick MF. Vitamin D: important for prevention of osteoporosis, cardiovascular heart disease, type 1 diabetes, autoimmune diseases, and some cancers. South Med J. 2005 Oct;98(10):1024-7.
Czernichow S, Fan T, Nocea G, Sen SS. Calcium and vitamin D intake by postmenopausal women with osteoporosis in France. Curr Med Res Opin. 2010 Jul;26(7):1667-74. [PubMed] 4.
Naeem Z. Vitamin d deficiency- an ignored epidemic. Int J Health Sci (Qassim). 2010 Jan;4(1):V-VI.
Singh Parul,Rawat Arun,Alwakeel Mariam et al. The potential role of vitamin D supplementation as a gut microbiota modifier in healthy individuals.[J] .Sci Rep, 2020, 10: 21641.
Thomas MK, Lloyd-Jones DM, Thadhani RI, Shaw AC, Deraska DJ, Kitch BT, Vamvakas EC, Dick IM, Prince RL, Finkelstein JS. Hypovitaminosis D in medical inpatients. N Engl J Med. 1998 Mar 19;338(12):777-83.
Gröber U, Kisters K. Influence of drugs on vitamin D and calcium metabolism. Dermatoendocrinol. 2012 Apr 01;4(2):158-66.
Pereira-Santos M, Costa PR, Assis AM, Santos CA, Santos DB. Obesity and vitamin D deficiency: a systematic review and meta-analysis. Obes Rev. 2015 Apr;16(4):341-9.
Elliott ME, Binkley NC, Carnes M, Zimmerman DR, Petersen K, Knapp K, Behlke JM, Ahmann N, Kieser MA. Fracture risks for women in long-term care: high prevalence of calcaneal osteoporosis and hypovitaminosis D. Pharmacotherapy. 2003 Jun;23(6):702-10.
Kennel KA, Drake MT, Hurley DL. Vitamin D deficiency in adults: when to test and how to treat. Mayo Clin Proc. 2010 Aug;85(8):752-7
Palacios C, Gonzalez L. Is vitamin D deficiency a major global public health problem? J Steroid Biochem Mol Biol. 2014 Oct;144 Pt A:138-45.
Zadka K, Pałkowska-Goździk E, Rosołowska-Huszcz D. The State of Knowledge about Nutrition Sources of Vitamin D, Its Role in the Human Body, and Necessity of Supplementation among Parents in Central Poland. Int J Environ Res Public Health. 2018;15(7):1489. Published 2018 Jul 14.
Sordillo JE, Zhou Y, McGeachie MJ, et al. Factors influencing the infant gut microbiome at age 3-6 months: Findings from the ethnically diverse Vitamin D Antenatal Asthma Reduction Trial (VDAART) [J]. J Allergy Clin Immunol, 2017, 139(2):482-491.
Luthold RV, Fernandes GR, Franco-de-Moraes AC, et al. Gut microbiota interactions with the immunomodulatory role of vitamin D in normal individuals[J]. Metabolism, 2017,69:76-86.
Mai V , Mccrary Q M , Sinha R , et al. Associations between dietary habits and body mass index with gut microbiota composition and fecal water genotoxicity: an observational study in African American and Caucasian American volunteers[J]. Nutrition Journal, 2009, 8(1):49.
White JH. Vitamin D deficiency and the pathogenesis of Crohn’s disease[J]. Steroid Biochem Mol Biol, 2018,175:23-28.
Schäffler H, Herlemann DP, Klinitzke P, et al. Vitamin D administration leads to a shift of the intestinal bacterial composition in Crohn’s Disease patients, but not in healthy controls[J]. Journal of Digestive Diseases, 2018, 19(4): 225- 234.
Ooi JH, Li Y, Rogers CJ, et al. Vitamin D regulates the gut microbiome and protects mice from dextran sodium sulfate-induced colitis[J]. J Nutr, 2013,143(10):1679-1686.
Shang M, Sun J. Vitamin D/VDR, Probiotics, and Gastrointestinal Diseases[J]. Curr Med Chem, 2017,24(9):876-887.
李子傲. 母乳喂养对肠道菌群的影响[J]. 营养与健康,2016, 10( 8) : 97-99.
汪英,袁莉,李广利,等. 肥胖与血清维生素D 水平的关系研究[J]. 中华内分泌代谢杂志,2011,27 ( 7) :589-590.
祝海波,周苗苗,王海,张晓宇.婴儿早期维生素D补充水平对生长发育和肠道菌群的影响[J].中国食物与营养,2018,24(12):87-89.
李苹,昌雪莲,尚煜,刘雅静,陈晓宇,梁爱民,齐可民.婴儿早期维生素D补充对生长发育及肠道菌群的影响[J].公共卫生与预防医学,2018,29(01):12-16.
杜琳. 维生素D介导的Th17/Treg细胞稳态调节对孤独症大鼠肠道菌群失调作用的研究[D].吉林大学,2019.

谷禾健康
“ 在人类适应中整合宿主-微生物群的相互作用可以提供新的方法来提高我们对人类健康和进化的理解。”

面对快速的环境变化,生物如何适应新环境是进化生物学中的一个核心问题。
刚刚,来自德国马克斯·普朗克研究所研究人员在science发表了最新文章:

回顾了人类适应新环境的例子,这些例子表明宿主基因和微生物群之间的相互作用。提出宿主机制可以在本地适应过程中替代或补充有益的微生物群功能。寻找微生物群与人类遗传适应性有关的其他例子。
当人类在基因上适应新环境时,他们的微生物群也可能参与这个过程。微生物可以比宿主进化得更快,这使它们能够对环境变化做出快速反应。微生物群还过滤宿主的环境,从而改变宿主的选择性压力。
图1 人类及其微生物的局部适应

举例说明适应性宿主等位基因和适应性微生物群功能之间的相互作用。
选择压力的差异,包括饮食、气候和病原体暴露,导致了人类群体在性状上的差异,与消化、生理和免疫相关。这些差异是由跨多代发生的局部遗传适应造成的,与宿主一生中免疫系统的适应或生理适应是有区别的。
在许多情况下,主要的遗传变异体被确定为如下:
乳糖消化所需的乳糖酶:(由基因区域LCT编码)突变和截短
缺氧诱导因子2α:(由PAS1编码)在高原适应中产生作用
β-珠蛋白基因单倍型或镰状细胞性状:(由血红蛋白编码)在疟疾抗性中产生作用
此外,人类微生物群的组成和功能变化已被广泛描述。Amato等人最近回顾了可能影响人类本地适应性的微生物群的有益功能,例如微生物群增强营养和预防传染病。然而,在人类局部适应过程中,宿主适应性等位基因和适应性微生物群功能之间的特定相互作用仍有待研究。
接下来的两小节,作者详细研究了LCT–双歧杆菌和AMY1–瘤胃球菌之间的相互作用。在这些例子中,适应性宿主等位基因和适应性微生物功能是相联系的。
婴儿的主要能量来源于乳糖。在上消化道(GIT,下同),宿主酶乳糖酶将乳糖分解成葡萄糖和半乳糖。饮食中没有牛奶,乳糖酶的生产就会停止。乳糖酶非持续性LNP(乳糖不耐受)是祖先的条件。
注:由于乳糖酶活性的不同,人群中存在三种基因型,即纯合乳糖酶持续性(LP)、纯合乳糖酶非持续性(LNP)和杂合子(heterozygotes)。LNP是人群中最常见的表型。因乳糖酶的相对或绝对缺乏而导致的以腹泻为主的相关消化道症状的现象称之为乳糖不耐受。LNP是原发性乳糖不耐受的生理基础。
不同大陆的动物驯化(2500年到10000年前)和对非人类牛奶的反复消费导致了强烈的选择压力,使乳糖酶的生产持续到成年,即乳糖酶持久性LP(乳糖耐受)。LP是突变,在有长期放牧和挤奶历史的人群中常见。
宿主和微生物共争乳糖
许多肠道微生物群中常见的微生物可以使用β-半乳糖苷酶来裂解乳糖,然后发酵,乳糖可能被乳酸菌隔离。不管如何加工,微生物利用乳糖的副产物给宿主产生的能量更少。这意味着宿主和微生物在争夺乳糖,宿主有强烈的动机去战胜微生物。
乳糖耐受——宿主赢
乳糖耐受宿主胜过微生物,因为乳糖酶在微生物生物量较低的上层消化道中具有活性。
乳糖不耐受——微生物赢
相比之下,乳糖不耐受宿主主要在较低的消化道中获取微生物乳糖代谢的低能量密度产物。
因此,未消化的乳糖进入结肠可以被认为是一种需要微生物酶来处理的纤维形式。
就像结肠中的纤维降解一样,来自发酵的微生物,和它们的发酵产物[短链脂肪酸(SCFAs)]组成,虽然微生物群吸收了乳糖的一部分能量,但乳糖不耐受宿主的另一种选择是完全失去能量来源。
LCT位点的相同变异与肠道微生物群中双歧杆菌的相对丰度有关。
这种关联是迄今为止微生物组全基因组关联研究最一致的信号,该关联取决于牛奶消费量。
乳糖耐受基因型和双歧杆菌相对丰度之间的反比关系支持了哺乳动物乳糖酶和细菌β-半乳糖苷酶直接竞争乳糖的观点。
双歧杆菌是重要的乳糖降解菌,可能在产奶动物驯化后和选择乳糖耐受等位基因前帮助成年宿主从牛奶中提取能量。在畜牧业兴起后,乳糖代谢细菌可能在缺乏乳糖耐受等位基因的情况下对宿主的适应性产生积极影响。后来,有益的宿主等位基因出现并取代了微生物群的功能,在一些人群中变得几乎固定。
双歧杆菌(或功能过剩的微生物)产生的β-半乳糖苷酶也可能通过降低LCT基因型之间的适应度差异而减轻了对乳糖耐受等位基因的选择压力。
此外,如果微生物摄入乳糖能最大限度地减少与乳糖消化不良相关的疾病后果,并促进牛奶摄入,从而带来额外的益处,则微生物群可能会减轻能量权衡并维持人类群体中的乳糖酶多态性。
乳糖的例子表明,由微生物组和宿主基因组编码的活性可能特别容易受到宿主和微生物组之间竞争的影响。
与乳糖相反,人类饮食中的各种淀粉为宿主提供了一种机会,在较高的消化道中获得一种带有淀粉酶的淀粉,而在较低的消化道中为微生物群消化留下抗性淀粉。
事实上,淀粉有许多不同的形式,包括难以被宿主淀粉酶降解的结构,淀粉酶将淀粉分解成葡萄糖亚基。宿主和微生物群分配淀粉底物的程度可能取决于宿主基因型。
由AMY1基因编码的唾液淀粉酶在口腔中开始淀粉分解为葡萄糖的过程。唾液淀粉酶活性的个体差异与AMY1基因的拷贝数呈正相关,通常在2到15之间。
与AMY1 拷贝数含量低的个体相比,AMY1 拷贝数含量高的个体在食物到达较低的消化道及其微生物群之前,更彻底地消耗了给定饮食中对淀粉酶敏感的淀粉(图2,C和 D)。
图2 宿主基因型和微生物群之间联系的例子
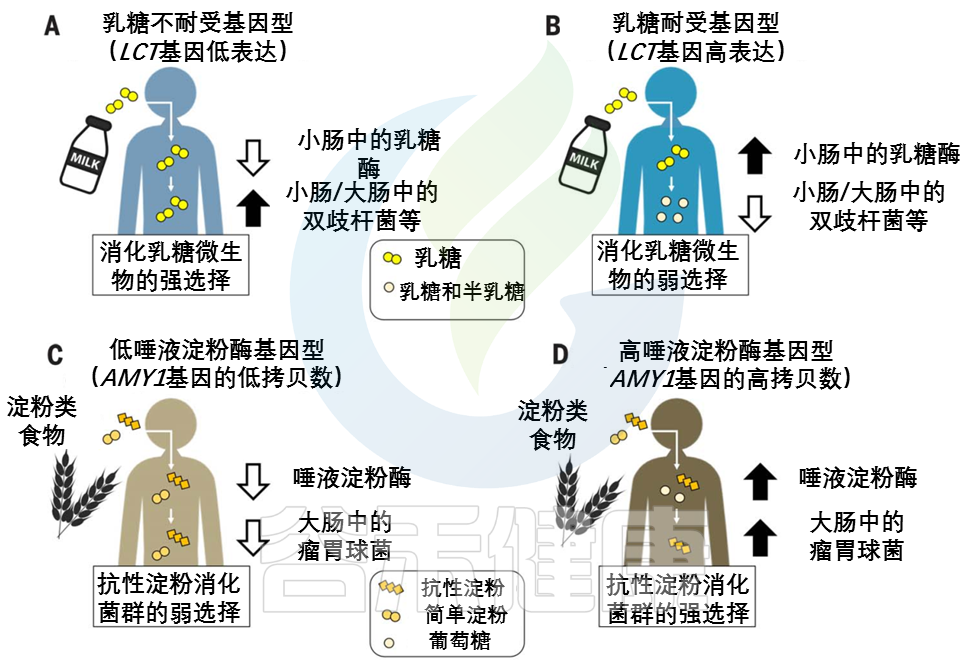
在(A) 乳糖不耐受宿主和(B) 乳糖耐受宿主中,乳糖酶基因型与双歧杆菌相对丰度呈负相关关系。
AMY1基因拷贝数(CN)和瘤胃球菌相对丰度之间的正相关性被描述为(C)低AMY1 拷贝数宿主和(D)高AMY1 拷贝数宿主。
不同类型的糖分以不同的形状和颜色显示。
AMY1 拷贝数为2是祖先的情况:AMY1 拷贝数在人类中的扩展可能是由从低淀粉饮食到高淀粉饮食的饮食转变所驱动的。
随着农业的发展,唾液淀粉酶水平的提高可能会促进高淀粉饮食人群的健康。与乳糖耐受的进化相似,宿主淀粉酶通过其比微生物发酵产物更大的能量产量而具有适应性优势。
具有AMY1高拷贝数的宿主还能从结肠微生物对抗性淀粉的强化降解中获益。
抗性淀粉发酵中的重要属——Ruminococcus
与AMY1低拷贝数个体的肠道微生物群相比,高AMY1拷贝数宿主的肠道微生物群富含瘤胃球菌属(Ruminococcus)。
在AMY1 低拷贝数宿主中,肠道细菌优先发酵淀粉酶敏感型淀粉,宿主不能使用这些淀粉,因为它们的唾液淀粉酶水平较低。
乳球菌,或功能冗余的微生物,可能通过放大AMY1高拷贝数和低拷贝数个体之间的相对适应度差异,增强了对AMY1 拷贝数扩增的选择压力,并可能增强了宿主基因-微生物的关联(图2,C和D)。因此,这种动态可能导致基于宿主基因型的微生物补充。
以上,作者提出宿主机制可以在本地适应过程中替代或补充有益的微生物群功能。接下来的章节,作者寻找微生物群与人类遗传适应性有关的其他例子,其中适应性的遗传基础得到了很好的描述。
有新的证据表明,微生物参与了其他几种人类遗传适应(图3),之前在这些适应中,微生物的作用没有得到充分认识。
图3 人类的局部适应性与微生物群有关
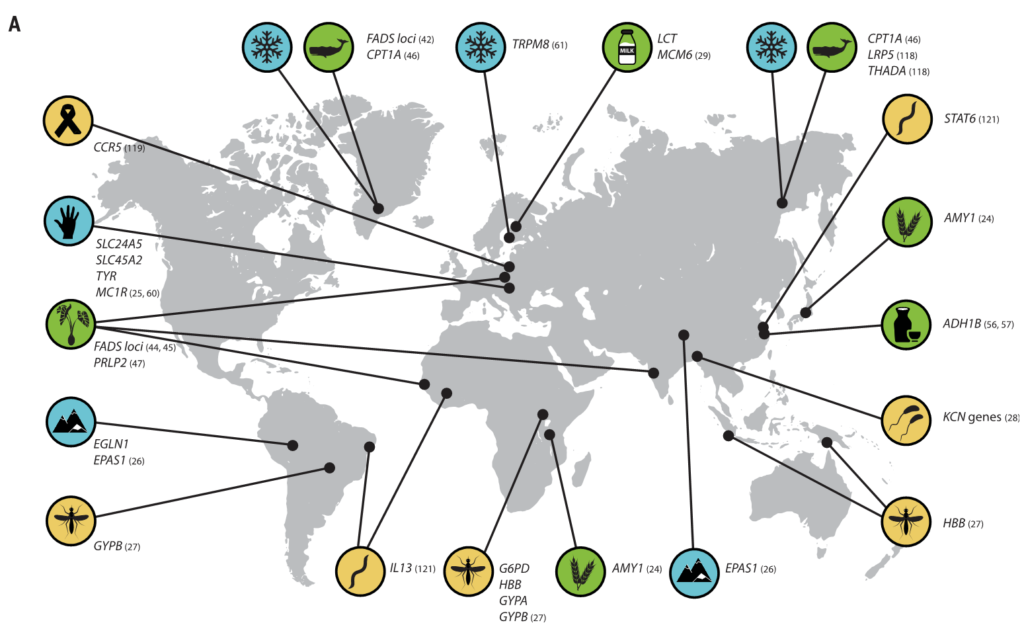
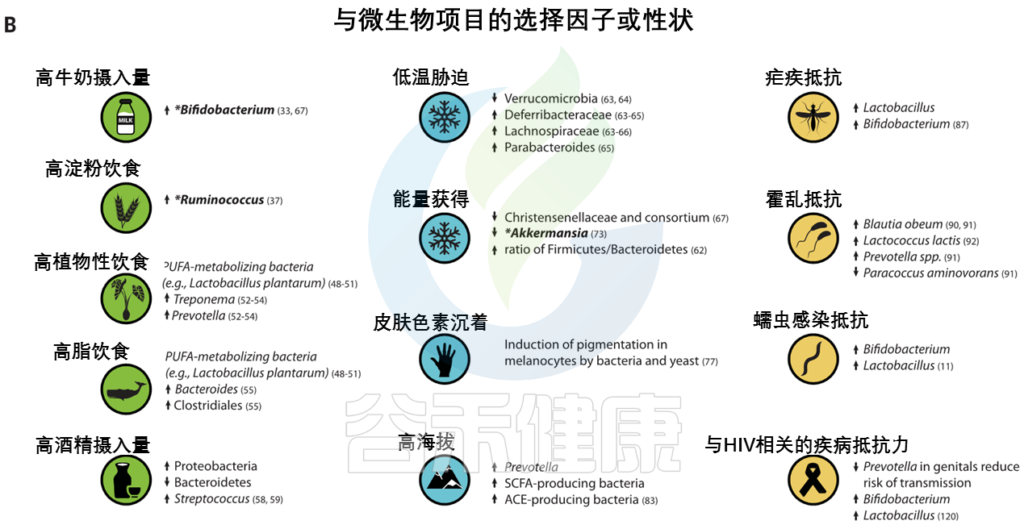
(A) 与适应性性状相关的人类基因位点的全球分布。这些符号对应于(B)中的选择因子和/或特征。
向上和向下的箭头分别表示微生物类群相对丰度与阳性或阴性相关的趋势。
微生物群的研究群体与宿主基因位点的研究群体不匹配.
星号和黑体字表示正向选择下与宿主基因位点相关的微生物类群: LCT和双歧杆菌,AMY1和Ruminococcus,PLD1和Akkermansia
乳糖和淀粉消耗可能是第一个已知的与饮食相关的基因-微生物相互作用的例子,可以进一步研究其在人类适应中的作用,但其他例子可能会出现。例如,长链多不饱和脂肪酸(LC-PUFAs)是关键的营养物质,可以直接从动物性饮食中获得,或者使用宿主衍生的酶从植物衍生的前体中合成。
脂肪酸代谢相关基因【包括脂肪酸去饱和酶(FADS)、肉碱棕榈酰转移酶1A (CPT1A) 和胰脂肪酶相关蛋白2 (PLRP2)】的正向选择证据与多种人群的饮食适应有关。
FADS基因区域的祖先等位基因与食用植物性食物的人群相关,衍生等位基因与当前和历史上偏爱高脂肪食物的人群相关。属于至少10个门的细菌似乎能合成长链多不饱和脂肪酸。
在人类肠道微生物中,植物乳杆菌和其他细菌已被证明能代谢植物来源的前体,并影响多不饱和脂肪酸来源的代谢物在体外和体内的生物利用度。分别通过植物性饮食或动物性饮食富集的人类肠道微生物 。
类似地,酒精脱氢酶1B (ADH1B)位点在人类中显示出正向选择的证据,并与脱脂摄入有关,人们认为这是伴随着农业的发展而产生的。
人和细菌的酶可以将乙醇转化为乙醛;肠道微生物群的变异与饮酒和相关疾病有关。
这些观察表明:
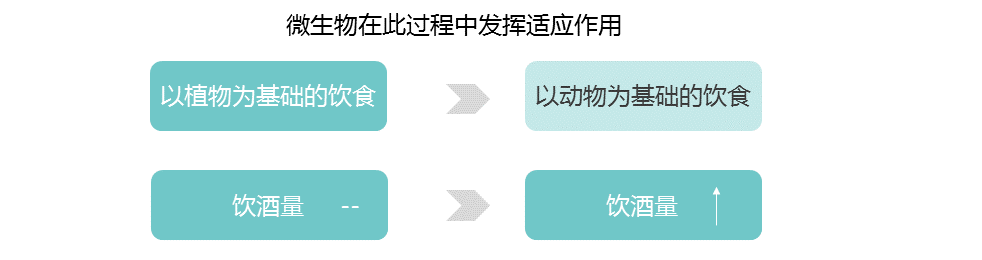
我们已经适应了自己所在地的气候。生活在高纬度地区的人面临冷胁迫,与温度感应(TRPM8)和能量收获(如FADS和CPT1A)相关的基因与气候相关的适应有关。
气候是参与常见代谢紊乱基因的重要选择压力
人类微生物群也因纬度而异,并可能通过提高从饮食中提取能量和随后储存脂肪的效率来促进与气候相关的适应。微生物群中的一些变异与环境温度和宿主体型有关。对小鼠的实验研究表明,仅环境温度的变化就能引起肠道菌群的变化。
低温
低温暴露宿主的微生物表现出更高的吸收能力和能量收获能力,以及棕色脂肪组织产生的热量增加。然而,微生物介导的产热和TRPM8多态性在人类中是否相关仍有待确定。
· Christensenellaceae
人类肠道中最紧密和高度可遗传的分类单元,细菌家族christensenellaceae,它与较低的体重指数(BMI) 和代谢紊乱有关。甲烷菌科、脱水杆菌科 和Tenericutes在多个人类群体中也表现出遗传性,并与Christensenellaceae共存。
最近的证据表明,这些菌群之间的代谢相互作用可能会影响宿主对代谢物的利用。
GWASs没有发现显著的宿主遗传变异。这表明,一些明显可遗传的分类群可能具有多基因性质,其中许多基因的影响很小。
· Akkermansia
Akkermansia是一种微生物降解菌,也与体重指数有关,在多项研究中显示为一个可遗传的分类群。Akkermansia的相对丰度与PLD1 中的宿主遗传变异有关,PLD1是之前与BMI相关的基因。PLD1显示了非洲人群中正向选择的证据,但选择剂尚不清楚。
太阳辐射
生活在低纬度地区的人群面临高水平的太阳辐射,与皮肤色素沉着相关的基因是人类本地适应的典型例子(图3)。
皮肤中的黑色素保护皮肤免受紫外线辐射,其抗菌特性被认为是皮肤色素沉着纬度梯度的替代假设。因此,人类黑素细胞中皮肤色素沉着的增加可以通过暴露于微生物产物如脂多糖来诱导。
Guéniche等人报道了乳酸菌对皮肤的保护作用,紫外线照射后的全身免疫反应。鉴于肠道微生物群在调节皮肤内环境稳定中的新兴作用,进一步的研究可能有助于确定色素沉着基因与皮肤和肠道微生物之间的潜在相互作用。
高海拔环境
高海拔环境也带来了许多生理挑战,包括缺氧应激,这也可能被微生物群改变。在高海拔环境中,已经鉴定出与氧呼吸和血液循环相关的多种适应性特征和基因,这些特征和基因通常涉及血压的调节(图3)。
最近,肠道微生物群被证明通过产生短链脂肪酸来调节血压,短链脂肪酸有可能在高海拔环境中帮助调节血压。
包括丁酸盐在内的短链脂肪酸的产生影响小鼠缺氧诱导因子的活性,这也是多个人类群体中参与高海拔适应的关键途径。作用于肾素-血管紧张素系统的细菌血管紧张素转换酶(ACEs)的产生也可能改变高海拔环境中的血压。细菌ACEs已被证明能在体外转化哺乳动物血管紧张素。
病原体被认为是人类适应性进化的主要驱动力之一(图3),微生物群与病原体防御和免疫之间的界面已被广泛研究。
人类本地适应的一个典型例子是疟疾抗性的遗传基础,由疟原虫属的几种寄生原生动物引起。皮肤和肠道微生物可能影响疟疾的不同阶段。
相同细胞因子相关基因的遗传变异,包括IL10、IL12、肿瘤坏死因子、干扰素α和干扰素γ,与疟疾严重程度和微生物群组成的变异有关。
Ippolito等人总结了微生物群在疟疾抗性中可能扮演的多重角色的几种假说:
1 皮肤细菌产生可改变蚊子媒介吸引力的挥发性化合物
2 由肠道微生物诱导的抗原表达,该抗原表达诱导针对疟原虫传播的保护性抗体
3 影响血液阶段疟原虫感染严重性的微生物群的差异
其他可能受到微生物群和宿主遗传变异组合影响的感染包括霍乱、艾滋病毒和蠕虫感染。
霍 乱
霍乱毒素针对几个带有显示选择信号的基因的离子通道。已经在人群中鉴定出与霍乱易感性和从霍乱中恢复相关的肠道微生物分类群。实验研究已经确定了提供定殖抗性和抑制霍乱弧菌毒性的细菌分类群。
鉴于在胃肠疾病中离子通道的表达和功能经常被改变,探索微生物群和离子通道基因之间的联系对于治疗或预防腹泻疾病可能是有价值的。
艾滋病毒和蠕虫感染
阴道微生物群在艾滋病毒的获得和传播中起作用,
肠道微生物群与蠕虫相互作用并改变疾病结果。
对研究充分的宿主适应性等位基因和新兴有益微生物群功能之间的相互作用的进一步研究可能会揭示以前未知的抗病机制。
这里提到的所有宿主基因-微生物例子(图3)仍处于研究的早期阶段。但是在每一种情况下,微生物群都有可能通过改变适应性景观来影响宿主的进化。这些微生物可能改变了宿主的环境,导致宿主表型选择压力的改变。
在未来关于适应性宿主基因型和适应性微生物群功能之间关系的研究中,GWASs、候选基因方法或使用基因敲除模型的移植实验可以帮助确定维持有益微生物群功能的宿主遗传机制,并测试微生物群的替代和补充是否是适应性的一般特征。
基因研究已经将越来越多的群体纳入考虑范围,微生物群的特征研究也是如此。但到目前为止,包括宿主遗传学和微生物群的研究,除了少数例外,主要是在西方人群中进行的。
需要将人类基因研究的范围扩大到更广泛的人群和地点,其中包括微生物组成分。这些类型的研究结果将测试当前发现的普遍性,并描述更多微生物介导的适应情况。
还可以进一步研究特定分类群的遗传力,以更好地表征潜在的遗传多态性。遗传性既适用于分类群,也适用于微生物功能,因此通过宏基因组学将与人类遗传变异相关的性状从分类群扩展到功能群,将有助于进一步阐明微生物群在人类适应中的作用。
在这里关注的是宿主(主要是人类)的进化,但是微生物也在快速进化以适应它们的宿主环境。在适应性进化的背景下,对宿主基因组和微生物群之间相互作用的更好理解将阐明人类和其他动物及其微生物群是如何适应不断变化的世界的。
参考文献:
Suzuki Taichi A,Ley Ruth E,The role of the microbiota in human genetic adaptation.[J] .Science, 2020, 370
Bayless Theodore M,Brown Elizabeth,Paige David M,Lactase Non-persistence and Lactose Intolerance.[J] .Curr Gastroenterol Rep, 2017, 19: 23.
S. Fan, M. E. B. Hansen, Y. Lo, S. A. Tishkoff, Going global by adapting local: A review of recent human adaptation. Science 354, 5 4–59 (2016). doi: 10.1126/science.aaf5098; pmid: 27846491
M. Fumagalli et al., Greenlandic Inuit show genetic signatures of diet and climate adaptation. Science 349, 1343–1347 (2015). doi: 10.1126/science.aab2319; pmid: 26383953
T. A. Suzuki, F. M. Martins, M. W. Nachman, Altitudinal variation of the gut microbiota in wild house mice. Mol. Ecol. 28, 2378–2390 (2019). doi: 10.1111/mec.14905; pmid: 30346069

谷禾健康
2020年只剩最后一个月,全球范围内,疫情却未见消停。
新冠肺炎(COVID-19)不断威胁着全人类的健康,而对于癌症患者,更是雪上加霜。
来自意大利研究人员Angioletta等人带着这样的问题,对新冠和某些癌症的关系开始进行探索,回顾现有数据并且对乳腺癌(BC),肠道菌群(GM)和COVID-19之间的关系进行推测。
首先,他们发现新冠疫情期间的一项多中心研究指出,与没有癌症的COVID-19患者相比,感染COVID-19的癌症患者病情更严重,与没有患癌症的COVID-19患者相比,死亡率增加了近三倍。
现有的研究已经证实,病毒SARS-CoV-2利用ACE2受体进入,利用TMPRSS2进行S蛋白启动。
最近的一项Meta分析报告称,肺癌和结直肠癌患者更容易受到SARSCoV-2感染:支持生物信息学的数据显示,在这些癌症类型中,ACE2和TMPRSS2的mRNA表达水平均有所增加。
从上面的结论看来,也就是说癌症患者的生理环境恰好为新冠病毒的侵入提供了一个机会。然而,如果只是小样本量和仅包含七种类型的癌症,则解释结论需要更为谨慎。
所以到目前为止,可用的数据还不能完全确定是否有一种癌症类型更容易受到新冠肺炎病的影响,因为病例具有广泛的异质性,并且数据没有按性别进行分类和分析。
事实上,新的全球观察表明,与男性相比,新冠肺炎女性感染和死亡的风险较低。
这样的结论让研究人员更想探寻其中的缘由。
一般来说,基于不同的先天免疫、类固醇激素和与性染色体相关的因素等因素,女性不太容易受到新冠肺炎病的影响。
随机X染色体失活的影响是可能的解释之一,因为ACE2是由位于X染色体上的ACE2基因编码的。男性细胞总是表达单个ACE2等位基因,女性拥有一种更有效的ACE2受体,平均只存在于一半的细胞中。
虽然这种机制可能会限制感染SARS-CoV-2病毒的易感性,使女性具有相对的抵抗力,但这可能不足以解释女性对感染的不同易感性。
对此,Angioletta等人又找到了新的发现。

在最近对ACE2表达和活性的机制差异的综述中,作者报道了小鼠模型中ACE2活性在雄性肾脏中更大,这种性别差异是由雌二醇降低ACE2活性而引起的,而与性染色体补体无关。
性激素能够影响先天和适应性免疫反应:
雄激素具有抗炎作用;
雌激素既有促炎作用,也有抗炎作用。
正如这篇关于前列腺癌和SARS-CoV-2之间的生物相互作用和分子联系的综述所总结的那样,雌激素通过直接改变CD16表达来抑制促炎性IL-6的水平,并且可以影响自然杀伤细胞的水平。
注:自然杀伤细胞(natural killer cell,NK)是机体重要的免疫细胞,具有广谱的抗肿瘤细胞作用。

在武汉进行的一项横断面研究中,假设了雌激素的关键作用:丁及其同事分析了年龄小于60岁的女性患者的月经状况、与免疫和炎症相关的女性激素和细胞因子与严重程度和临床结局之间的相关性。雌二醇与感染的严重程度呈负相关,作者发现绝经是新冠肺炎病患者的独立危险因素。
可见雌激素的作用显得越来越重要,但雌激素具体是如何影响激素依赖性癌症的呢?
雌激素 & 肠道菌群

一项队列研究结果的分析强调了这个问题的相关性,该研究报告了来自新冠肺炎和癌症协会登记数据库的数据,乳腺癌 (21%)和前列腺癌(16%)这两种强烈依赖激素的癌症,是2020年3月至4月累积的1018例COVID-19病例中最常见的癌症类型。
该结果有点出乎意料。
我们再看该研究人群的年龄情况:中位年龄为66岁(IQR 57-76)【整体年龄偏大】,75岁及以上279例(30%),男性468例(50%)。
研究人员认为,这一发现支持了肠道菌群成分可能是解释这种联系的一个因素的假设。
原因:已有的数据表明,肠道菌群构成在物种和功能的流行方面存在性别差异,部分是由性激素驱动的:男性和绝经后女性的雌激素水平与肠道菌群丰度和多样性直接相关,而绝经前女性则没有相关性。
此外,肠道菌群能够调节血清雌激素水平,促进某些种类的菌群增殖;Plottel和Blaser将肠道细菌基因(其产物能够代谢雌激素)的集合定义为“estrobolome” (雌激素组)。
肝结合雌激素导致共轭雌激素和共轭雌激素代谢物分泌到胃肠道,在胃肠道中,它们被细菌来源的β-葡萄糖醛酸酶、葡糖苷酶和羟基类固醇脱氢酶转化为游离形式。如果雌激素组富含具有较高解偶联和羟基化酶活性的细菌,则将导致游离雌激素的相对水平更高。
在人类胃肠道中,最重要的β-葡萄糖醛酸酶编码基因称它为GUS基因;大约有112种新的GUS基因被鉴定出来,并被分为六类,分别在四个细菌门表达,即拟杆菌门、厚壁菌门、疣微菌门和变形菌门。其中,拟杆菌属的GUS酶的丰度和多样性最高。
肠道菌群 & 雌激素 & 乳腺癌
乳腺癌是女性中最常见的肿瘤,积累的数据表明发病率不断上升: 2018年,28个欧盟国家经年龄调整的乳腺癌年发病率估计为144.9/100000,死亡率为32.9/100000。
乳腺癌有许多危险因素,但一些研究分析了乳腺癌和胃肠道微生物对雌激素的依赖作用之间的复杂关系。

在2018年发表的一项研究中,与对照组相比,乳腺癌病例与IgA阳性和IgA阴性肠道菌群具有显著的雌激素独立相关性。
这些发现表明,乳腺癌风险可能通过IgA阴性菌群的雌激素肠肝循环和IgA阳性菌群的免疫介导途径受到影响。
事实上,当评估具有不同临床特征的乳腺癌患者中肠道菌群的组成时,双歧杆菌和Blautia的绝对数量以及柔嫩梭菌Faecalibacterium prausnitzii和Blautia的比例,会根据癌症的临床分期而有所不同。
与没有乳腺癌的个体相比,在乳腺癌患者中观察到厚壁菌门与拟杆菌门的比例增加。
如果肠道菌群的改变影响乳腺癌患者的雌激素活性,我们对菌群多样性和COVID-19了解多少?
肠道菌群 & COVID-19

最近一篇论文报道了肠道厚壁菌与COVID-19严重程度之间的联系:
与新冠肺炎严重程度呈负相关的主要细菌种类有:Alistipes onderdonkii 和F. prausnitzii
拟杆菌通过阻止宿主进入ACE2而表现出抵抗SARS-CoV-2感染的潜在保护作用
虽然说这项探索性研究的样本量不大(15名新冠肺炎病患者、6名社区获得性肺炎受试者和15名健康个体的粪便样本),但这些结果表明,个体的肠道微生物群可能会影响其对SARS-CoV-2感染的易感性和反应。
鉴于所有这些证据,两个问题可能会出现:
新冠肺炎和乳腺癌之间的关系是什么?
雌激素会影响它吗?
目前已有的信息中,并没有关于乳腺癌患者感染的真实发生率的信息,也没有关于乳腺癌的哪种亚型最相关的信息。
Angioletta等人认为还没有足够的数据来得出明确的结论,但可以提出假设,并尝试着提出研究方法。
乳腺癌 & 雌激素 & 肠道菌群
已知在乳腺癌患者中,雌激素主要由脂肪组织产生,并通过雄激素前体的芳构化产生;芳香酶抑制剂能有效抑制芳香酶活性,抑制血浆和组织中的雌激素水平,是绝经后妇女雌激素受体阳性乳腺癌的主要治疗方法之一。
注:芳香酶抑制剂可以阻止从脂肪组织中产生雌激素,但对卵巢产生的雌激素没有影响。它不能阻止卵巢产生雌激素,因此仅对绝经后女性有效。
肠道菌群成分影响激素调节的各个方面。此过程会上调类固醇激素和细胞因子的循环水平,而这些激素和细胞因子会增加乳腺癌的风险和进展。已经发现菌群失调与绝经后乳腺癌患者有关,与绝经前无关。但是芳香酶抑制剂和肠道菌群之间相互作用的性质尚未完全确定。
选择性雌激素受体调节剂可以调节肠道菌群,但迄今为止,关于芳香酶抑制剂和肠道菌群成分之间关系的数据仍然缺乏。
肠道菌群 & 雌激素 & 乳腺癌 & 新冠
当然还有许多混杂因素需要考虑,如BMI、年龄、抗生素摄入量;所有这些都可能改变肠道菌群,增加这个情况的复杂性。此外,体重指数和衰老是众所周知的新冠肺炎风险因素。
Angioletta等人推测雌激素组通过调节雌激素水平在改变对新冠肺炎的易感性中起作用。
建立与乳腺癌和新冠肺炎相关的肠道菌群改变之间的关系很重要,病例数量多,需要一种个性化的方法来识别有风险的患者。
为了评估乳腺癌和新冠肺炎易感性之间的联系,他们提出了三个策略:
1 收集和储存生物样本(粪便和血浆),通过横断面研究评估肠道菌群构成和雌激素活性之间的联系
2 建立一个大型数据库,收集正在进行的治疗中乳腺癌的特征,特别是接受内分泌治疗的雌激素受体阳性亚型,以及COVID-19的发病率
3 使用机器学习算法对数据库进行分析,以发现雌激素、乳腺癌和COVID-19之间的相关性
根据现有的数据,乳腺癌患者可能通过雌激素水平的增加来保护其免受新冠肺炎病的侵害,这一模式目前正在讨论中,并出现了许多问题,包括内分泌治疗是否会干扰雌激素受体,是否会使患者更容易受到新冠肺炎感染。
在不久的将来,研究乳腺癌、雌激素和新冠肺炎之间的联系,确定激素敏感型癌症患者对新冠肺炎的风险增加,并制定策略通过改变他们的肠道菌群来预防,可能是一种有效方式,当然也可能是挑战。
当前迫切需要更可靠的流行病学、临床前和临床数据来指导日常实践。
参考文献:
Lasagna Angioletta, Zuccaro Valentina, Ferraris Elisa et al. COVID-19 and breast cancer: may the microbiome be the issue? [J] .Future Oncol, 2020.
Hoffmann M, Kleine-Weber H, Schroeder S et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 181(2), 271–280e278 (2020).
Dai M, Liu D, Liu M et al. Patients with cancer appear more vulnerable to SARS-COV-2: a multi-center study during the COVID-19 outbreak. Cancer Discov. 10(6), 783–791 2020
Wang B, Huang Y. Which type of cancer patients are more susceptible to the SARS-COX-2: evidence from a meta-analysis and bioinformatics analysis. Crit. Rev. Oncol. Hematol. 153, 103032 2020.
Conti P, Younes A. Coronavirus COV-19/SARS-CoV-2 affects women less than men: clinical response to viral infection. J. Biol. Regul. Homeost. Agents 34(2), 339–343 (2020).
Klein SL, Dhakal S, Ursin RL, Deshpande S, Sandberg K, Mauvais-Jarvis F. Biological sex impacts COVID-19 outcomes. PLoS Pathog.16(6), e1008570 (2020).
Chakravarty D, Nair SS, Hammouda N et al. Sex differences in SARS-CoV-2 infection rates and the potential link to prostate cancer. Commun. Biol. 3(1), 374 (2020).
Ding T, Zhang J, Wang T et al. Potential influence of menstrual status and sex hormones on female SARS-CoV-2 infection: a cross-sectional study from multicentre in Wuhan, China. Clin. Infect. Dis.Ciaa1022 doi:10.1093/cid/ciaa1022 (2020) (Epub ahead of print).
Kuderer NM, Choueiri TK, Shah DP et al. Clinical impact of COVID-19 on patients with cancer (CCC19): a cohort study. Lancet 395(10241), 1907–1918 (2020)
Zuo T, Zhang F, Lui GCY et al. Alterations in gut microbiota of patients with COVID-19 during time of hospitalization. Gastroenterology 159( 3), 944.e8–955.e8 (2020).
Augusto TV, Correia-da-Silva G, Rodrigues CMP, T eixeira N, Amaral C. Acquired resistance to aromatase inhibitors: where we stand.Endocr. Relat. Cancer 25(5), 283–301 (2018).
谷禾健康
朋友小杨是个科研人,每天按时吃饭,三餐规律,但在某次腹泻之后,小杨突然想起了医生的建议:做肠镜检查。
肠镜检查对于小杨而言是陌生而又新奇的,于是小杨开始飞速查阅相关文献。
用于检查肠道内部病变。对大肠息肉;大肠炎症性疾病如溃疡性结肠炎;慢性结肠炎;结肠癌等诊断有重要意义。
https://v.qq.com/x/page/r05303w8449.html谷禾肠镜过程介绍及肠镜检查后的注意事项动画_腾讯视频
谷禾肠镜过程介绍及肠镜检查后的注意事项动画_腾讯视频
查完后,小杨对肠镜检查有了大概的了解,需要先喝泻药排便清肠,才能进行正式的肠镜检查。与此同时,他脑海中又浮现出新的问题:
都说肠道中有大量的微生物,那么在大量排出粪便时会不会同时将肠道内的肠道菌群大量清除?
一次肠镜检查究竟对肠道菌群会有多大影响呢?
遗憾的是,目前文献较少提及此方面的内容。与我们进行沟通之后,小杨决定亲自做肠镜前后的肠道菌群检测,对比看看究竟会有什么变化。
肠镜检查前需要进行清肠准备,良好的准备可以为肠镜提供更好的视野,也会决定肠镜检查的效果。
目前常见的清肠方式主要采用复方聚乙二醇电解质方案,也就是泻药,主要成分为PEG4000,同时补充电解质。
好的清肠准备一般需要彻底排净粪便,直到排出无色清水便。
肠镜前准备会对肠道菌群产生怎样的变化呢?
是简单的等比例减少还是完全改变菌群构成?
满怀疑问的小杨开始了肠镜检查之旅。
检查当天,小杨开始服用聚乙二醇后,很快开始排便,状态为稀便,此时肠道尚未完全清空。等排空后,马上进行了肠镜检查。
做完肠镜检查,还没来得及思考太多,小杨就拿到了肠镜检查报告。
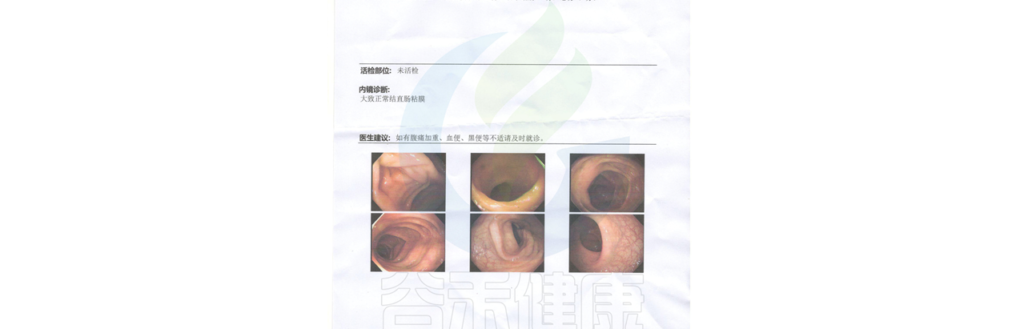
“ 肠镜结果显示正常 ”。
耳边传来了医生的话,小杨感到松了口气。
确保身体健康没问题了,小杨心中的疑惑却没有打消,是不是原本健康的肠道菌群会因此紊乱?
如果有不好的变化,我是不是应该做些什么帮助肠道菌群恢复?
小杨积极与我们进行沟通,希望我们给出相应的建议。我们基于文献及临床菌群干预经验给予饮食方面的小建议,这部分后面详述。
他按自己的理解结合我们的建议,采取了一些饮食干预措施。
当肠道菌群检测报告出来时,他很兴奋。
在整个过程包括前后,小杨完整地采集,总共完成了4次肠道菌群检测,分别为以下几个时间点:
· 肠镜检查前
· 检查当天服用聚乙二醇后首次排便
· 肠镜完成后首次排便
· 一周后
“怎么样?有变化吗?变化如何?”
…显然小杨很想知道结果。
为了便于理解,我们仔细帮他做了肠镜前一周和肠镜当天菌群的对比分析。
“一般人可没这待遇啊”,我们打趣道。
“来,看这些图表吧”…
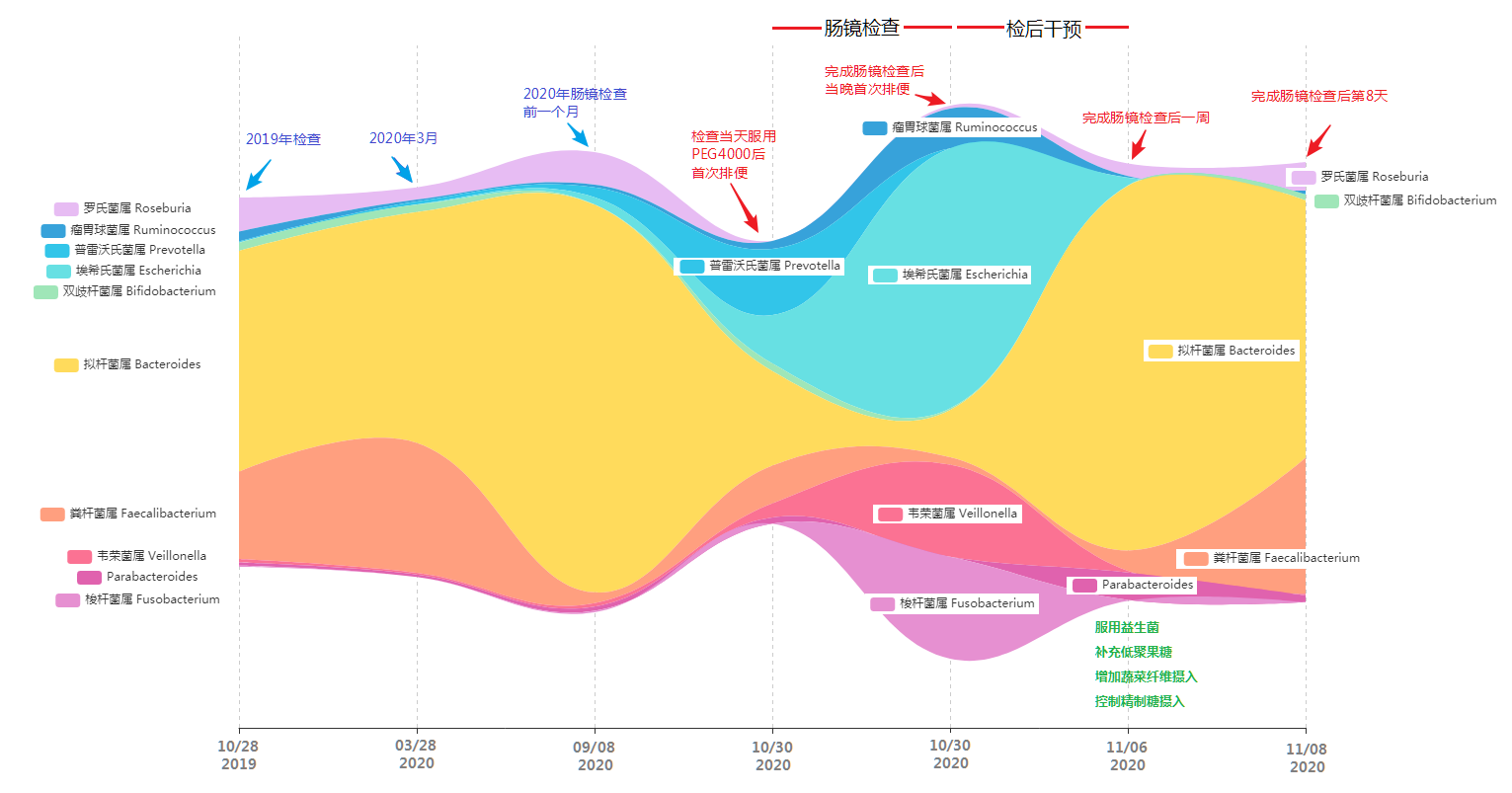
肠镜当天菌群构成和丰度变化

从图中可以看到,原本肠道内占极高比例的黄色拟杆菌属比例大幅压缩,罗氏菌属几乎消失不见。对应的普雷沃氏菌属以及埃希氏菌属(主要是大肠杆菌)比例快速增加,另外除了这些菌外还有大量的原占比很低的其他菌属出现(图中未展示)。
而更大的变化出现在完成肠镜检查后,他发现核心菌属变为埃希氏菌属、韦荣菌属、梭杆菌属和瘤胃球菌属。原来的三大核心菌属占比被压缩到了5%左右。
这样的结果虽然与预想的差不多,小杨不禁感慨道:
“ 看来做一次肠镜,菌群确实受到影响了 ”。
肠镜当天肠道菌群总体状况变化
从肠道菌群总体状况来看,也有非常明显的变化。
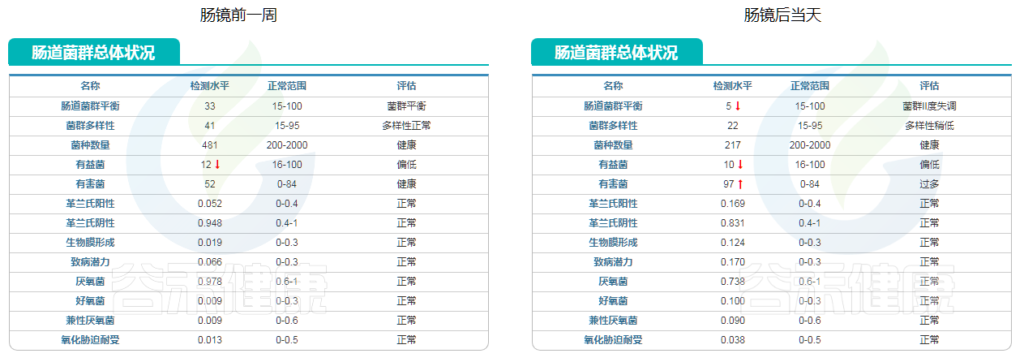
上图显示,肠道菌群平衡这项指标明显下降,菌群多样性也随之下降。紧接着的指标也很有意思,有益菌下降,有害菌大幅上升。说明肠镜检查当天服用聚乙二醇后大量排便扰乱了原先的菌群平衡。
这样的结果再一次印证了他当初的想法。
“ 菌群也许失衡了 ”,小杨开始低落了。
“做完肠镜当天,你吃了什么?”
“肠镜检查后当天及之后的一天,海带、蔬菜…” 小杨仔细回忆了一下,“哦,对了,发现粪便中存在较多未消化的蔬菜和纤维…”
“说明肠道对膳食纤维的消化利用能力尚未恢复”。
“参考肠镜检查后第一次排便的菌群结构” ,我们分析人员指着那张图说到,“其中埃希氏菌属以及韦荣菌属和梭杆菌属都没有代谢复杂碳水化合物的能力…”
“这不就对上了嘛…那之后呢?菌群怎么样了?” 小杨着急地问道。
“别担心,马上给你看一周后的检测结果”
肠镜后一周检测结果
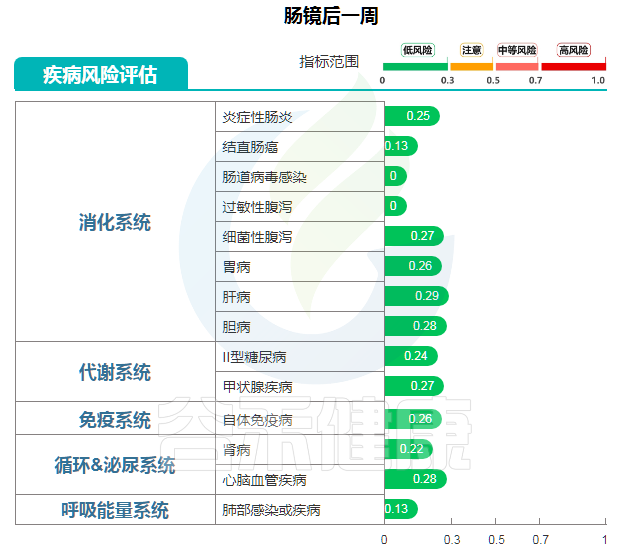
图中可以明显看到,肠镜后一周恢复健康,结直肠癌风险值为0.13,风险很低,也未见肠炎等消化道疾病风险。
“ 一切正常 ”
“肠镜检查我也很正常的,那就是说和肠镜检查结果一致咯…” 这回小杨神色又轻松了不少。
“当然”
“那我能不能知道,我的菌群到底恢复了没有?” 小杨果然没有忘记。
“行吧,既然你这么惦记,再给你整明白点儿…还记得2019年,你在我们这儿测的菌群结果吗?”
“怎么可能忘记,那可是我第一次尝试肠道菌群检测啊” 小杨好像有些得意。
“那这次要不要拿出来一起分析下?”
“可以啊,求之不得,就问你数据还在不?” 说着,小杨笑了起来。
“在啊,我们可是经过你同意的哦” …
“哪儿那么多废话,赶紧吧” 小杨知道这样的机会并不多。
“让我们把时间线再拉长到1年前…”
下图是基于小杨 2019-2020年不同时间点取样的肠道菌群结果变化图(“河流”图),从图中我们可以看到肠镜检查对肠道菌群产生的影响。
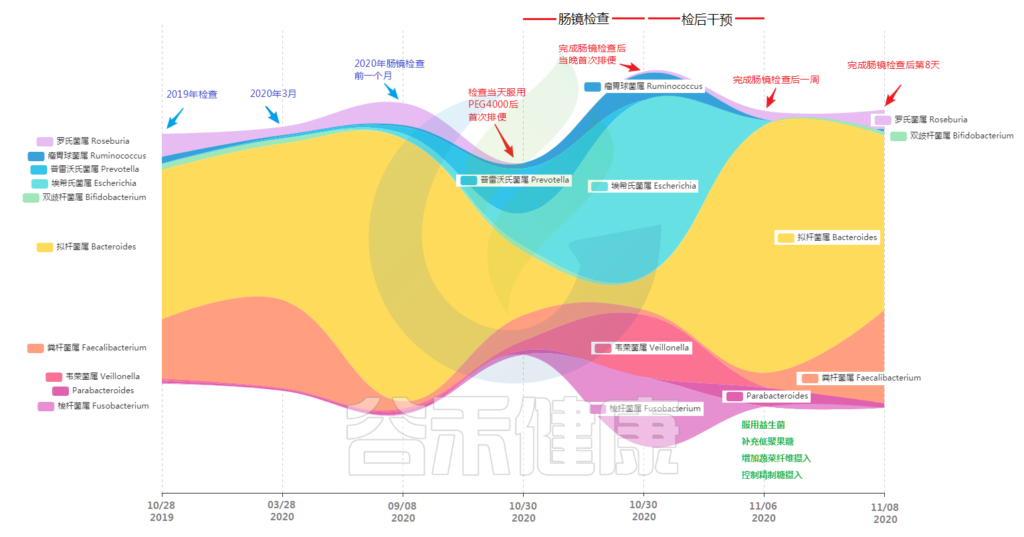
首先图表中的第1/2/3个时间点分别是小杨检查前一年、半年和一个多月前,可以看到其中肠道菌群核心构成的菌属为拟杆菌属。
此外粪杆菌属也就是柔嫩梭菌属Faecalibacterium也是主要菌属,但是期间比例变化较大,从半年前的近30%到了检查前不到5%,另外罗氏菌属也是核心菌属。以上这几个属占据了80%以上的菌群构成,而且存在相对稳定,其他菌属丰度含量较低,均在1%左右。
“我能不能问下,肠镜检查后占比大幅提高的几种菌,是一直存在于肠道内?还是经过清肠准备时引入的?” 小杨又开始好奇了。
“ 经过对之前几次数据的比对,确认这些菌在之前的几个时间点就存在于肠道内,但是比例不高 ”
“那么为什么聚乙二醇服用后在没有引入新的菌的情况下,会改变原来菌群的构成比例?” 小杨的大脑飞速运转着。
“ 这里可能的原因是肠道内不同空间位置的菌分布存在不同。”
“不好意思,我还是不太明白,能给我详细解释下吗?”
“ 固体粪便主体包含了绝大部分的肠道菌群,这些肠道菌群是利用结肠食物残渣的主要菌群,此外肠粘膜部位的黏膜部分也存在大量菌群,这些菌群的构成与粪便主体的菌群有所差异,除了食物残渣之外还可以利用粘液作为食物来源。” 分析人员耐心解释,
“ 梭杆菌属、韦荣菌属以及埃希氏菌属可能在肠粘膜部位含量较丰富,清肠过程后期随着水分排出的黏膜部分菌占据主要比例。”
“哦,我好像又明白了…” 小杨点了点头。
“对了,从图中看来,菌群差不多恢复了” 小杨的眼里闪烁着一丝喜悦。
“你说的没错”
“多亏了你们的饮食建议啊…” 小杨还没来得及长篇大论发表感谢,却又被阻止了。
“另外,健康人的肠道菌群具有恢复健康的能力”。
“能详细解释下吗?”显然又勾起了小杨的好奇心。
“你想啊,吃药,各种不健康的方式都会影响肠道菌群变化没错吧?而你现在却这么健康地站在这儿,某种程度上,肠道菌群有自己的恢复力,当然你可以理解为,整个肠道菌群像一个弹簧一样,有弹性,可受到干扰也可以恢复健康…” 分析人员说着,又甩给小杨一个链接,“可以看看这个文章”
看完后,小杨若有所思,“这么说,肠道菌群恢复力和饮食干预都有功劳,你能告诉我,你们是怎么想到这样的干预方式吗?”
“这…我们其实参考的是抗生素的情况,把你的肠镜检查比作一次抗生素的干扰,这么说明白吗?”
“哦,就是按照抗生素干扰处理呗”,小杨秒懂。
“是的,我们查阅相关文献结合临床经验…” 分析人员开始了讲述。
“ 与抗生素杀死菌群的情况类似,服用泻药将大量菌群排出同样也会导致菌群数量和丰度下降,和抗生素不同的是肠道不同部位的菌可能清除比例存在差异。”
小杨觉得很有道理,频频点头示意。
“ 如果在这种情况下直接摄入大量精制碳水化合物或游离糖,很容易使上述这些菌占据主要菌群,所以在菌群恢复的过程中需要结合自身原有菌群特点进行饮食调节和干预。”
“ 小杨你看,这是2020年一项针对抗生素服用后肠道菌群恢复情况的研究 ” 分析人员拿出早已看过几遍的资料。
研究显示,肠道菌群能否恢复(以是否恢复之前的菌群多样性为标准)包括以下几个阶段:
1、主要恢复菌首先利用黏蛋白多糖在肠粘膜中定植,部分可以通过降解复杂碳水化合物利用能量(如单形拟杆菌)
2、上述产物可以对下游其他菌进行交叉喂养,这部分菌其中一些可以生成短链脂肪酸(如柔嫩梭菌和罗氏菌属的一些菌),进一步为下游其他菌定植提供能量。以上所有菌及代谢物进一步帮助肠粘膜的重建,从而形成正向循环
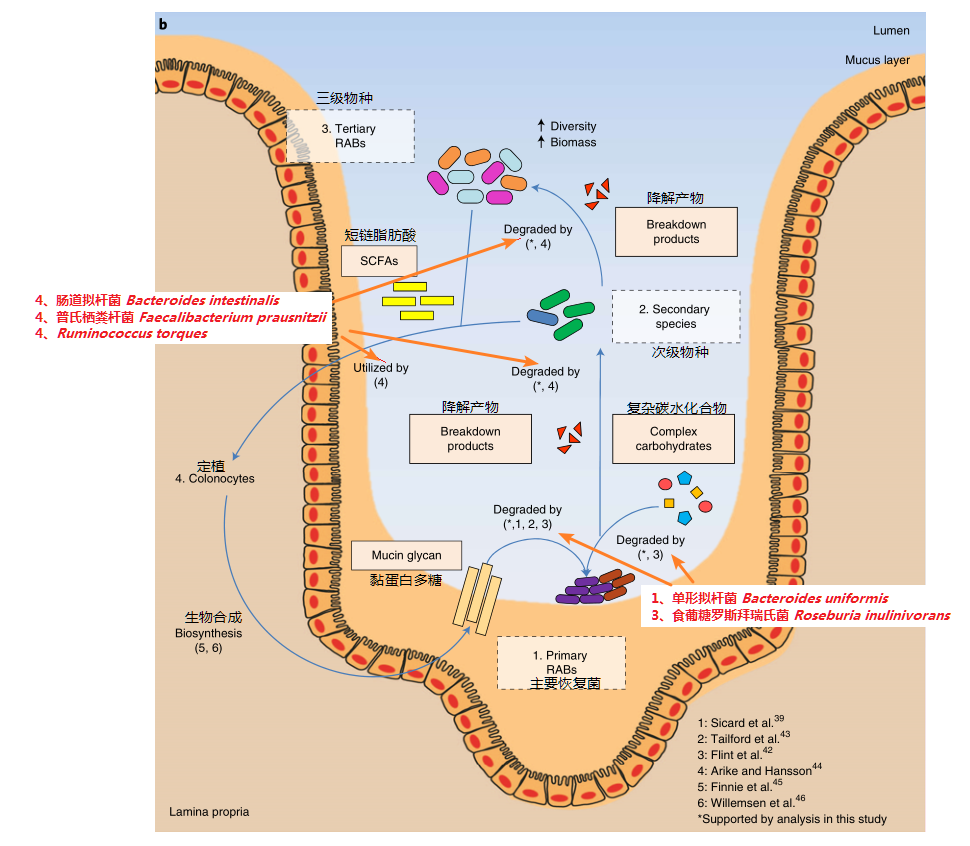
Chng KR et al., Nat Ecol Evol. 2020
从我们这个例子来看,核心菌群中就包括有主要重建菌:单形拟杆菌Bacteroides uniformis、多形拟杆菌Bacteroides thetaiotaomicron、食葡糖罗斯拜瑞氏菌Roseburia inulinivorans以及普氏栖粪杆菌Faecalibacterium prausnitzii 等,属于较易恢复的菌群结构。
“所以说,让我多吃膳食纤维和复杂碳水化合物,另外同时补充益生菌和低聚果糖,控制糖的摄入。其实是为了重建菌群对吧? ” 小杨觉得自己一下懂了好多。
“当然要尽可能地在短时间内,让你的主要核心菌大部分得到恢复。对了,你在第8天有改变了什么饮食习惯吗?”
“增加了低聚果糖的摄入,从每天10g增加到每天20g,怎么了?是有什么变化吗?” 小杨有些不解。
“ 看这里,第8天可以看到明显的粪杆菌属和罗氏菌属的丰度上升… ”
……
这一天,小杨感到格外轻松自在。
关于小杨的故事,到这里,告一段落。
文中描述的人物是真实案例(小杨是化名),前后多次进行谷禾肠道菌群健康检测,以及在医院做的肠镜检查。
我们在征得本人同意的情况下,将此案例发布于此。在这里,我们也要感谢小杨(化名)愿意提供部分报告信息,以便于我们分享时,大家能更好地理解。
最后,分享两个需要注意的点。
#
如果肠道菌群构成中:
单形拟杆菌Bacteroides uniformis、多形拟杆菌Bacteroides thetaiotaomicron、食葡糖罗斯拜瑞氏菌Roseburia inulinivorans以及普氏栖粪杆菌Faecalibacterium prausnitzii这几种菌占比很少,那么在进行如肠镜检查、抗生素或者较严重腹泻之后,需要特别注意肠道菌群的重建。
如果肠型构成中主要以普雷沃氏菌属或瘤胃球菌为主,且普氏栖粪杆菌Faecalibacterium prausnitzii 比例较低,那么需要考虑增加益生元的供给,并注意主食中以抗性淀粉为主,帮助第二级菌群定植,辅助重建。
总之,无论是关于肠镜检查,还是菌群恢复,都希望大家能从中得到一些收获。
谷禾健康
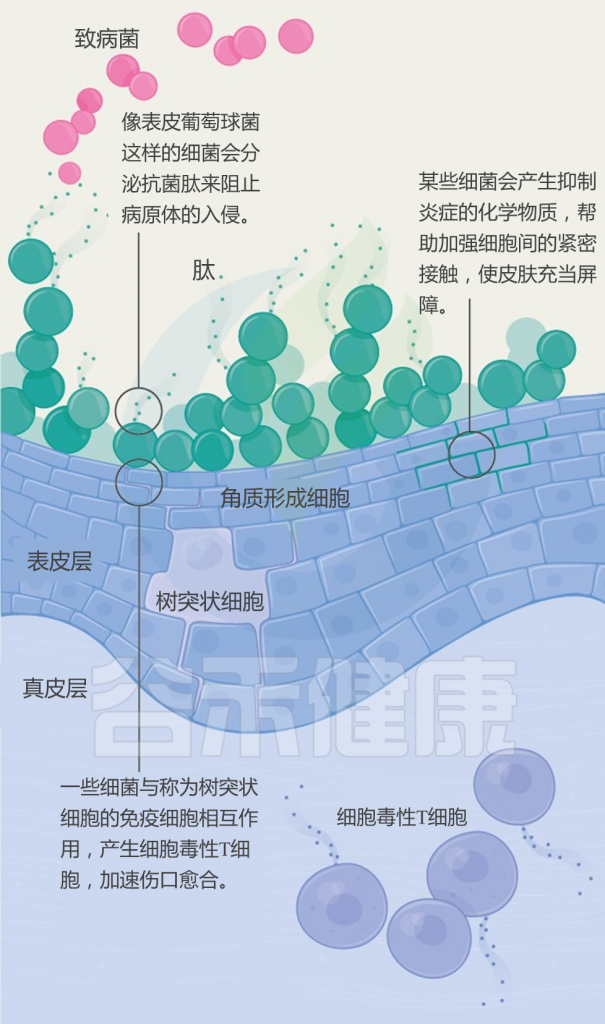
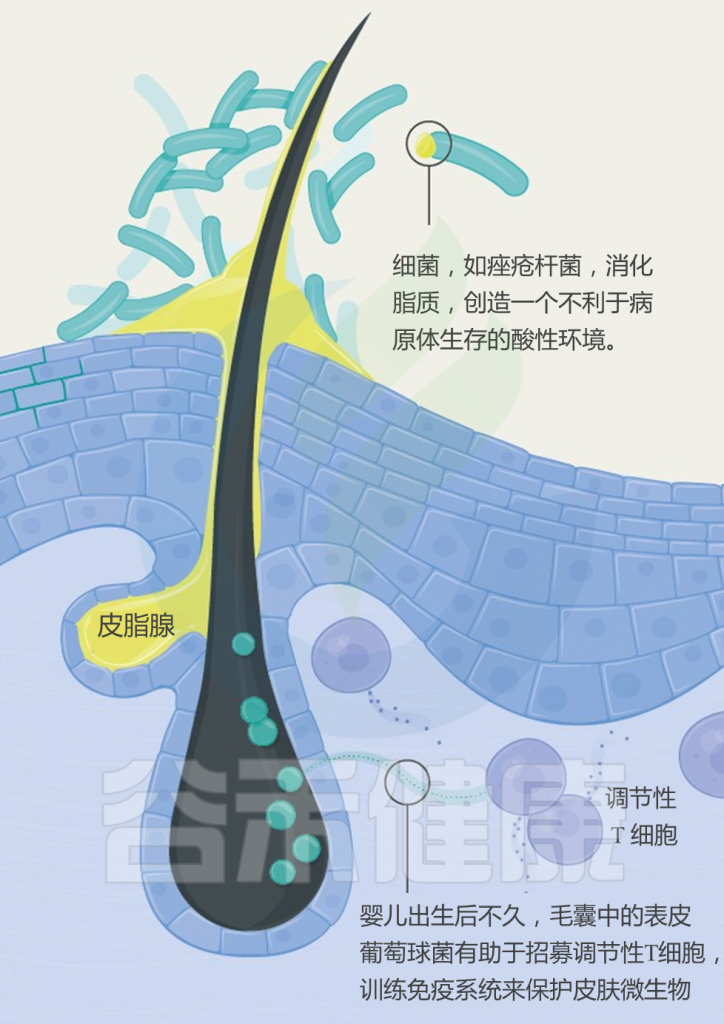
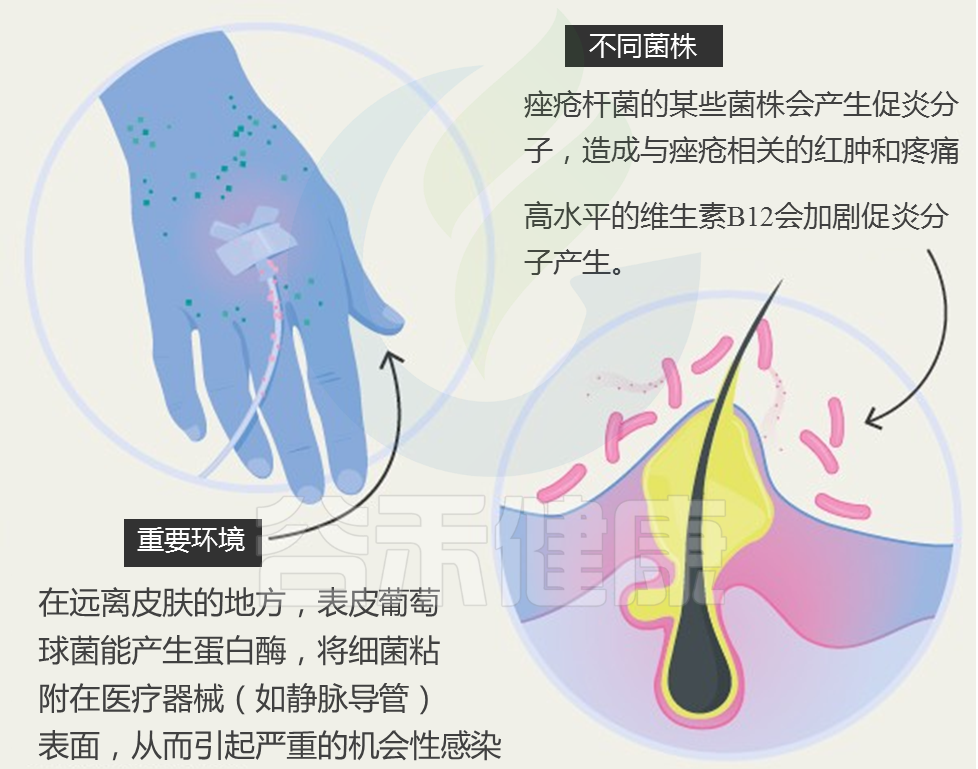
世界卫生组织报告说,银屑病(俗称“牛皮癣”)影响全世界近一亿人。 发病的平均年龄在15至35岁之间。


牛皮癣最常见的形式是斑块状牛皮癣
皮肤细胞异常快速地生长并在皮肤上堆积
多余的细胞会产生厚而有光泽的鳞片
带有干燥,发痒的红色斑块,有时会很痛
关于牛皮癣的常见类型详见附录。
大多数牛皮癣患者会经历症状的“周期”。有时可能会在几天或几周内出现严重的症状,然后症状可能会消失。在几周内,病情可能会再次发作。周而复始。没有症状的时候并不意味着牛皮癣不会复发。
有一点需要说明的是,牛皮癣并不会传染。


牛皮癣的症状是如何产生的?
与其说牛皮癣是皮肤病,倒不如说是一种自身免疫性疾病。自身免疫性疾病是身体自我攻击的结果。当牛皮癣发生时,T细胞会错误地攻击皮肤细胞。
这种错误的攻击会导致皮肤细胞生产过程过度运转。加速的皮肤细胞生成会导致新的皮肤细胞发育过快,然后被推到皮肤表面堆积。
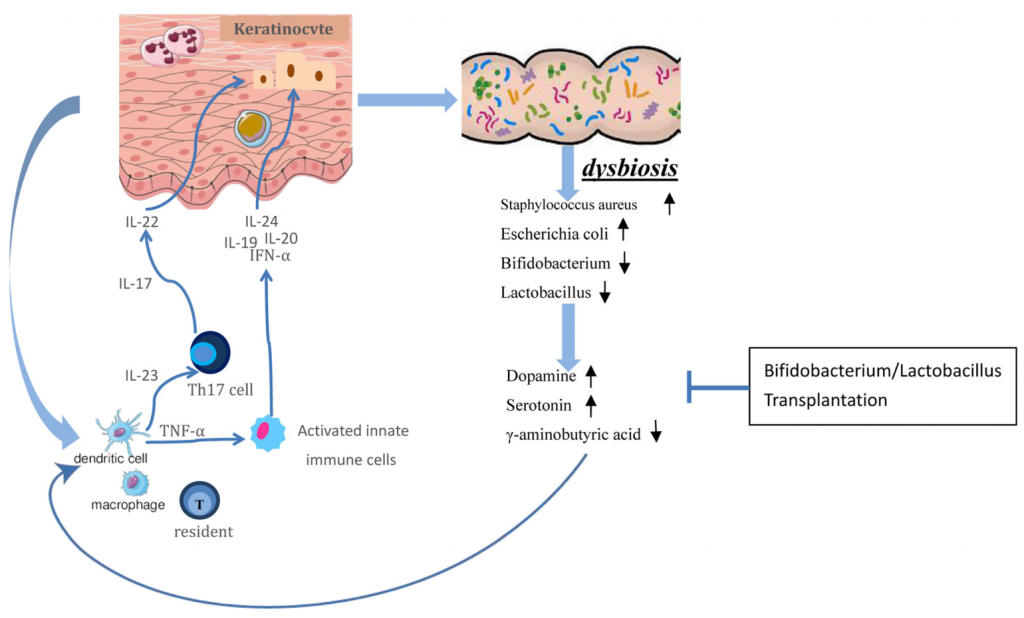
那么,肠道微生物如何影响皮肤?肠-脑-皮肤轴如何发挥作用?肠道神经递质有什么样的调节功能?与牛皮癣之间有什么关系?具体的病理作用是什么?我们来详细了解一下。
牛皮癣患者肠道微生物组成
多项研究采用16S rRNA测序技术,分析了牛皮癣患者和健康人的粪便微生物组。结果显示,牛皮癣患者的肠道菌群结构与健康人不同。
在门水平上,牛皮癣患者的厚壁菌门和放线菌门丰度显著增加;在种水平上,牛皮癣患者的Ruminoccocus gnavus, Dorea formicigenerans,Collinsella aerofaciens丰度显著增加,而Prevotella copri ,Parabacteroides distasonis,Akkermansia muciniphila丰度显着降低。
最近的研究发现,牛皮癣患者的厚壁菌门和拟杆菌门的相对丰度在门的水平上是颠倒的,16种系统型在属水平上存在显著差异。
Hidalgo-Cantabrana 等人在牛皮癣患者中观察到严重的菌群失调,某些菌群多样性较低且相对丰度发生改变。研究表明,牛皮癣改变了患者肠道菌群的特性和功能。
牛皮癣和其他全身性炎症性疾病一样,涉及各种免疫途径的不适当激活,导致促炎性细胞因子升高。而肠道菌群参与促炎性Th17细胞的发育,调节炎症性肠病和肥胖症等疾病中的炎症。
前面提到牛皮癣患者肠道中的艾克曼菌(A. muciniphila)明显减少,该菌是可以增强肠道上皮的完整性并能抵抗全身性炎性疾病。
同样发现艾克曼菌减少的还有Scher等人,他们的研究,发现牛皮癣患者同时还伴有Ruminococcus, Pseudobutyrivibrio的相对丰度降低。而金黄色葡萄球菌(Staphylococcus aureus),白色念珠菌(Candida albicans)和马拉色菌(Malassezia)在皮肤和肠道中定植的增加与牛皮癣加剧有关。所以说,肠道和皮肤之间的联系是强烈的和双向的。
那么,肠道菌群是怎么对皮肤产生影响的呢?
取决于肠道微生物群及其代谢产物从肠道到皮肤的传播。某些致病细菌可以产生某些代谢产物,例如苯酚和对甲酚。 这些代谢物可以破坏皮肤屏障的完整性和表皮分化,减少皮肤的水合作用,并损害角化作用,这些作用与进入血液和在皮肤中积累有关。
短链脂肪酸(SCFA)是一种代谢产物,参与免疫细胞的活化和凋亡。 越来越多动物实验证据表明,慢性系统性炎症是肠道生态失调的主要后果。由于促炎性细胞因子的分泌,上皮通透性增加和激活的效应T细胞之间的不平衡,这导致了慢性全身性炎症的恶性循环。 这可能是肠道微生物组诱导皮肤损伤的众多机制之一。
肠道微生物的畸变导致多种炎症性皮肤病,例如寻常痤疮,脂溢性皮炎和牛皮癣。 越来越多的证据表明肠道菌群失衡会诱发炎症性皮肤病。
当然这也为我们提供了一种改善皮肤状况的可行方法——肠道菌群调节。基于这些发现,益生菌可以治皮肤炎症,方法简单,安全且便宜。(更多关于牛皮癣的预防措施详见后面章节)
关于微生物群和神经系统之间的干扰,研究表明宿主神经递质和/或相关途径在交流过程中起着关键作用。
斯托克斯和皮尔斯伯里首先基于各种实验证据和临床轶事,提出了肠-脑-皮轴概念,表明胃肠道调节介导了情绪和神经状态对皮肤的影响,情绪状态(如抑郁、焦虑等)会促进局部和全身性炎症。
研究还发现,情绪状态与炎症性皮肤状况之间的重要联系可以通过肠道菌群来调节。
最近的研究支持肠-脑-皮肤理论的证据。 我们前面的文章说过,肠道微生物会影响皮肤状况(痤疮的严重程度,点此查看 痘痘?粉刺?皮肤问题很可能是肠道问题),这与它们调节全身性炎症,氧化应激和情绪变化的能力是一致的。
牛皮癣患者肠道微生物通过分泌神经递质介导免疫系统和神经系统之间的相互作用

Guang Chenet al., Dermatol Ther (Heidelb),2020
细菌代谢产物对内源性中枢神经系统信号的激活涉及许多神经递质,包括GABA,5-羟基色胺(5-HT),去甲肾上腺素,多巴胺等。
以下对3种常见的神经递质展开讨论。
多 巴 胺
多巴胺是中枢神经系统中的一种重要神经递质,可控制认知、情绪、免疫和神经内分泌。
1. 在牛皮癣中,免疫系统和神经系统之间的串扰通过神经递质(如多巴胺)通过影响局部组织环境来调节免疫反应,并可能例如影响T细胞的活化状态和迁移。 免疫细胞还利用神经递质相互交流,以牛皮癣的组织特异性炎症为目标。
2. 证据表明,在31–88%的病例中,患者报告称压力是其牛皮癣的诱因。压力也是牛皮癣暴发的结果。 因此,所有的压力激素,包括多巴胺,都应该是牛皮癣的重要调节剂。
3. 多巴胺可以增加角质形成细胞的活性,这在细胞因子和趋化因子的释放中起作用。
5- 羟 色 胺
5-羟色胺是一种信号分子,通过旁分泌,内分泌和邻分泌调节多种生理过程,包括免疫系统,胃肠道和神经系统。超过90%5-HT是由胃肠道肠嗜铬细胞(EC)合成的,而5%的5-HT是在肠系膜神经元中合成的,还有一部分是在大脑中合成的。
血清素水平可以通过色氨酸羟化酶的帮助,通过分泌小分子(如SCFAs)来促进ECs产生血清素。
80年代,5-HT被定义为一种免疫调节剂,因为它具有抑制炎症的能力,包括牛皮癣,5-HT改变了血清素能系统失调患者的免疫反应。
GABA
GABA作为CNS的主要抑制性神经递质,具有广泛的生物学活性,包括调节突触传递,促进神经元发育和放松,预防失眠和抑郁。
越来越多的证据表明,GABA可以被细菌消耗和/或产生。乳酸菌和双歧杆菌是GABA的主要来源。
最近的一项人类研究中,从瘦到肥胖个体的粪便微生物组移植提高了血浆中的GABA水平,这表明微生物组可以控制GABA的产生水平。
GABA 的免疫作用
研究人员发现了GABA的各种生物活性,例如抗高血压,抗糖尿病,抗癌,抗氧化剂,抗发炎,抗微生物和抗过敏作用。
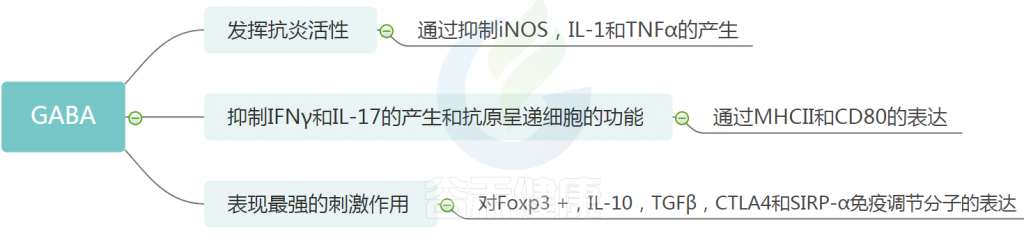

GABA与牛皮癣
牛皮癣患者通常有瘙痒,可能与GABA系统有关。 实验研究报道了牛皮癣患者皮肤中GABA配体和GABAA受体的表达增加,表明它们在与牛皮癣相关的发病和瘙痒中具有关键作用。 因此,GABA可以被认为是预防和治疗牛皮癣的潜在替代疗法。
表 神经递质产生菌(代表性的菌)
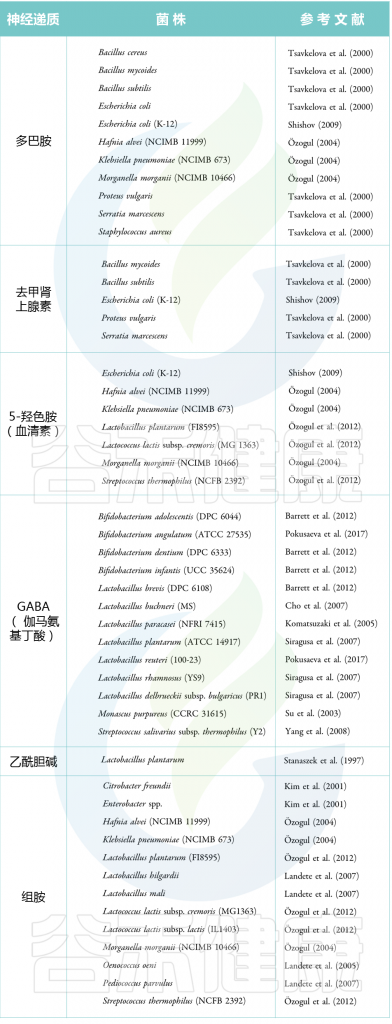

Guang Chenet al., Dermatol Ther (Heidelb),2020
综上,肠道菌群可能通过分泌神经递质来介导神经系统和免疫系统之间的串扰。FMT(粪菌移植)可能是牛皮癣的一种新型疗法。 需要进一步的临床试验以提供可靠的证据,验证潜在机制。
肠-脑-皮轴是神经传递调节中可能的通讯途径。通过了解这些途径及其与疾病特征的关系,可以设计肠道菌群介导的干预措施来操纵这些靶点以治疗牛皮癣。
虽然牛皮癣的症状可能反反复复,但至少可以做一些相应的预防措施,避免不断诱发其发作。
1. 减 轻 压 力
生活中的日常压力可能会对任何人产生负面影响,但对牛皮癣患者而言尤其成问题。身体容易对压力产生炎症反应。这种反应会引起牛皮癣发作。
——心理咨询
牛皮癣患者更容易出现抑郁等心理问题。当出现新斑点时,可能会不太自信。这种情况的持续循环会造成心理伤害。可以咨询专业的心理专家或加入牛皮癣患者正规论坛等进行交流。
——瑜珈
瑜伽促进身心放松,帮助缓解压力。
——冥想和其他放松技术。
尝试读书,跑步,与家人朋友聊天等。
2. 避 免 某 些 yao 物
某些yao物可能会干扰人体的自身免疫反应并引起炎症,从而引发牛皮癣。
3. 防 止 皮 肤 受 伤
有些人的皮肤损伤会进一步引发牛皮癣。这就是所谓的Koebner现象。引发牛皮癣的常见皮肤损伤包括晒伤和擦伤。
保护好皮肤可以帮助预防牛皮癣。如果外出时,应始终采取额外的预防措施,可以包括以下措施:
–外出时使用防晒霜并戴上帽子
–进行园艺时戴长袖手套
–在户外活动前喷杀虫剂
–参加户外活动时要谨慎
对于牛皮癣患者来说,过多的阳光会导致症状加剧。虽然说,适量的阳光可以缓解某些症状,保持10分钟左右,晒伤会引起耀斑。
4. 温 和 清 洁
牛皮癣患者应选择温和的保湿清洁剂,避免使用会导致皮肤干燥或刺激皮肤的配方。
避免含以下刺激成分:
月桂醇、肉豆蔻醇、棕榈醇、鲸蜡醇、二十二醇、视黄酸等。
5. 注 意 保 湿
寒冷干燥的气候也会加剧牛皮癣的症状。在这种天气下,皮肤水分较少,尤其热空调会更加干燥,实在太干的情况,最好配备加湿器。
洗澡不要超过10分钟,洗完澡后尽快保湿。可用毛巾拍干皮肤并涂上无气味的乳液,帮助锁住水分。
6. 避 免 感 染
感染会引发牛皮癣,因为感染会对免疫系统造成压力,从而引起炎症反应。
特别是链球菌性咽喉炎与小儿牛皮癣的发作有关。牛皮癣突然发作可能发生在耳痛,扁桃体炎或呼吸道或皮肤感染后。
如果发现有皮肤受伤(例如割伤或伤口),请确保正确清洁包扎以防感染。其他预防感染的方法包括:
–全天经常洗手
–避免与他人共享食物,饮料或饮食用具
–限制接触生病的人,尤其是儿童
7. 饮 食 健 康
如果是肥胖或超重患者,牛皮癣症状可能加重。因此,需要注意通过锻炼和饮食健康的饮食来控制体重。2013年发表在JAMA Dermatology上的一项研究发现,低热量饮食有利于改善牛皮癣。
另外,某些食物可能会引起人体内的炎症,从而引发牛皮癣症状。当然也有食物可以帮助减轻炎症和爆发现象。
可能引起炎症的食物包括:
红肉、乳制品、高脂食品、加工食品、精制糖、茄属植物、柑橘水果等。
可能减轻炎症的食物包括:
肥鱼,例如鲑鱼和金枪鱼
种子,例如亚麻籽和南瓜籽
坚果,尤其是核桃和杏仁
绿叶蔬菜,例如菠菜和羽衣甘蓝
关于抗炎饮食,促炎饮食可查看我们之前的文章:
8. 避 免 酗 酒
酒精是许多牛皮癣患者的诱因。哈佛大学医学院附属医院的一项研究发现,每周喝两到三杯酒(不是淡啤酒)和牛皮癣的患病率增加有关。
9. 避 免 吸 烟
如果患有牛皮癣,请避免吸烟。烟草会增加牛皮癣的风险,加剧牛皮癣症状。
10. 保 证 睡 眠
建议成年人每天睡七至八个小时。充足的睡眠可以支持免疫系统,并可以帮你保持健康的体重和控制压力。所有这些对于预防牛皮癣都很重要。
附录:
牛皮癣的常见类型
斑块状银屑病——常见类型
导致红色发炎的斑块,覆盖皮肤区域。这些斑块通常覆盖着发白的银鳞或斑块。
肠状牛皮癣——儿童时期很常见
引起小的粉红色斑点。常见的部位包括躯干,手臂和腿。很少像斑块状牛皮癣那样浓密或凸起。
脓疱性牛皮癣——多见于成人
导致白色的脓性水疱和大面积的红色发炎皮肤。
反向银屑病——特殊部位
导致皮肤发红,发亮,发炎。在腋下或乳房等部位皮肤皱褶周围发展。
红皮病性牛皮癣——严重且非常罕见
通常一次覆盖身体的大部分,鳞片经常脱落成大块或片状脱落。患有这种牛皮癣的人可能经常发烧或生病。
相关阅读:
主 要 参 考 资 料:
Mayo Clinic Staff. (2017). Psoriasis. mayoclinic.org/diseases-conditions/psoriasis/symptoms-causes/syc-20355840
Chen Guang,Chen Zai-Ming,Fan Xiao-Yan et al. Gut-Brain-Skin Axis in Psoriasis: A Review.[J] .Dermatol Ther (Heidelb), 2020.
Kurd SK, et al. (2010) The risk of depression, anxiety and suicidality in patients with psoriasis: A population-based cohort study. DOI:10.1001/archdermatol.2010.186
Rachakonda TD, Schupp CW, Armstrong AW. Psoriasis prevalence among adults in the United States. J Am Acad Dermatol. 2014 Mar;70(3):512-6. doi: 10.1016/j.jaad.2013.11.013. Epub 2014 Jan 2. PMID: 24388724.
Psoriasis. (n.d.).aad.org/public/diseases/scaly-skin/psoriasis
Women who drink beer more likely to develop psoriasis.(2010).sciencedaily.com/releases/2010/08/100816162645.htm
Mayo Clinic Staff. (2018). Psoriasis. mayoclinic.org/diseases-conditions/psoriasis/symptoms-causes/syc-20355840

谷禾健康

我们之前几篇文章都讲过关于抑郁,焦虑和肠道菌群的关系。
最新研究速递 | 柳叶刀:肠道微生物群在神经系统疾病中的作用
不仅如此,还和微量元素、维生素缺乏有关。
1. 微 量 元 素 篇
锌是帮助调节许多生物过程(包括我们的情绪)的核心营养素之一。锌的饮食来源多种多样,尤其是在红肉,家禽和鱼类中。当人的锌水平低时,可能会出现焦虑,抑郁,失眠和情绪失调等状况。
Petrilli 等人研究发现抑郁症和精神病患者的锌含量低。在患有帕金森氏病,阿尔茨海默氏病,唐氏综合症和多动症的患者中也发现了锌缺乏。
对于缺锌的患者,补锌已被证明有助于改善抑郁症状,是 “自然的抗抑郁药”。


一项研究表明,与仅接受SSRI治疗的患者相比,补充锌和SSRI可以更有效地改善重度抑郁症。对实验动物进行锌的长期治疗具有与抗抑郁药相同的作用。
注:SSRI是一类新型的抗抑郁药
当然,在评估锌水平和补充需求时,还需要考虑许多其他因素(例如炎症,其他微量元素的吸收等)。
碘缺乏,特别是在早年间缺乏,会阻碍身体和大脑的发育。碘是甲状腺激素必不可少的组成部分,这对大脑的发育和维持至关重要。碘缺乏症困扰着非常多人,但实际上因碘缺乏引起的智力障碍是可预防的。
大多数植物性食品的碘含量都非常低。素食主义者通常相对容易缺碘。
大脑非常需要铁。铁是产生神经递质(血清素,多巴胺和去甲肾上腺素),脑能量生成,海马功能(记忆),细胞信号传导和婴儿大脑发育所必需的。
许多植物性食物中的铁含量低于动物性食物,且更难吸收。素食者血液中铁的含量与杂食者差不多,但是他们体内的总铁存储量(储备量)较低。


2. 维 生 素 篇
维生素B在调节我们的心理健康方面也起着重要作用。
“B族维生素” 是指有助于人体细胞正常运作的八种必需营养素。B族维生素们既可以一起协同工作,又可以发挥各自的独特功能。


B维生素的八种类型包括:
B1,B2,B3,B5,B6,B7,B9,B12
某些与心理健康有关的疾病也可能与维生素B缺乏症并存。例如,早期研究表明,自闭症儿童的B6偏低,补充B6后,有些儿童表现出明显的改善,并且是第一次开始说话。
维生素B1,B3,B6
已成功用于治疗许多患有焦虑症和其他心理疾病的患者。在小鼠研究中,实际上发现VB3与苯二氮卓类和巴比妥类具有共同的特性。
维生素B12
有抑郁症患者存在B12缺乏症,而B12缺乏症的人比非缺乏症的人更容易患严重抑郁症。B12缺乏的其他症状可包括疲劳,嗜睡,躁狂等。
素食主义者饮食中几乎不含维生素B12,长期严重缺乏B12会致命。素食主义者可以服用补品或食用强化酵母(未经强化的天然酵母不包含维生素B12)。


维生素B12缺乏很普遍。一些研究发现,多达86%的成年人(无论选择哪种饮食)都缺乏。研究人员报告说,其价值范围很广,总的来说,素食主义者的B12含量往往较低。
B类维生素包括B2,B6,B9,B12也已被证明有助于治疗精神分裂症的症状。
维生素D——“阳光”维生素,对我们的心理健康也至关重要。大约50-90%的维生素D是由阳光照射产生的,其余的来自饮食。
维生素D缺乏症的人数众多,一些专家认为维生素D缺乏症是“全球健康问题”。
目前还不清楚维生素D与精神障碍之间的确切关系,但发现存在维生素D缺乏症风险的人群患抑郁症的风险增加,而维生素D含量较低可能会导致或加剧抑郁症状。
神经退行性疾病,肥胖,糖尿病,高血压,纤维肌痛,慢性疲劳综合症和骨质疏松症也与维生素D缺乏症有关。
关于神经退行性疾病与菌群详见文章 最新研究进展 | 行为和神经退行性疾病中的肠道微生物分子
维生素D3


维生素D3是我们身体需要的维生素D形式。在大脑的生长和发育中很重要,调节大脑中的钙水平,帮助保护脑细胞免受有害的氧化作用,并支持海马体(大脑记忆中心)的健康。
大多数研究发现,纯素食者的血液中维生素D3含量较低,并且在冬季,素食者更有可能出现维生素D3不足。
维生素K1在许多植物性食品中都丰富,但是维生素K2同样重要,而且常常被忽略。


维生素K2有多种形式,但我们需要的基本形式称为MK-4。在大脑中,需要MK-4来构建鞘脂的关键细胞膜成分,以及维持脑细胞的整体健康和功能。
维生素K2的MK-4形式仅存在于动物性食品中。
那么,是不是这些微量营养素补充越多越好呢?
国际自闭症研究会议上,约翰·霍普金斯大学研究人员提出:怀孕期间叶酸和维生素B12含量过高的女性,孩子患自闭症风险较高。
我们看看研究人员是如何得出这样的结论:
样本量及来源
研究人员分析了1391名母亲及其子女的数据。在孩子出生时招募志愿者,然后追踪长达15年。
检测指标及相关调查
在分娩的头几天检查母亲的血液中叶酸和维生素B12的水平。所有母亲均在怀孕期间进行了有关产前维生素和其他补充剂使用情况的调查。
发 现
在1391名儿童中有107名被诊断出患有自闭症谱系障碍,但这种自闭症的风险并不是在所有母婴中平均分布的。
· 每周服用三至五次产前复合维生素的母亲,孩子患自闭症的可能性较小。
·在头三个月服用这些补充剂的母亲生自闭症孩子的可能性要低67%
·在中晚期服用,其孩子自闭症可能性要低62%
·在妊娠中期服用,孩子患病的可能性要低57%
至此,服用产前维生素听起来很不错吧,问题来了:
在1391名母亲中,有95名母亲的VB12血液水平被世界卫生组织认为过高。在这些母亲中,有15个孩子被诊断出患有自闭症谱系障碍,这一风险是其他母亲的三倍。
同样,140名母亲的血液中叶酸水平“过高”,其中16名儿童患有自闭症谱系障碍。
而这其中有21位母亲两种维生素都“过量”。
更夸张的是,这21位母亲,有10个孩子被诊断出患有自闭症。换句话说,维生素水平过高的人中有将近一半有自闭症儿童。


当然,此研究也存在缺陷。
缺点之一是血样是在孩子出生后立即采集的,而不是在怀孕期间采集的。
此外也有猜测,是否因为她们知道正在接受研究而服用较大剂量的维生素?这些不得而知。
综上,如果您或者您家人现在正在孕育新的生命,需要考虑维生素服用的剂量以及评估相应风险。
换句话说,可以服用,但不宜过量。
相关阅读:
主要参考文献:
Naeem Z. (2010). Vitamin d deficiency- an ignored epidemic. International journal of health sciences, 4(1), V–VI.
Penckofer, S., Kouba, J., Byrn, M., & Estwing Ferrans, C. (2010). Vitamin D and depression: where is all the sunshine?. Issues in mental health nursing, 31(6), 385–393. doi:10.3109/01612840903437657
Ranjbar, E., Kasaei, M. S., Mohammad-Shirazi, et al., (2013). Effects of zinc supplementation in patients with major depression: a randomized clinical trial. Iranian journal of psychiatry, 8(2), 73–79.
Petrilli, M. A., Kranz, T. M., Kleinhaus, K., Joe, P., Getz, , et al., (2017). The Emerging Role for Zinc in Depression and Psychosis. Frontiers in pharmacology, 8, 414. doi:10.3389/fphar.2017.00414
Grabrucker A. M., Rowan M., Garner C. C. (2011). Brain-delivery of zinc-ions as potential treatment for neurological diseases: mini review. Drug Deliv. Lett. 1, 13–23.
免责声明:本公众号内容仅作交流参考,不作为诊断及医疗依据,身体问题请及时就医。
图片来自网络,侵删