 国家高新企业 | ISO9001认证
国家高新企业 | ISO9001认证 二级病原微生物安全实验室
二级病原微生物安全实验室- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
国家高新企业 | ISO9001认证 二级病原微生物安全实验室

谷禾健康
中风是全球范围内最具破坏性的最严重的脑部疾病之一,治疗选择非常有限。中风和其他神经系统疾病与微生物群破坏有关。反过来,中风诱发的菌群失调会通过调节免疫反应而对疾病结果产生重大影响。
了解肠细菌-脑串扰的性质可能会导致基于微生物组的治疗方法,从而可以改善患者的康复。
本文主要介绍关于肠道微生物组在中风中的作用的最新知识,以及肠-免疫-脑串扰的潜在机制。
肠道微生物群和周围器官之间的信号传导是由微生物组相关分子模式(MAMP)和微生物组分泌的代谢产物介导的,这些代谢产物可以与粘膜上皮和肠道免疫细胞相互作用,刺激迷走神经或到达全身循环以向大脑发出信号和可能调节神经元和免疫应答。
反过来,支配肠壁的副交感神经和交感神经纤维会从大脑传递输入信号,从而影响肠蠕动,免疫细胞活性,并能诱导肠成分改变。
01
中风改变了微生物群的组成
我们知道,肠道微生物群稳态的破坏,就会容易患病。
实验和临床研究中越来越多的证据表明中风会改变肠道菌群的组成。对中风患者肠道菌群的分析显示,与健康个体相比,微生物组成发生了变化,而菌群失调程度与病变的严重程度相关。
同样,观察小鼠的粪便微生物丰度的变化,发现严重中风要比轻微中风更为明显。但是,在分析菌群多样性和特定菌群时,在临床和实验研究中有些不同结果。
为什么会出现不同结果?
Yin 等人发现与无症状对照组相比,中风患者粪便样本的多样性增加,Stanley等人发现假手术组和中风小鼠之间的微生物群多样性没有变化。相反,其他研究人员则在实验和临床环境中观察到某些菌减少和其他细菌的过度繁殖导致多样性降低,表明多样性程度不一定能预测中风的严重程度。
不同的研究发现,沙门氏菌总体上减少了,同时伴有拟杆菌的过度生长。然而,当在较低的生物分类等级上分析微生物群变化时,在临床和实验性中风中特定细菌变化方面缺乏一致性。
可能的混淆因素
粪便采样位置的差异(在小鼠的胃肠道和患者的粪便中),用于基因组DNA提取和16S rRNA基因分析的方法,中风模型的严重程度以及基线的差异微生物群组成(人类的小鼠起源或个体间和个体内变异)可能是造成矛盾结果的混淆因素。
影响肠道菌群组成的另一个混杂因素是饮食类型。
严重的中风病例总是住院治疗,他们将接受药物治疗并接受医院控制的饮食,而实验性中风后对小鼠的干预很少,这将以啮齿动物和人类的不同方式影响微生物群的组成。
总体而言,临床和实验数据表明中风后微生物群组成发生了变化。然而,迄今为止,尚不清楚哪种类型的肠道细菌参与中风的病理生物学机制。
年龄对微生物群状态的重要性
衰老会改变肠道菌群的组成,并与炎症和肠道通透性增加有关。中风后,与年轻小鼠相比,有高龄微生物群的小鼠具有更高的死亡率,运动功能受损和促炎性细胞因子反应。
粪便微生物群移植(FMT)可以在年轻和老年小鼠中恢复中风的结果。这些实验强调了年龄对微生物群状态的重要性及其对疾病发展的影响,以及微生物群的操纵如何能够导致非侵入性疗法以预防和改善中风预后。
虽然实验结果表明与年龄、肠道生物失调和中风结果相关,但其他风险因素(糖尿病、高血胆固醇水平、饮酒、高脂肪饮食、缺乏锻炼)和性别差异对肠道微生物群和中风结果的影响仍有待实验和临床研究解决。
脑损伤改变微生物组组成的机制
由于中风后自主神经系统(ANS)的失调而降低肠道运动性和粘膜屏障的完整性。脑损伤后,肠道内胆碱能活性下降,肾上腺素能信号传导增加,这与肠屏障的破坏有关。
因此,使用β受体阻滞剂抑制β肾上腺素能活性可显着恢复中风诱导的肠道通透性,并减少细菌向周围器官的扩散。
在这方面,Singh等人证明了回肠瘫痪本身就可以驱动微生物群变化,这是一种中枢神经系统介导的生态失调。
肠道细菌可以感觉到来自胃肠道的激素和神经递质,并且在体外培养中,在应激激素去甲肾上腺素(NE)存在下,某些条件致病菌能够增殖更多。
有趣的是,在实验性中风后,盲肠中的去甲肾上腺素释放增加,这可能是某些病原体或共生菌过度生长的原因。
另外,ANS调节肠粘液分泌,大脑对肠信号的扰动可能对肠粘液层的大小和质量产生重要影响。
Houlden等人观察到,中风后盲肠内,粘液蛋白产生的失调和产粘液的杯状细胞降低。中风后粘液层的形成受损可能使共生膜与上皮直接接触,转移并引发免疫反应,以及中风后感染。此外,还需要更多关于细菌移位导致实验性中风感染的直接影响的数据。
脑损伤触发肠道中胆碱能和肾上腺素能信号的失衡,增加释放的NE的浓度,降低肠道运动性,并改变与微生物群失调有关的肠脑屏障。所有这些事件都将影响肠道微生物的功能输出,并可能进一步扰乱肠道的免疫稳态。
02
中风的肠-免疫-脑轴
迄今为止,使用动物模型进行的研究提供了最有力的证据,并有助于阐明中风后肠-免疫-脑相互作用的潜在机制。
T淋巴细胞的关键作用
已知T淋巴细胞在脑损伤后继发的组织损伤中起关键作用。促炎性T辅助1细胞(Th1),T辅助17细胞(Th17)和γδT-IL-17+细胞与中风后神经毒性有关,而调节性T细胞(Treg)发挥作用抗炎和神经保护特性。
越来越多的证据,支持微生物群通过调节胃肠道中促炎Th17细胞与抗炎Treg细胞之间的平衡来维持免疫稳态。
通过诱导促炎Th1和Th17介导的反应,具有中风后遗症粪便含量的无菌(GF)小鼠的定殖导致中风结果的恶化,这表明中风诱导的微生物失调触发促炎性免疫反应,从而加剧脑损伤。
这组作者证明,来自派尔氏淋巴集结(Peyer’s patches)的T细胞会侵袭梗塞周围组织,并在中风的急性期产生免疫反应。
肠道菌群的免疫调节作用对于中风恢复至关重要
此外,通过健康捐献者的FMT恢复肠道微生物群的稳态,减少了损伤大小并增加了缺血性脑中Treg细胞的数量。
然而,单独存在的微生物群具有神经保护作用,因为与定殖小鼠相比,完全根除GF小鼠的肠道细菌导致更大的梗塞体积。
同样,这种微生物群介导的对中风的有益作用取决于淋巴细胞反应,表明肠道细菌的免疫调节作用对于中风恢复至关重要。通过显示在缺血性损伤之前使用抗生素对微生物群组成进行控制可减少梗塞体积并改善感觉运动功能,从而证明了肠-脑双向沟通在中风中的关键参与。
在这项研究中,他们表明神经保护作用是由微生物依赖性的肠道树突状细胞(DC)引发的,介导小肠Treg的扩增,通过抑制促炎性γδT-IL-17+细胞发挥作用。
此外,他们还认为IL-10是Treg细胞介导的γδT细胞增殖抑制的主要协调器,因为IL-10-/- 小鼠不受保护作用的影响。由于较少的促炎IL-17 +γδT细胞积聚在脑膜中,因此肠道中的这种抗炎环境会传导到大脑,这与较小的梗死面积有关。
有趣的是,肠道T细胞从肠道迁移到脑膜和脑实质中,表明中风有直接的肠脑沟通途径。
另外,肠道微生物群组成的修饰可通过致耐受性DC使肠道免疫应答偏向抗炎环境。
这些研究表明中风后微生物群受到影响,中风后细菌数量的变化会促进促炎性T细胞反应,肠道免疫细胞迁移至脑膜,这可能会参与继发性脑损伤并恶化中风结果。
也突显了通过肠道淋巴细胞从肠道运输到大脑的过程,沿着肠道-脑轴的直接联系,在那里它们调节了对中风的神经炎症反应。
当然,对于肠免疫细胞是否直接导致脑损伤或修复机制还不是很清楚。重要的是,这种肠道细菌-DC之间相互作用的分子途径仍然难以捉摸。
图 肠道代谢物——中风中的免疫系统干扰
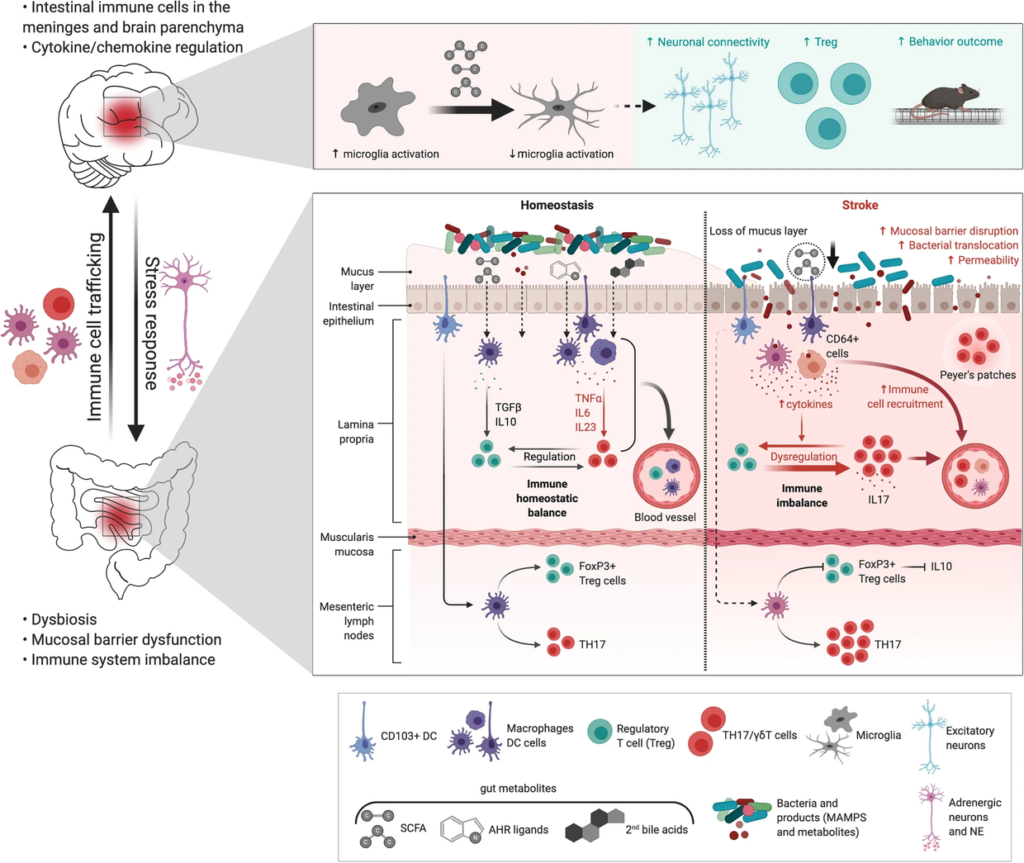
Delgado.et al., Neuromolecular Med, 2020.
中风后肠道微生物对免疫系统的调节。
下图:在肠道中,中风引起生物失调、粘膜屏障功能障碍、肠道通透性增加、细菌移位、中风后感染和通过树突细胞的促炎性T细胞反应(DC)。中风后,免疫细胞,特别是T细胞、CD64 +巨噬细胞和树突状细胞从肠道迁移到脑膜和大脑。肠道代谢物(AHR,次生胆汁酸,MAMPs)作为肠道免疫细胞免疫调节剂的作用尚未明确。中风后微生物来源的SCFA减少,可能引发γδT-IL-17 +细胞和调节性T细胞(Treg)的失衡。
上图:补充SCFA调节神经元活动和突触密度,并且与小胶质细胞激活减少和Tregs增加以及中风后更好的恢复相关。
03
中风肠道菌群的操纵
Benakis等人证明了在诱导缺血性损伤之前用氨苄西林或万古霉素治疗小鼠具有神经保护作用,而使用新霉素则不能保护小鼠免于中风。这种神经保护作用与微生物组组成中的抗生素特异性变化有关,变形菌和厚壁菌门增多,而拟杆菌属的减少。
此外,当他们分析与有益菌群变化相关的预测酶途径时,他们发现异种/芳香族化合物代谢可预测缺血性病变的大小。
微生物代谢产物是细菌-宿主串扰的主要沟通渠道之一
与中风的肠道免疫脑轴有关的一个可能的缺失环节可能是仅由细菌产生的代谢产物。
已知必需芳香族氨基酸色氨酸的代谢产物可通过在DC和T细胞中表达的芳烃受体(AHR)调节免疫细胞功能。一些菌群能够将肠道中的色氨酸分解为AHR的配体。
在小鼠中,微生物群衍生的吲哚激活AHR通过诱导免疫细胞中IL-22促进肠道稳态。在星形胶质细胞的AHR通过激活在多发性硬化的小鼠模型中色氨酸代谢物的神经保护作用的驱动器。
然而,保护作用是配体依赖,其他色氨酸代谢产物已报告加重病程。有趣的是,负责色氨酸分解代谢的内源性犬尿氨酸途径的活性被上调下列缺血性中风和已与炎性反应和恶化的结果。因此,AHR受体的药理和遗传阻断改善了中风恢复。
这些发现暗示了色氨酸衍生化合物在中风中的关键作用。尚不清楚微生物源性色氨酸代谢产物在缺血性损伤中的免疫调节作用。
短链脂肪酸(SCFA)是细菌发酵的生物产物,具有免疫调节作用:它们能够根据免疫情况直接诱导T细胞分化为效应细胞和调节细胞。
小鼠中风后血浆SCFA的水平显着降低,这与恶化的结果有关。
中风之前在小鼠中补充SCFA改善了行为恢复,改善了皮质网络的连通性,并改变了突触可塑性的组织学标志,这与改善中风的长期中风有关。这些作用与小胶质细胞形态向稳态状态的改变和侵袭性淋巴细胞的减少有关。
菊粉改善中风后神经功能缺损
在另一项研究中,他们观察到,与年幼小鼠相比,口服菊粉(产生SCFA的细菌和制造SCFA的细菌基质),可减少老年小鼠中风后的神经功能缺损和改善抑郁样行为。
此外,这些变化伴随着缺血性脑中IL-17 +γδT细胞百分比的降低,但SCFA是否直接影响T细胞极化和迁移尚未解决。出人意料的是,在两项研究中,梗死面积均不受SCFA干预的影响。
所有这些证据表明,微生物代谢产物SCFA在中风后恢复中起重要作用,并且可能参与中风后肠道菌群发挥的免疫调节作用。
04
结 语
本文总结了微生物群组成如何通过调节免疫反应影响中风预后的最新发现。肠道菌群的变化引起主要的促炎性T细胞反应,这与更大的梗塞体积和恶化的预后相关。
由于肠道菌群是一个非常复杂的生态系统,受环境和宿主的影响,具有冗余功能和协同关系,因此,研究微生物组代谢组学特征以将其转化为临床实验研究是一种更好的策略。
然而,尚无实验数据将特定微生物群合成的化合物与脑损伤后的免疫反应之间的因果关系联系起来。为了在中风患者中成功应用这些发现,有必要进一步研究中风后共生细菌和免疫系统之间相互作用的分子机制。
相关阅读:
参考文献:
Singh, V., Roth, S., Llovera, G., Sadler, R., Garzetti, D.,Stecher, B.,et al. (2016). Microbiota dysbiosis controls the neuroinflammatory response after stroke. Journal of Neuroscience, 36(28), 7428–7440.
Houlden, A., Goldrick, M., Brough, D., Vizi, E. S., Lénárt, N., Martinecz, B., et al. (2016). Brain injury induces specific changes in the caecal microbiota of mice via altered autonomic activity and mucoprotein production. Brain, Behavior, and Immunity, 57, 10–20.
Lee, J., d’ Aigle, J., Atadja, L., Quaicoe, V., Honarpisheh, P., Ganesh, B. P., et al. (2020). Gut microbiota-derived short-chain fatty acids promote poststroke recovery in aged mice. Circulation Research, 127(4), 453–465.
Delgado Jiménez Rosa,Benakis Corinne,The Gut Ecosystem: A Critical Player in Stroke.[J] .Neuromolecular Med, 2020.
Yin, J., Liao, S. X., He, Y., Wang, S., Xia, G. H., Liu, F. T., et al. (2015). Dysbiosis of gut microbiota with reduced trimethylamine-N-oxide level in patients with large-artery atherosclerotic stroke or transient ischemic attack. Journal of the American Heart Association.
Stanley, D., Mason, L. J., Mackin, K. E., Srikhanta, Y. N., Lyras, D., Prakash, M. D., et al. (2016). Translocation and dissemination of commensal bacteria in post-stroke infection. Nature Medicine.
Benakis, C., Brea, D., Caballero, S., Faraco, G., Moore, J., Murphy, M., et al. (2016). Commensal microbiota affects ischemic stroke outcome by regulating intestinal γδ T cells. Nature Medicine, 22(5), 516–523.
Lamas, B., Natividad, J. M., & Sokol, H. (2018). Aryl hydrocarbon receptor and intestinal immunity. Mucosal Immunology, 11(4), 1024–1038.

谷禾健康
电离辐射可以引起造血系统,肠道系统以及心脑血管神经系统的多器官损伤。
放射治疗在临床上被广泛用于治疗多种肿瘤。但是由于放射射线不仅可以杀死癌细胞,还会对周围的正常细胞和组织造成损伤,因此放疗经常会引起各种副作用,比如疲劳,恶心,呕吐和腹泻等等。
肠道是辐射的主要目标,也是肠道菌群的最大生态位。虽然有小部分描述性研究表明,肠道微生物群与辐射损伤之间存在潜在的相关性,但这种关系的具体基础仍然不清楚。
近日,来自美国北卡罗拉纳大学教堂山分校的 Jenny P.Y. Ting 团队在 Science在线发表研究论文,题为“Multi-omics analyses of radiation survivors identify radioprotective microbes and metabolites”。

发现在致死剂量的全身性辐射模型中,肠道微生物群,特别是Lachnospiraceae和Enterococcaceae可以保护小鼠抵抗辐射引起的造血系统和肠道系统的损伤,从而在致死剂量的辐射后存活下来。并且这些有益的微生物在放疗副作用轻微的白血病病人的粪便中含量显著提高。
通过靶向性以及非靶向性代谢组学研究,该研究发现SCFAs和色氨酸代谢物能降低肿瘤坏死因子-α、白细胞介素-6、干扰素-γ等促炎细胞因子的生成,这些细胞因子都是辐射损伤的重要介质。
这些发现提出了肠道微生物群和代谢产物在辐射激发后疾病易感性调节中起关键作用的可能性。
研究人员发现,一小部分小鼠可以在高剂量的辐射下存活并正常生活。这些“精英幸存者”拥有一个独特的肠道微生物组,该微生物组是在辐射后形成的。
然后将这部分小鼠称为“精英存活小鼠”,并且收集了他们的粪便进行16srRNA的测序。与同年龄的对照小鼠相比,“精英存活小鼠”的肠道菌群有很明显的不同。
随后,研究人员通过多种不同的菌群干扰实验来直接验证肠道菌群是否可以影响机体对于辐射的反应。在“dirty cage sharing”(脏笼)实验中,作者将“精英存活小鼠”和同年龄常规饲养的的对照小鼠使用过的脏笼子收集起来,用来饲养SPF小鼠和接受者小鼠。
每一周他们都会将接受者小鼠转移到相对应的新鲜脏笼中。经过8周的 “dirty cage” 实验,对接受者小鼠进行致死剂量的辐射。
接受了 “精英存活小鼠” 脏笼子饲养后的接受者小鼠对致死剂量辐射具有了明显的抵抗效果,30天内生存率达到了70%。相反,接收对照组脏笼子饲养的接受者小鼠,30天存活率仅为20%左右。
另外,作者分别利用SPF小鼠和无菌小鼠进行了粪菌移植实验。
接下来他们直接收集了“精英存活小鼠”和对照小鼠的粪便,并将粪便处理物通过灌胃的方法转移到受体小鼠中。接收“精英存活小鼠”粪便移植的SPF小鼠和GF小鼠都表现出明显的抗辐射效果。
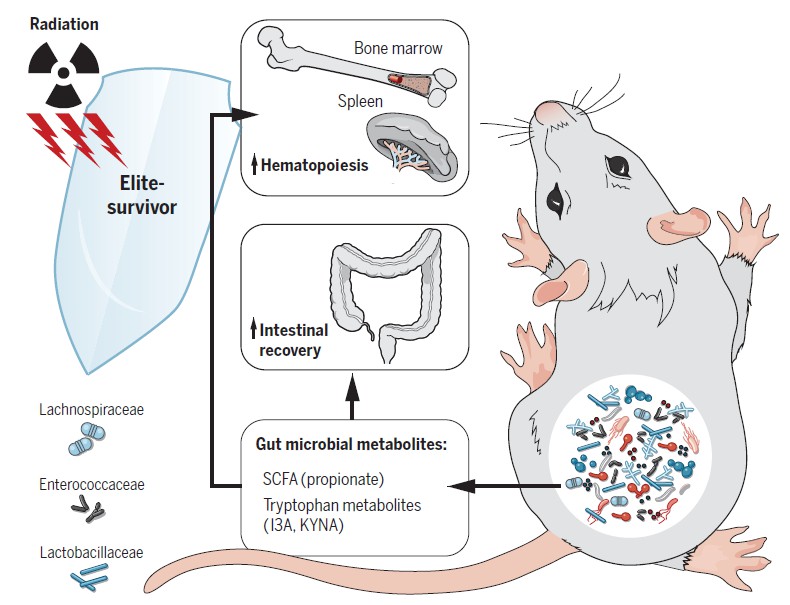
利用这一发现,结合粪便植入和‘dirty cage sharing’实验,证明来自精英幸存者的微生物群在无菌的和常规饲养的接受者中均提供了重要的放射防护,其特点是提高了存活率并改善了临床评分。
他们找出了8种在“精英存活小鼠”受体组显著升高的细菌以及5种显著下降的细菌。那么作者推测,这些在“精英存活小鼠”受体组显著升高的细菌就可能是使机体对辐射有抵抗效果的“有益菌”,相反在“精英存活小鼠”受体组显著下降的细菌则可能是“有害菌”。
那么,这些肠道菌群对辐射的影响在人体内是否也成立呢?
作者联合杜克大学以及纪念斯隆-凯特琳癌症中心的实验室,一起进行了一个小型临床研究。
他们收集了一些白血病病人的粪便并进行了16srRNA测序。由于腹泻是放疗之后常见的副作用,他们根据病人腹泻的持续时间,将病人样本分为轻度腹泻和重度腹泻两组。
肠道菌群分析
通过肠道菌群的分析发现,在轻度腹泻病人中,Lachnospiraceae,Enterococcaceae以及Lactobacillaceae这三种在小鼠中可能的“有益菌”数量明显高于重度腹泻的病人。Lachnospiraceae的数量与病人腹泻时间存在显著地负相关性。因此,作者推断在人体中,这些可能的“有益菌”也可以帮助病人降低放疗引起的副作用。
随后,作者挑选了三种可能的“有益菌”,三种可能的“有害菌”以及一种最著名的益生菌 Lactobacillus rhamnosus, 进行了单一菌株的体内实验。
组织学分析
作者发现,Lachnospiraceae对小鼠的保护作用极明显。并且,通过组织学分析,Lachnospiraceae极大地降低了辐射对于造血系统和肠道系统造成的损伤。骨髓和脾脏的组织结构和细胞死亡得到了极大地保护。大肠和小肠的损伤以及肠道渗透性也明显降低。
短链脂肪酸分析
由于Jenny Ting课题组和其他课题组已经报道过,Lachnospiraceae可以产生大量的短链脂肪酸,并且短链脂肪酸是调节机体免疫反应和炎症反应的重要物质,因此,作者检测了三种短链脂肪酸对辐射的影响。
他们发现,在接受了propionate丙酸处理后,小鼠可以有效地抵抗致死剂量辐射带来的造血系统和肠道系统的损伤,生存率也得到了极大的提高。
细胞分析
作者同时检测了骨髓中造血干细胞和祖细胞的比例,发现propionate处理小鼠要远高于对照小鼠。另外,辐射所导致的DNA损伤与活性氧的释放,在propionate处理后也明显降低。
非靶向性代谢组学分析
最后,作者通过非靶向性代谢组学研究发现了一系列的代谢产物在“精英存活小鼠”的粪便中含量显著提高。其中,升高最为明显的是色氨酸代谢通路的成员。作者选取了indole-3-carboxaldehyde (I3A)和kynurenic acid(KYNA)进行了体内验证。两个色氨酸途径代谢产物1H-吲哚-3-甲醛(I3A)和强尿酸(KYNA)在体内提供了长期放射防护。
该研究的发现强调肠道菌群作为宿主抵抗辐射的主要调节剂的关键作用,能够保护造血和胃肠系统。
“精英存活小鼠” 的肠道菌群组成和普通小鼠不同,他们的肠道中含有大量的Lachnospiraceae等“有益菌”。这些“有益菌”通过产生大量的短链脂肪酸和色氨酸代谢通路的代谢产物来促进造血发生以及肠道损伤修复,从而帮组机体抵抗辐射引起的损伤和死亡。
毛螺菌科和肠球菌科,以及以丙酸和色氨酸途径菌群为代表的下游代谢产物,对辐射防护起着重要作用。
这项研究揭示了微生物代谢物轴在产生广泛的抗辐射保护中所起的关键作用,并为治疗辐射暴露的不良副作用提供了有希望的治疗靶标。
Tips
该文做了很多生理,化学,生物代谢和病理等大小20多项实验,包括小鼠实验,辐射处理实验,“dirty-cage sharing”实验,FMT实验,细菌菌种管理实验,SCFA和色氨酸代谢处理实验,病理组织学,肿瘤接种和放射治疗实验,免疫染色和免疫印迹解折,肠通透性检测,骨髓干细胞,肠上皮细胞和肠上皮内淋巴细胞分离,ROS检测,流式细胞仪分析,粪便16S rRNA基因测序分析,气相色谱-质谱检测,非靶向代谢组实验。一篇高水平论文的研究思路和严谨的实验论证过程,值得我们借鉴和学习。
参考文献:
Guo Hao,Chou Wei-Chun,Lai Yunjia et al. Multi-omics analyses of radiation survivors identify radioprotective microbes and metabolites.[J] .Science, 2020, 370

谷禾健康
痤疮(俗称“痘痘”)是一种常见的皮肤病,会影响全球近10%的人口。导致痤疮发展的因素很多,包括皮脂和角蛋白的产生,引起痤疮的细菌,激素,毛孔阻塞和炎症。
饮食与痤疮之间的联系一直存在争议,但最近的研究表明饮食在痤疮的发生中可以发挥重要作用。
本文将列举以下7种可能引起痤疮的食物,并提供一些相应的饮食建议。


吃大量精制碳水化合物可能会增加血糖和胰岛素水平,并促进痤疮的发生。
与很少或没有痤疮的人相比,有痤疮的人往往消耗更多的精制碳水化合物。
富含精制碳水化合物的食物包括:
用白面粉制成的面包,饼干,谷类食品或甜点;
白面粉意面;
白米饭和米粉;
苏打水和其他含糖饮料;
甜味剂,如蔗糖,枫糖浆,蜂蜜;
…
一项研究发现,经常食用添加糖的人患痤疮的风险增加30%,而那些经常吃糕点和蛋糕的人患痤疮的风险则增加20%。
这可以通过精制碳水化合物对血糖和胰岛素水平的影响来解释。
精制的碳水化合物会迅速吸收到血液中,从而迅速提高血糖水平。当血糖升高时,胰岛素水平也会升高,以促进血糖从血液中传出并进入细胞。
但是,高水平的胰岛素对痤疮患者并不友好。胰岛素可使雄激素更加活跃,并增加胰岛素样生长因子1(IGF-1)。通过使皮肤细胞更快生长并促进皮脂生成,这有助于痤疮的发展。
另一方面,低升糖指数饮食法不会显著提高血糖或胰岛素水平,却可以减少痤疮的严重程度。
当然,仍需要进一步研究,来了解精制碳水化合物如何促进痤疮。


经常食用乳制品与痤疮严重程度的增加有关
许多研究发现,乳制品与青少年痤疮严重程度之间存在关联。
两项研究还发现,经常食用牛奶或冰淇淋的年轻人患粉刺的几率是不经常吃的人的 4 倍。
但是,迄今为止的研究主要集中在青少年和年轻人,并且仅显示了牛奶和痤疮之间的相关性,而不是因果关系。尚不清楚牛奶如何促进痤疮的形成,但是有几种理论:
已知牛奶会增加胰岛素水平,而不受其对血糖的影响,这可能会使痤疮严重程度恶化。
牛奶还含有刺激肝脏产生更多IGF-1的氨基酸,这与痤疮的发生有关。
需要进行更多的研究,以确定是否存在特定数量或类型的乳制品,可能加剧痤疮。
西式快餐

经常吃西式快餐与痤疮发生的风险增加有关
痤疮与进食富含卡路里,脂肪和精制碳水化合物的西式饮食密切相关。
西式快餐食品,例如汉堡,矿块,热狗,炸薯条,苏打水和奶昔,是典型西方饮食中常见的食物,可能增加痤疮的风险。
一项针对 5000 多名中国青少年的研究发现,高脂饮食会使痤疮发生风险增加43%。定期吃西式快餐会增加17%的风险。
另一项针对 2300 名土耳其男子的单独研究发现,经常吃汉堡或香肠与痤疮发生风险增加24%有关。
目前尚不清楚为什么吃西式快餐会增加患痤疮的风险,但一些研究人员提出,它可能以促进痤疮发展的方式影响基因表达并改变激素水平。
必须要说明的是,大多数有关西式快餐和痤疮的研究都使用了自我报告的数据。这种类型的研究仅显示饮食习惯和痤疮风险的关联,而不能证明西式快餐会直接导致痤疮。因此,需要更多的研究。
富含Omega-6脂肪的食物

富含omega-6脂肪酸和omega-3含量低的饮食具有促炎作用,并且可能使痤疮恶化。
跟典型的西方饮食一样,含有大量omega-6脂肪酸的饮食与炎症和痤疮的增加有关。
这可能是因为西方饮食中含有大量的玉米和豆油,它们富含omega-6脂肪,而很少包含omega-3脂肪的食物,例如鱼和核桃。
omega-6和omega-3脂肪酸的这种不平衡会促使人体进入炎症状态,这可能会加剧痤疮的严重性。相反,补充omega-3脂肪酸可降低炎症水平,并可减轻痤疮严重程度。
尽管Omega-6脂肪酸和痤疮之间有关联,但尚未对此主题进行随机对照研究,需要进行更多研究。
巧克力

研究支持食用巧克力和痤疮之间的联系
自上世纪二十年代以来,巧克力就一直被认为是痤疮的诱因,但到目前为止,尚未达成共识。
多项非正式调查已将食用巧克力与增加痤疮的风险联系起来,但这不足以证明巧克力会引起痤疮。
最近的一项研究发现,易长粉刺的男性每天食用25克99%黑巧克力,仅在两周后出现了更多的痤疮病变。
另一项研究发现,与服用安慰剂的男性相比,每天服用100%可可粉胶囊的男性,在一周后的痤疮病变明显更多。
至于巧克力为什么会增加痤疮的确切原因还不太清楚,有一项研究发现,吃巧克力会增加免疫系统对引起痤疮细菌的反应,这可能有助于解释上述发现。
乳清蛋白粉

少量数据表明,服用乳清蛋白粉与痤疮之间存在联系。
乳清蛋白粉是一种流行的膳食补充剂。
它是氨基酸、亮氨酸和谷氨酰胺的丰富来源。这些氨基酸可使皮肤细胞生长和分裂更快,这可能有助于痤疮的形成。
乳清蛋白中的氨基酸还可以刺激人体产生更高水平的胰岛素,这与痤疮的发生有关。
一些案例研究报告了男性运动员乳清蛋白消耗与粉刺之间的联系。另一项研究发现痤疮严重程度与乳清蛋白补充剂天数之间的直接相关性。
这些研究支持乳清蛋白和痤疮之间的联系,但需要更多的研究来确定乳清蛋白是否引起痤疮。
个人过敏食物
食物敏感性反应可增加体内的炎症程度,理论上可能会加剧痤疮。
有人提出,痤疮从根本上说是一种炎症性疾病。事实证明,抗炎药如皮质类固醇是治疗严重痤疮的有效方法,而且痤疮患者血液中的炎症分子水平升高。
食物导致炎症的一种方法是通过食物敏感性,也称为迟发型超敏反应。
当免疫系统错误地将食物识别为威胁并对其发起免疫攻击时,就会出现食物过敏。这会导致高水平的促炎分子在体内循环,从而加剧痤疮。
尽管炎症和痤疮之间似乎存在联系,但尚无研究直接调查食物敏感性在其发展中的特定作用。这是一个值得研究的领域,有助于更好地了解食物,免疫系统和炎症如何影响痤疮的发生。
知道自己对哪些食物过敏,就别吃了吧~
以上7种是可能引起痤疮的食物,那么到底该吃什么呢?
以下建议可供参考。
Omega-3脂肪酸
Omega-3具有抗发炎作用,经常食用富含Omega-3的食物可降低痤疮的发生风险。
益生菌
益生菌可促进健康的肠道和平衡的微生物组,这与减少炎症和降低痤疮发生的风险有关。
绿 茶
绿茶含有多酚,与减少发炎和降低皮脂产生有关。绿茶提取物被发现可以减少痤疮的严重程度。
姜 黄
姜黄含有抗炎多酚姜黄素,可帮助调节血糖,提高胰岛素敏感性并抑制引起痤疮细菌的生长,从而减少痤疮。
维生素A,D,E和锌
这些营养物质在皮肤和免疫健康中起着至关重要的作用,可能有助于预防痤疮。
旧石器时代饮食
旧石器时代饮食富含瘦肉,水果,蔬菜和坚果,而谷物,奶制品和豆类则少。与降低血糖和胰岛素水平有关。
地中海式饮食
地中海式饮食富含水果,蔬菜,全谷物,豆类,鱼和橄榄油,而乳制品和饱和脂肪则少。它也与降低痤疮严重程度有关。
结 语
你可以不必完全避免与痤疮有关的所有食物(一般也做不到
),但应与上述其他营养丰富的食物一起,保持均衡饮食。

小 妙 招
如果有时间,可以坚持把吃的东西记录下来,看看某些症状是否与特定食物有关联。
长此以往,你可以慢慢去寻找正在吃的食物和皮肤健康之间的某种关系,那也将会是属于你个人的饮食宝典。
主要参考文献:
向上滑动阅览
Burris J, Rietkerk W, Woolf K. Acne: the role of medical nutrition therapy. J Acad Nutr Diet. 2013 Mar;113(3):416-30. doi: 10.1016/j.jand.2012.11.016. PMID: 23438493.
Burris J, Rietkerk W, Shikany JM, Woolf K. Differences in Dietary Glycemic Load and Hormones in New York City Adults with No and Moderate/Severe Acne. J Acad Nutr Diet. 2017 Sep;117(9):1375-1383. doi: 10.1016/j.jand.2017.03.024. Epub 2017 Jun 9. PMID: 28606553.
Blasbalg TL, Hibbeln JR, Ramsden CE, Majchrzak SF, Rawlings RR. Changes in consumption of omega-3 and omega-6 fatty acids in the United States during the 20th century. Am J Clin Nutr. 2011 May;93(5):950-62. doi: 10.3945/ajcn.110.006643. Epub 2011 Mar 2. PMID: 21367944; PMCID: PMC3076650.
Khayef G, Young J, Burns-Whitmore B, Spalding T. Effects of fish oil supplementation on inflammatory acne. Lipids Health Dis. 2012 Dec 3;11:165. doi: 10.1186/1476-511X-11-165. PMID: 23206895; PMCID: PMC3543297.
Aksu AE, Metintas S, Saracoglu ZN, Gurel G, Sabuncu I, Arikan I, Kalyoncu C. Acne: prevalence and relationship with dietary habits in Eskisehir, Turkey. J Eur Acad Dermatol Venereol. 2012 Dec;26(12):1503-9. doi: 10.1111/j.1468-3083.2011.04329.x. Epub 2011 Nov 10. PMID: 22070422.
Adebamowo CA, Spiegelman D, Danby FW, Frazier AL, Willett WC, Holmes MD. High school dietary dairy intake and teenage acne. J Am Acad Dermatol. 2005 Feb;52(2):207-14. doi: 10.1016/j.jaad.2004.08.007. PMID: 15692464.
Melnik BC, John SM, Schmitz G. Milk is not just food but most likely a genetic transfection system activating mTORC1 signaling for postnatal growth. Nutr J. 2013 Jul 25;12:103. doi: 10.1186/1475-2891-12-103. PMID: 23883112; PMCID: PMC3725179.
Vongraviopap S, Asawanonda P. Dark chocolate exacerbates acne. Int J Dermatol. 2016 May;55(5):587-91. doi: 10.1111/ijd.13188. Epub 2015 Dec 29. PMID: 26711092.
Bar-Peled L, Sabatini DM. Regulation of mTORC1 by amino acids. Trends Cell Biol. 2014 Jul;24(7):400-6. doi: 10.1016/j.tcb.2014.03.003. Epub 2014 Mar 31. PMID: 24698685; PMCID: PMC4074565.
Silverberg NB. Whey protein precipitating moderate to severe acne flares in 5 teenaged athletes. Cutis. 2012 Aug;90(2):70-2. PMID: 22988649.
Pietschmann N. Food Intolerance: Immune Activation Through Diet-associated Stimuli in Chronic Disease. Altern Ther Health Med. 2015 Jul-Aug;21(4):42-52. PMID: 26030116.

谷禾健康
痘痘,粉刺,酒渣鼻等皮肤问题,困扰着一些爱美的朋友。这关乎到个人形象,甚至有些朋友还会因为这个感到焦虑,社交尴尬,严重影响自信。
为了祛痘,盲目往脸上涂抹太多祛痘护肤产品,正所谓“过犹不及”,涂抹太多,过量,不断刺激皮肤,雪上加霜。


改善皮肤和整体健康的关键,从你的肠道开始。
我们常常说,一个人由内而外散发着美。其实是有科学依据的。
你的肠道和皮肤有很多共同点:
· 都在保护你,不受外界影响。
· 各自都有微生物群,彼此直接交流。
· 都富含肥大细胞,是免疫系统抗病第一线。
· 它们在早期发育过程中来自同一类细胞。
为什么吃的东西会影响你的皮肤?
皮肤与周围的世界相互作用,而胃肠道则处理你吃的、呼吸的和吞咽的一切。你吃的食物经过新陈代谢,食物分解后的副产物会直接进入到你的皮肤。
把皮肤看成是肠道健康的窗口
以下是11种皮肤状况,实际上是从肠道开始的:


看过我们文章的小伙伴都知道,人体肠道中有大量的微生物群。那肠道微生物群是怎么和皮肤进行交流的呢?
这就涉及一个:肠道-皮肤轴。
肠道-皮肤轴是连接肠道及其微生物与皮肤的信号分子网络,跟脑-肠轴类似。

免疫系统、皮肤和大脑之间的沟通是通过微生物来调节的。
炎症—— 大多数疾病的根本原因。
氧化应激—— 炎症的主要原因。
组织脂质水平—— 健康新陈代谢的重要因素。
血糖—— 控制平衡血糖的能力。
神经肽水平—— 一个与你的情绪、疼痛耐受力和身体内环境平衡有关的因素。
致病菌—— 使你生病或引起疾病的坏细菌。
神经递质—— 调节情绪,例如,大约90%的血清素(你的快乐神经递质)是由肠道微生物群产生的。
肠道微生物组与以上协调皮肤稳态的各个途径都有关联,最终影响皮肤健康。
研究发现,痤疮与其说是皮肤的问题,不如说是肠道的问题。
脂多糖(LPS)内毒素是有害的革兰氏阴性菌的组成成分,已被证明对肠道和皮肤造成严重破坏。研究发现,当体内有高水平的脂多糖内毒素时,它们会干扰伤口愈合,使疤痕更严重。
LPS还与寻常痤疮(也就是粉刺)有关,使你更可能对大肠杆菌脂多糖内毒素(E.coli LPS)产生强烈反应。对E.coli LPS有很强的反应性意味着你更可能有纤维蛋白微凝块,这会导致小而疼痛的组织疤痕。
更可怕的是,高水平的脂多糖和大肠杆菌脂多糖会导致肠漏,使细菌进入血液,身体就会出现炎症,表现在皮肤上就是痤疮、粉刺等问题。它还会使你更容易患上肠易激综合征和焦虑症、抑郁症等。
另外,肠道菌群失衡,有害菌大量繁殖,促进炎性物质释放,从而也会引起痤疮等皮肤问题。
特应性皮炎是最常见的慢性瘙痒性炎症性皮肤病。
西方饮食的低纤维和高脂肪含量特征从根本上改变了肠道菌群,导致免疫调节代谢产物特别是短链脂肪酸的产生不足。短链脂肪酸对上皮屏障完整性起重要作用。
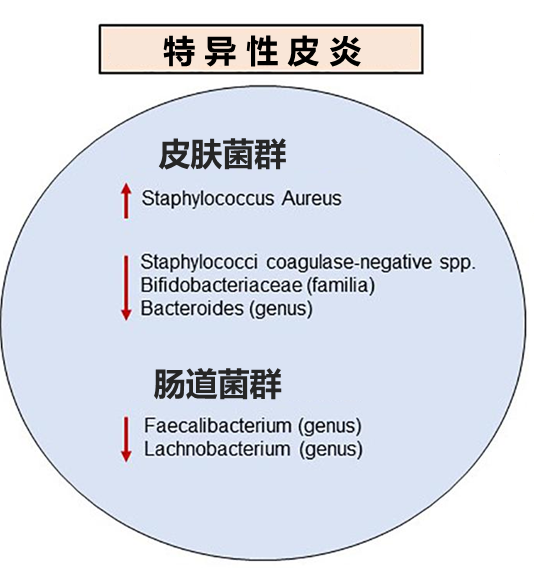
在两项韩国研究中,对特异性皮炎患者粪便样本的宏基因组分析显示,与对照组相比,普氏栖粪杆菌Faecalibacterium prausnitzii显著减少。
关于普氏栖粪杆菌的失调与上皮屏障破坏之间可能存在正反馈回路。肠屏障的破坏,允许消化不良的食物、微生物和毒素进入循环,到达目标组织包括皮肤,触发Th2免疫反应,导致进一步的组织损伤,从而促进该反馈回路。
关于特异性皮炎,更多详见:微生物群对三大过敏性疾病发展的影响
银屑病(俗称牛皮癣),是一种由免疫介导的慢性炎症性皮肤病,病程较长且易复发。
牛皮癣与胃肠道炎症的相关性特别强,7 – 11%的炎症性肠病(IBD)患者被诊断为牛皮癣。
某些共同的遗传和环境因素以及免疫途径在这两种疾病的发病机制中都有牵连。例如,Th17细胞及其细胞因子,已知在牛皮癣的发展中起主要作用,在IBD的病理生理学中也有牵连。
牛皮癣患者肠道菌群特征与IBD患者类似。与健康对照组相比,牛皮癣患者体内普氏栖粪杆菌Faecalibacterium prausnitzii的数量显著减少。
湿疹始于肠道。
当医生和皮肤科医生在寻找皮肤表面的解决方案时,他们忽略了湿疹是由免疫系统过度活跃引起的事实。这种过度活跃的免疫系统通常是肠道微生物群失调的结果。事实上,微生物多样性的下降会导致湿疹发作。
为了缓解湿疹症状,采取措施减少炎症和通过饮食增加微生物多样性是至关重要的。对抗湿疹的饮食需要针对你的微生物群和身体。
乳糜泻乳糜泻是一种常见的慢性疾病。它和皮肤状况密切相关。
患有乳糜泻的人更有可能有:
疱疹样皮炎、脱发、湿疹、
荨麻疹、白癜风、口腔黏膜病变
因为当患有乳糜泻的时候,免疫系统会释放出具有防御性质的抗体,也就是免疫球蛋白,它会在皮肤下面血管集中,导致皮疹的出现。
患有乳糜泻的人往往需要解决肠道的问题。他们有一个优势,就是他们会被引导去寻找与肠道有关的解决方案。
这就意味着,当他们进行无麸质或其他措施来减少乳糜泻的影响时,顺便在不经意间解决了皮肤状况。
至于其他的皮肤问题,往往需要比乳糜泻花更多的时间,才意识到原来肠道是问题的根源。
医生正在发现小肠细菌过度生长(SIBO)患者与酒渣鼻之间有着密切的联系。一些医生甚至报告说,当他们为患者治疗SIBO时,他们多年来的酒渣鼻会突然消失。
这对酒渣鼻患者来说绝对是个好消息。
SIBO是在2004年才被发现,许多人患有酒渣鼻,却不知道问题的根源可能是他们的肠道。现在可能许多人还不知道这一重要的联系。
每个人的肠道微生物组对他们来说都是独一无二的,因此每个人都有不同的适合自己的方法。
这里有些比较普适的建议。
减轻生活压力

持续的压力会对肠道菌群产生负面影响。减少压力时,进行规律饮食,保证良好睡眠,可以适当进行冥想、瑜伽、听音乐等方式,当然改善肠道健康也同样重要。
少吃糖

各种甜食虽然诱人,但还是尽量不要吃太多。糖分会刺激皮脂腺分泌,吃过多会导致皮肤爱出油、出油多自然容易长痘痘、粉刺等。另外,糖可以喂食许多有害菌,当你少吃糖时,提供给有害菌的能量少了,有害菌就会被饿死。
详见:深度解析 | 炎症,肠道菌群以及抗炎饮食 文中除了详细介绍机制外,还有相关的抗炎和促炎食物介绍。
均衡饮食
健康的饮食,最好由多种不同类型的食物组成,包含水果、蔬菜、谷物、肉类等。丰富的食物有利于肠道内不同类型细菌的生长,从而帮助肠道菌群多样性的增加。菌群多样性可能对恢复力起到积极作用,更能抵抗外界刺激引起的微生物群的扰动。
适量锻炼

适量锻炼,但不要太过 。锻炼对肠道微生物组有益,但如果锻炼过多,则无济于事。过度劳累又相当于形成了一种压力,并可能导致微生物失衡。
肠道菌群检测
每个人的肠道菌群都需要独特的饮食。对别人健康的食物可能对你来说并不理想。肠道菌群检测可以是一种工具,来帮你找到最适合你自己的健康食物。
其他方式,详见:
结 语
我们都知道分析问题要有整体观。面对疾病也一样,表面看是皮肤问题,实际可能是肠道的问题。
调理肠道健康,从根本上入手,一些皮肤问题也就迎刃而解了,而皮肤的改善提升颜值,带来的心情愉悦也可以进一步促进健康。
相关阅读:
参考文献:
Guo, Shuhong et al. “Lipopolysaccharide causes an increase in intestinal tight junction permeability in vitro and in vivo by inducing enterocyte membrane expression and localization of TLR-4 and CD14.” The American journal of pathology vol. 182,2 (2013): 375-87. doi:10.1016/j.ajpath.2012.10.014
O’Neill CA, Monteleone G, McLaughlin JT, Paus R. The gut-skin axis in health and disease: A paradigm with therapeutic implications. Bioessays. 2016 Nov;38(11):1167-1176.
Huang R, Wang K, Hu J. Effect of Probiotics on Depression: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients. 2016;8(8):483. Published 2016 Aug 6. doi:10.3390/nu8080483
Bowe WP, Logan AC. Acne vulgaris, probiotics and the gut-brain-skin axis – back to the future? Gut Pathog. 2011 Jan 31;3(1):1. doi: 10.1186/1757-4749-3-1. PMID: 21281494; PMCID: PMC3038963.
Caproni M, Bonciolini V, D’Errico A, Antiga E, Fabbri P. Celiac disease and dermatologic manifestations: many skin clue to unfold gluten-sensitive enteropathy. Gastroenterol Res Pract. 2012:952-753.
Parodi A, Paolino S, Greco A, Drago F, Mansi C, Rebora A, Parodi A, Savarino V. Small intestinal bacterial overgrowth in rosacea: clinical effectiveness of its eradication. Clin Gastroenterol Hepatol. 2008 Jul;6(7):759-64.
Wollina U. Microbiome in atopic dermatitis. Clin Cosmet Investig Dermatol. 2017;10:51-56.
Juhlin L, Michaëlsson G. Fibrin microclot formation in patients with acne. Acta Derm Venereol. 1983;63(6):538-40. PMID: 6198846
Yang H, Hu C, Li F, Liang L, Liu L. Effect of lipopolysaccharide on the biological characteristics of human skin fibroblasts and hypertrophic scar tissue formation. IUBMB Life. 2013 Jun;65(6):526-32. doi: 10.1002/iub.1159. Epub 2013 May 7. PMID: 23653386

谷禾健康
说起蟑螂,大部分人都不太喜欢,甚至生理性恐惧…
你可能有100个讨厌它的理由,见到它就想消灭它,然而怎么都灭不完…


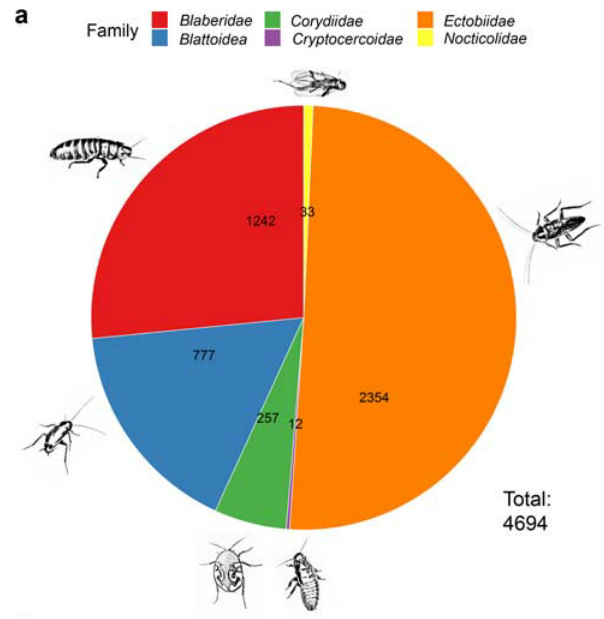
算起来,蟑螂早在3亿年前就已经存在了。蟑螂在线数据库中登记了4685种蟑螂(截止2020.8)。它拥有极强的适应力,亿万年来生生不息,广泛分布在世界各个角落。


可以忍受很长时间的饥饿和脱水。例如,美洲大蠊在没有食物和水的情况下可以存活1个月。
一些雌性蟑螂在交配后会吃掉雄性蟑螂。
蟑螂也可以通过单性生殖繁殖。
一些雌性蟑螂可以在一次交配后终生储存精子,使卵在没有雄性存在的情况下受精。
几乎所有的蟑螂都有内共生细菌,能够将尿素和氨水中的氮回收到氨基酸中。

的确,蟑螂可以通过粪便等方式传播潜在的致病菌,给我们造成了一定的困扰;然而,从另一方面考虑,尽管蟑螂可能携带致病细菌,但大多数菌株是无害的,细菌产生代谢物或蛋白质,具有潜在的工业应用价值。
那么,如何“变害为宝”,在生物技术应用中发挥价值?本文来为你详细介绍下。
超过99%的蟑螂不是寄居性害虫,而是生活在腐烂的树叶中、树皮下、石头下或土壤中,而这一巨大而未开发的资源很可能提供丰富的微生物多样性来源。
Blattabacterium 菌株 BPLAN基因组详细分析显示,13%的蛋白质编码基因用于氨基酸生物合成,7.8%用于辅酶的生产。昆虫宿主将尿酸(以尿酸盐的形式储存在尿细胞中)转化为尿素,然后由Blattabacterium 输入并转化为氨。
布拉德细菌酶谷氨酸脱氢酶(GdhA)催化将氨加到2-氧代戊二酸中,生成D-谷氨酸,然后可以将其转化为大多数其他氨基酸。
此外,蟑螂粪便中的真菌活性与微生物产生的化合物有关。
昆虫被认为是产生生物活性分子的微生物的重要来源,尤其是蟑螂,鉴于其独特的进化历史和弹性,它们是具有代谢能力细菌的家园。
蟑螂是人类住所和医疗环境中的危险害虫,是细菌感染的载体。两项经典的研究将卫生中心消灭蟑螂与较低的感染频率,特别是鼠伤寒沙门氏菌和肝炎发病率联系在一起。
有强有力的证据表明,幽门螺杆菌、分枝杆菌、铜绿假单胞菌、沙门氏菌和其他细菌能够通过蟑螂消化系统存活下来。它们通常以腐烂的有机物质为食,因此能传播致病菌。此外,蟑螂的一些细菌分离物显示出抗生素耐药性。
然而,没有直接证据表明蟑螂会传播医院感染,根除蟑螂与降低感染率之间的联系可能与更普遍的清洁度改善有关。最近一项强调蟑螂与传染病之间关系的综述得出结论,尽管蟑螂会增加风险,医院不应容忍蟑螂,但目前还没有确切证据证明蟑螂直接参与了传染病向患者的传播。
从蟑螂中培养出来的大多数细菌是变形菌门。
培养方法的一个主要优点是它有助于对分离的细菌进行基因组、转录组学、蛋白质组学和代谢组学分析,允许直接的生物技术开发。
另一方面,非培养方法可以更真实地了解微生物生态位的真实多样性,是分析细菌群落的金标准。然而,对数据的解释应谨慎,因为随着时间的推移,菌群组成的变化以及同一菌群个体之间的差异,很难对数据进行概括。
微生物组学研究结果受到生物因素的影响,如年龄、性别和健康状况;环境因素,如饮食、温度和湿度;以及分析方法(DNA提取、引物、测序、数据库和生物信息学流程)的差异,虽然影响程度较小。
据研究,蟑螂菌群主要的门是厚壁菌,其次是拟杆菌,最后是变形菌,三者加起来占细菌多样性的80%以上。
厚 壁 菌 门
厚壁菌在蟑螂的中肠最丰富( 43%),前肠30%,后肠34%,中肠的碱性更强(pH值6.1–8.9),大量嗜碱需氧细菌属芽孢杆菌、拟杆菌属和肠球菌在那里大量繁殖。

芽孢杆菌
芽孢杆菌属很容易从蟑螂中培养出来,特别是与蜡样芽孢杆菌和枯草芽孢杆菌有关的菌株。
分子微生物学研究表明,在蟑螂中广泛存在芽孢杆菌科的不同分支,包括芽孢杆菌属、地芽孢杆菌属Geobacillus、海洋芽孢杆菌属Oceanobacillus、尿素芽孢杆菌属Ureibacillus以及其他相关分类群,如类芽孢杆菌属Paenibacillus和赖氨酸芽孢杆菌属Lysinibacillus以及几个未分类群。
蜡样芽胞杆菌的丝状形态被称为节状体,附着在肠道上皮细胞上。蜡样芽孢杆菌和相关菌,如炭疽芽孢杆菌和苏云金芽孢杆菌可以分泌裂解酶和毒素,可在人类和其他动物中引起疾病。
蜡样芽胞杆菌B1分离自克氏芽胞杆菌Blaberus craniifer, 对马德拉大蠊具有高致病性,磷脂酶C对这种作用负责。
从蟑螂中分离出的枯草芽孢杆菌BGI-1菌株,对球孢白僵菌Beauveria bassiana表现出很强的杀菌活性。
另一种从Pe. americana分离出来的芽孢杆菌菌株29K,在其他芽孢杆菌菌株中表现出最强的角质溶解和蛋白水解活性,在7天内完成羽毛消化。
由此我们可以看到蟑螂芽孢杆菌的潜力,特别是生物活性代谢物和工业上有价值的酶的生产,以及作为生产重组蛋白的平台。
葡萄球菌
金黄色葡萄球菌是最常见的分离种。葡萄球菌大量存在于樱桃蟑螂的后肠中,其他蟑螂中较少。
虽然金黄色葡萄球菌是一种共生细菌,无症状地寄居在约30%的人类群体的皮肤和口腔粘膜上,但它也能获得毒力并引起危险的感染。
蟑螂体内金黄色葡萄球菌的显著存在,使蟑螂有了传播病原体的坏名声,尤其是一些医院蟑螂中分离出了抗生素抗性金黄色葡萄球菌菌株。
乳杆菌
乳杆菌目在蟑螂中普遍存在,特别是肠球菌属和链球菌属。乳酸杆菌属是东方蜚蠊和Pe. americana的一个属。
在无培养方法中,乳杆菌科在枯叶蟑螂Ergaula capuchina和 Byrsotria fumigata 中特别丰富,占40%以上。
肠球菌和乳酸杆菌是常见的肠道细菌,尽管它们的确切作用尚不清楚,但它们可能通过将糖类转化为易于吸收的乙酸盐和乳酸来提高食物的消化率。
一些肠球菌是与医院感染相关的条件致病菌,在昆虫中,它们转移到血淋巴与疾病有关。另一方面,用肠球菌或乳酸杆菌喂养昆虫可以防止苏云金芽孢杆菌引起的死亡。
肠球菌可以产生细菌素来抑制竞争细菌的生长
并且由于它们自然存在于许多手工食品中,因此它们可以直接用作发酵剂培养物或纯化抗生素肽的来源。
例如, 乳酸链球菌肽是一种授权食品防腐剂,用于香肠、奶酪和其他肉类和奶制品。在食品加工过程中,它是安全的、耐热的,但可被胃蛋白酶和胰蛋白酶降解,因此对天然肠道微生物没有影响。
从尼日利亚一只蟑螂中分离出的粪肠球菌E18菌株被证明能产生肠毒素或具有强抗菌活性的细菌素混合物。
我们知道从蜜蜂中分离出的乳酸杆菌已经被提议作为替代益生菌,那么类似的概念也适用于蟑螂的乳酸杆菌。
梭状芽孢杆菌
蟑螂中肠和后肠的缺氧部分是厌氧菌的家园,尤其是梭状芽胞杆菌,梭菌已从Blatta orientalis,Eu. posticus 和 Pe. americana 分离。其他还有属于梭状芽孢杆菌科(梭菌属、Paeniclostridium、Paraclostridium、八叠球菌Sarcina)、真杆菌科(真杆菌属)、毛螺菌科(Blautia、丁酸弧菌属Butyrivibrio、粪球菌属Coprococcus)、颤螺菌科Oscillospiraceae(粪杆菌属、瘤胃球菌属)、Peptoniphilaceae(Finegoldia)和消化链球菌科(梭状芽孢杆菌属、消化链球菌属)
另外还有两个不太常见的厚壁菌纲也从蟑螂身上培养出来的,Tissierellia、Erysipelotrichia。
B. blatticola是一种厌氧杆状细菌,能从D-葡萄糖中产生甲酸盐、乙醇和乙酸盐。
此外,梭状芽孢杆菌目的毛螺菌科和疣微菌科在所有蟑螂的后肠中有特别好的代表性(5-28%)。
有趣的是,韦荣球菌科被认为是畜牧业中益生菌的潜在来源,因为它减少了瘤胃中乳酸的积累,提高了整体生产力,同时也阻止了致病菌在猪和鸡的定殖。
克里斯滕森菌科(Christensenellaceae)是2012年根据从人类粪便中提取的一种物种确定的,它是当今研究的重要焦点,因为它能促进健康,被认为是一种益生菌。这个菌也已通过分子方法在蟑螂(蜚蠊)中检测到,但尚未从蟑螂中培养出。
从蟑螂中分离出的一些梭状芽孢杆菌菌株可以降解羧甲基纤维素,并可用于生物炼制和生物转化过程,将廉价且丰富的纤维素转化为可发酵糖,然后将其转化为乙酸、丙酮、丁醇、乙醇,或其他产物。
富含复杂碳水化合物(如麸皮)的饮食可以提高蟑螂肠道中氢气的生成。这可能是由大量梭菌介导的,也可能是在拟杆菌的帮助下。
由CO2还原产生的乙酰化也发生在蟑螂的内脏和梭状芽孢杆菌Lowell cluster A中,它们是这种转化的部分原因。利用这种过程捕获大气中的二氧化碳有助于减缓气候变化。
拟 杆 菌 门
非培养方法研究表明,拟杆菌门的相对丰度仅次于厚壁菌门。但在某些物种中,如大蠊属和小蠊属的室内蟑螂,拟杆菌目通常是最突出的一门。拟杆菌物种优先定居在蟑螂后肠,其特点是缺氧和还原。拟杆菌是非产孢的专性厌氧菌。
拟杆菌目在蟑螂中最丰富,最常见的科是以树叶和木材为食的蟑螂中的紫单胞菌科Porphyromonadaceae,以及杂食性蟑螂中的理研菌科Rikenellaceae.
拟杆菌属存在于以落叶为食的蟑螂的中肠和后肠,但在以木材为食的蟑螂中尚未发现,这表明这些细菌能够消化纤维素、淀粉和果胶等复杂多糖。
对蟑螂来源的拟杆菌、Dysgonomonas、Paludibacter、Parabacteroides分离株的全基因组测序和体外试验证实了这一发现。这些细菌产生强大的降解酶,在食品/饲料、纺织、造纸和化妆品行业以及农业中具有潜在的应用。
从蟑螂后肠分离得到黄色金黄杆菌菌株FR2。Blattabacterium属于黄杆菌目,蟑螂中现存的代表黄杆菌目的可培育细菌很有趣,因为它们可能是Blattabacterium进化的中间形式。
此外,黄杆菌在工业生物技术的背景下是有用的,因为它们产生柔红霉素色素以及水解酶。
变 形 菌 门
变形菌是蟑螂中最常被培养的细菌,其中丙型变形菌纲尤其丰富。变形菌在前肠最为丰富(~ 48%),中肠(~ 31%)和后肠(~ 19%)较少,可能反映了前肠的酸性和需氧环境(pH 5.0–6.8)。

根据分子研究,肠杆菌目(特别是肠杆菌科)最丰富,其次是黄单胞菌目和假单胞菌目。
沙雷氏菌
沙雷氏菌是蟑螂中最常见的分离属。共有21种得到验证,其中5种(液化沙雷氏菌Serratia liquefaciens、粘质沙雷菌Serratia marcescens、气味沙雷菌Serratia odorifera、普城沙雷菌Serratia plymuthica和深红沙雷菌菌Serratia rubidaea)已被分离。
然而,所有病例的鉴定都是基于表型和生化特征的,并不像16srRNA基因测序那样精确。在健康、生病和死亡的昆虫中都发现了沙雷菌,它们是正常昆虫的菌群。
它们也可能是条件致病菌,尤其是粘质沙雷菌通常在拥挤的群体中引起红色疾病。粘质沙雷菌分泌的几丁质酶和蛋白酶在昆虫血腔中是剧毒的。
嗜虫沙雷氏菌Serratia entomophila通过表达质粒编码的Sep毒素(抑制消化酶的分泌)介导引起新西兰蛴螬的琥珀病。
其他沙雷氏菌物种显然是共生的,例如,沙雷氏菌共生菌是蚜虫中布氏沙雷氏菌的共生菌,为宿主提供必需的代谢物。
肠杆菌科的其他成员,尤其是柠檬酸杆菌属、肠杆菌属、埃希氏菌属和克雷伯氏菌属经常从蟑螂中分离出来,而其他属则不太突出。虽然许多肠杆菌科细菌可以成为重要的人类病原体,但生活方式广泛,基因型和表型多样,因此,致病性无法从分类学推断。
昆虫源性肠杆菌科的生物技术应用包括它们向害虫传递有毒基因的能力,它们被用作植物刺激源和抗菌素代谢物及其在昆虫饲养设施中的用途。Shimwellia blattae最初是从东方小蠊以及后来从其他蟑螂中分离出来的,有趣的是,它可以从头合成钴胺素,并已被开发为维生素B12的生物技术来源。
假单胞菌目
不动杆菌属和假单胞菌属属于假单胞菌目,常从蟑螂中培养出来,而培养方法的研究也表明,它们在潜伏蟑螂中非常丰富(相对丰度>16%)。
由于普遍存在抗生素耐药性,鲍曼不动杆菌Acinetobacter baumanii 或铜绿假单胞菌引起的人类感染在临床实践中难以治疗,但在从蟑螂分离的铜绿假单胞菌菌株中未发现耐药性证据。
假单胞菌在生物技术中非常重要,不仅因为它们产生生物活性代谢物,而且还用于生物修复,是工业过程中强大的溶解酶(蛋白酶、脂肪酶)的来源。
从东方小蠊分离到的嗜麦芽乳杆菌OG2可降解有毒有机氯农药硫丹,将其用作硫源,并将其转化为毒性较低的代谢物。
醋杆菌科
醋杆菌科在昆虫中广泛分布,这一分支菌在工业氧化反应中是有用的(如山梨糖和醋酸的生产),所以蟑螂衍生的醋杆菌科将来有可能被用于这一用途。
鞘氨醇单胞菌
鞘氨醇单胞菌 Sphingomonas paucimobilis 是一种与医院感染相关的条件致病菌,也被应用于生物修复,并且可以合成结冷胶。
粪产碱菌
粪产碱菌Alcaligenes faecalis存在于昆虫病原线虫体内,注射到大蜡螟的血腔内可造成损伤。
昆虫源Alcaligenes表现出抗真菌活性,并与无色杆菌属(Achromobacter)共同产生生物活性特异性代谢产物,类似于嗜酸杆菌Xenorhabdus和发光杆菌Photorhabdus.
脱硫菌
在杂食性蟑螂的后肠,尤其是脱硫菌目和脱硫弧菌目对生物技术过程非常有用,因为它们将硫酸盐还原为硫化物。已应用于受污染的土壤中,降低了可溶有毒镉的生物有效性,而镉仍不溶于硫化物。
放 线 菌 门
放线菌是重要类,因为链霉菌科产生一系列特殊的代谢产物。该门通常在蟑螂中大量存在(相对丰度为13-21%),但在以落叶为食的物种中则较少,双歧杆菌目和棒状杆菌目是最普遍的。
放线菌也存在于木虱蟑螂的产物中,但数量较少(1.7-4.1%)。分枝杆菌是一个经常被培养的属,并且因为一些物种是人类致病菌,能够通过蟑螂消化系统存活下来,蟑螂与分枝杆菌病的传播有关。
微球菌属、棒状杆菌属和角质杆菌属也常从蟑螂中培养出来。值得注意的是,白蚁是链霉菌科细菌的丰富来源,它们具有保护和维持菌落的功能,但从蟑螂中分离出的物种很少。首次报道的与蟑螂相关的链霉菌种被发现由反复感染蟑螂的线虫携带,但“leidnematis链霉菌”这一物种并未得到验证,目前在任何采集中都没有发现。
球形链霉菌WA5-2-7是近年来从蟑螂肠道培养出来的。并被发现与白化链球菌的昆虫分支相匹配。从40 L球形芽孢杆菌WA5-2-7培养液中纯化出具有抗耐甲氧西林金黄色葡萄球菌(MRSA)活性的抗生素放线菌素X2和柯利霉素A。强调蟑螂相关链霉菌的生物技术潜力。
梭 杆 菌 门
梭杆菌门主要见于以木材为食的蟑螂的中肠,相对丰度为2-7%,杂食性蟑螂的后肠中相对丰度为0.2-6.3%。
梭杆菌与拟杆菌相似(它们是革兰氏阴性、无孢子、厌氧菌),但根据16srRNA序列,它们聚集在不同的组中。它们是人类口腔和肠道的正常菌群,但有些物种是致病的,例如,坏死梭杆菌引起口咽感染,称为Lemierre综合征,而F.nucleatum与结肠癌有关。
F.necrophorum从Eu. posticus分离出坏死菌。这种蟑螂也是多形性物种变种的家,在它的生命周期中在杆状和球菌状之间转换。
梭杆菌用于生产琥珀酸的生物技术过程中,也是生物转化应用中作为酶的来源。
螺旋体门(Spirochaetes)
从一些蟑螂中分离出螺旋体门的螺旋形和一般厌氧菌,特别是钩端螺旋体属和密螺旋体属是相关的,因为这些属的一些物种是公认的人类病原体。
分子生物学研究表明,在蟑螂C. punctulatus和E. capucina的后肠中有大量的螺旋体,相对丰度达到8%左右,Py. surinamensis的中肠也有。
在从食木蟑螂C.punculatus分离出菌株JC227后,提出了一个新的属(碱性螺旋体),并在该属中添加了其他嗜碱螺旋体,其中包括从白蚁中分离出的一个物种。
碱性纤维素纤维素JC227可在高pH值和高盐浓度下降解纤维素,因此适用于洗涤剂工业中纤维素材料的生物转化。
浮霉菌门(Planctomycetes)
浮霉菌存在于以木材为食的蟑螂和杂食性蟑螂的后肠中,其相对丰度为0.2-8.4%。
浮霉菌能产生多种特殊的代谢物,发现具有未知代谢潜力的新物种可能会在医药和农药工业中带来新的应用。
疣微菌门(Verrucomicrobia)
据培养方法研究,该门在木食和杂食性蟑螂中有所代表,但从蟑螂中只培养出一种疣微菌门的单一菌株。
从蟑螂Sh. lateralis后肠中分离到一种微需氧植物黄褐假丝酵母(Ereboglobus luteus)。这种细菌可以代谢果胶,但不能在好氧环境下发育。
疣微菌是厌氧污泥降解群落的主要组成部分,因此可以开发用于厌氧生物炼制。
大多数与蟑螂相关的细菌都属于变形菌,厚壁菌和拟杆菌,合起来占微生物总数的80%以上。
在蟑螂中易于培养的细菌中,大多数研究集中在革兰氏阴性大肠菌,葡萄球菌和分枝杆菌的分离上,因此,蟑螂被认为是不健康的害虫,对健康构成威胁。的确,蟑螂可在粪便中传播病原体,应从食物制备区域和医疗机构中消除。
但是,蟑螂也可以是生物技术上有用有前途的微生物资源,因为蟑螂与细菌共同进化,因此与土壤细菌相比,它们可能产生毒性较小的抗菌剂,以确保宿主存活。
当然也许更重要的是扩大微生物在生物精炼厂,生物修复以及药物和农用化学产品以及工业酶的开发中的生物技术应用。
参考文献:
Lee B, Hwang JS, Lee DG (2019) Induction of apoptosis-like death byperiplanetasin-2 in Escherichia coli and contribution of SOS genes.Appl Microbiol Biotechnol 103(3):1417–1427
Chen Z, Ou P, Liu L, Jin X (2020) Anti-MRSA activity of actinomycinX2 and collismycin a produced by Streptomyces globisporus WA5-2-37 from the intestinal tract of American cockroach (Periplanetaamericana). Front Microbiol 11:555
Shi Y-M, Bode HB (2018) Chemical language and warfare of bacterialnatural products in bacteria–nematode–insect interactions. Nat ProdRep 35(4):309–335
Ali SM, Siddiqui R, Ong S-K, Shah MR, Anwar A, Heard PJ, Khan NA(2017) Identification and characterization of antibacterial compound(s) of cockroaches (Periplaneta americana). Appl MicrobiolBiotechnol 101(1):253–286
Guzman Juan,Vilcinskas Andreas,Bacteria associated with cockroaches: health risk or biotechnological opportunity?[J] .Appl Microbiol Biotechnol, 2020
Rosas T, García-Ferris C, Domínguez-Santos R, Llop P, Latorre A, MoyaA (2018) Rifampicin treatment of Blattella germanica evidences afecal transmission route of their gut microbiota. FEMS MicrobiolEcol 94(2):fiy002

谷禾健康
焦虑,抑郁,双相情感障碍,这类词越来越多出现在我们的生活。
你也许不知道,饮食可以通过多种途径影响心理健康
即便知道了饮食干预可以改善抑郁,那么如何进行饮食干预呢?
饮食干预措施有好几大类,包括:营养干预措施(例如锌,omega-3脂肪酸);食品干预措施(例如绿茶,橄榄油);全饮食干预措施(例如地中海饮食)
什么样的饮食干预措施有效?
对哪些人有效?
在什么情况下有效?
……
要探索这些问题的答案
就需要了解支持这种关系的关键生物学机制。
将饮食与健康结果相关联的作用机制是复杂的,多方面的,相互作用的,并不局限于任何一种生物途径。
饮食可能影响心理和脑部健康的基本作用机制
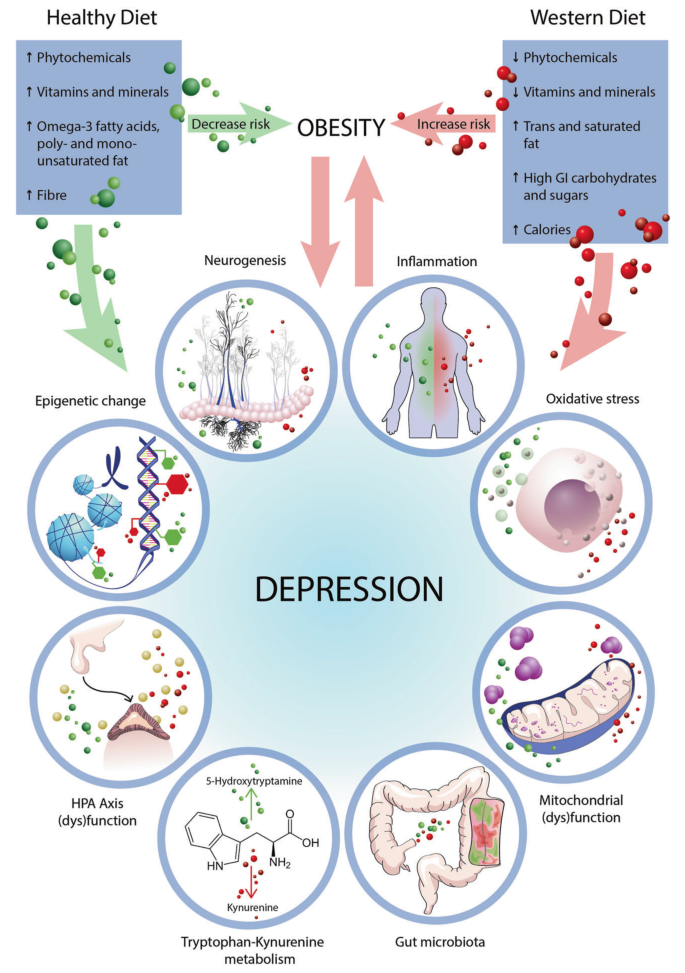
Wolfgang Marx,et al, Mol Psychiatry, 2020
有人说,抑郁和发炎有什么关系?
——大约25%的精神疾病患者(包括情绪障碍和精神分裂症)表现出较高的炎症水平。
炎症反应如何引起的?
免疫系统的这种过度激活是由多种因素引起的。生活中的困境,来自社会的压力,不良生活习惯(吸烟,缺少锻炼)等,都会引起炎症反应增加,从而促进抑郁症状。
典型的炎症反应包括三个主要成分:
炎症诱导剂(例如病原体或与受损相关的分子模式);
检测诱导物的传感器(例如免疫细胞表达的受体);
传感器诱导的炎症介质,包括细胞因子,趋化因子和前列腺素。
一旦被激活,这些炎症分子就可以影响与情绪障碍相关的生理领域,包括神经递质代谢,神经内分泌功能和大脑活动功能等。
抗炎剂可能辅助治疗抑郁症
用于医疗目的的细胞因子给药(如干扰素α注射)可引起情绪和行为的改变,例如情绪低落,疲劳,焦虑,睡眠障碍,快感缺乏和认知功能障碍,所有这些都与抑郁症的症状非常相似。Meta分析得出结论:抗炎剂,如细胞因子抑制剂、非甾体抗炎药和抗生素,包括米诺环素,可能是抑郁症的有效辅助治疗法。
健康饮食的抗炎特性
健康的饮食习惯(和单独的饮食成分)已显示出可能与精神疾病有关的抗炎特性。在慢性代谢疾病人群中进行的纵向观察研究和临床试验均表明,采用健康的饮食方式(例如地中海饮食)可以减少全身性炎症。
最近的研究还证实,患有严重精神疾病的个体比一般人群具有高得多的“饮食炎症”,即,促炎食物(如精制碳水化合物和反式脂肪)的摄入量较高,而抗炎食物的摄入量(主要来源于天然食品和植物)较低。
对纵向研究的Meta分析表明,具有炎性饮食模式的个体随着时间的推移发展为抑郁症的风险更大。
因此,促炎饮食改变为地中海或其他抗炎饮食模式,可能提供一种对抗与精神障碍的发作和严重程度相关的炎症状态的新策略。


抗炎食物
健康的饮食习惯会帮助摄入许多营养成分,可以减轻炎症。其中,蓝莓,可可和姜黄素等存在的植物化学物质(例如多酚)具有很强的抗炎特性,可能有助于治疗多种精神疾病。
Omega-3脂肪酸二十碳五烯酸和二十二碳六烯酸,多不饱和脂肪酸在鲑鱼等海洋食品中含量很高,具有抗炎特性,可以改善并延缓细胞因子诱发的抑郁症的发作。
氧化应激(氧化和抗氧化过程的不平衡)可能导致细胞对脂质,蛋白质和DNA的损伤。
持续的氧化应激是抑郁症和其他精神疾病的一种潜在的作用机制。
抑郁症氧化应激指标升高
对115项研究的Meta分析报告显示,与健康对照组相比,抑郁症患者的氧化应激指标(如丙二醛和8-F2-异前列腺素)升高,而抗氧化剂指标(如总抗氧化剂含量)较低。
此外,据报道抗抑郁药治疗后氧化应激标志物降低,证实了因果关系。一项验尸研究也显示,与健康对照相比,抑郁症、躁郁症和精神分裂症患者大脑中的氧化应激标志物升高。
除了氧化应激对细胞损伤的直接影响外,活性氧和氮的增加会导致线粒体功能障碍、炎症和色氨酸代谢改变,这些都与精神健康障碍有关。
西式饮食增加氧化应激的标志
饮食可以通过剥夺或增加具有抗氧化特性的饮食化合物的供应来加剧和改善氧化应激。动物研究表明,高脂肪西式饮食可以增加氧化应激的标志,如大脑和外周的蛋白质氧化和脂质过氧化反应。
提高膳食质量是可行的干预措施
营养丰富的饮食富含一系列具有直接和间接抗氧化特性的化合物,与氧化应激标志物(如f2 -异前列腺素和血浆氧化低密度脂蛋白)的降低有关。
维生素,例如抗坏血酸(维生素C)和α-生育酚(维生素E)具有直接清除自由基的特性。
营养素(例如硒,锌和半胱氨酸)是抗氧化剂系统(例如谷胱甘肽过氧化物酶和超氧化物歧化酶)的辅助因子。初步证据表明,补充抗氧化剂如n-乙酰半胱氨酸可以改善抑郁症状。
临床前研究表明,多酚还可以通过上调抗氧化防御系统来降低氧化应激,包括诱导核因子红细胞相关因子(Nrf)-2和调节炎症途径核因子κB(NFkB)和有丝分裂激活蛋白激酶(MAPK)。
肠道微生物群是饮食和大脑健康之间潜在的关键中介途径。
大量研究表明,肠道微生物群通过微生物群-肠-脑轴来调节生理过程,包括认知功能,神经精神疾病和行为。动物模型表明饮食、微生物群和与抑郁症相关的机制之间存在直接联系。
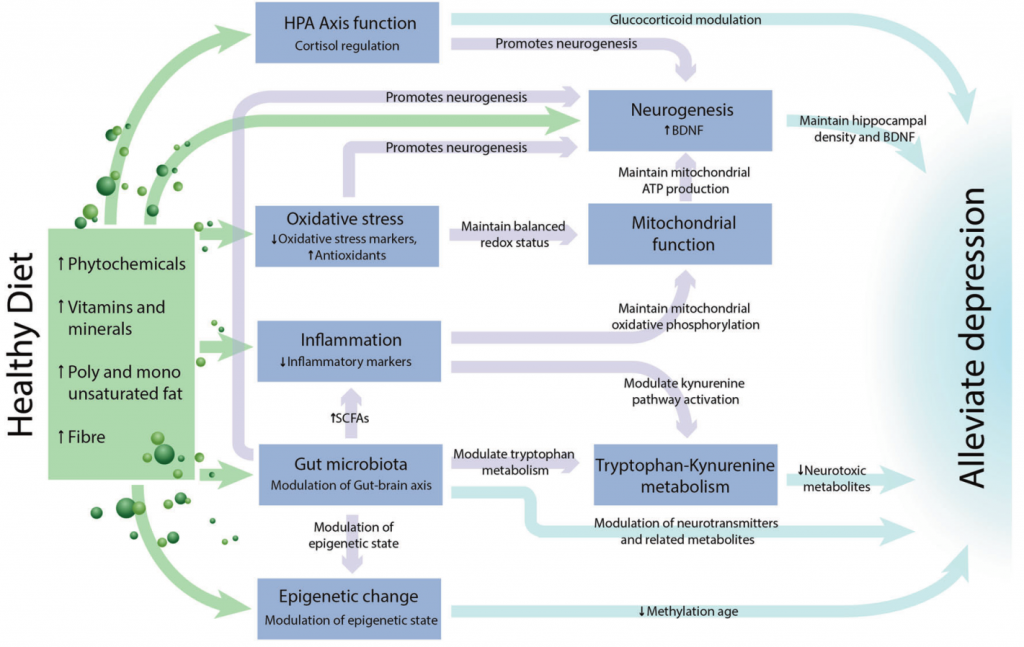

Wolfgang Marx,et al, Mol Psychiatry, 2020
高脂饮食导致焦虑样行为增加,记忆力下降
饮食引起的肠道微生物群的改变会导致行为改变,焦虑和抑郁。例如,在啮齿动物模型中,高脂肪西式饮食导致厚壁菌门/拟杆菌门比例增加,探索行为减少,焦虑样行为增加,记忆力下降。
其他临床前研究表明,高热量饮食会增加梭状芽孢杆菌 Clostridiales,疣微菌科 Ruminococcaceae 和拟杆菌目 Bacteroidales 的丰度,并导致认知灵活性较差,社会和物体识别能力受损。
益生元逆转慢性应激引起的肠道菌群变化
益生元补充剂(低聚果糖和低聚半乳糖)通过防止有益微生物(如双歧杆菌或乳杆菌)的减少,抑制正常的慢性应激诱导的促炎性细胞因子和小鼠的抑郁样行为的出现。
已经提出:肠道微生物群的多种直接和间接途径可以调节大脑功能和行为,包括微生物代谢产物(例如纤维细菌发酵产生的短链脂肪酸),神经元途径(例如迷走神经),神经活性途径(如5-羟色胺等神经递质和神经活性代谢物),下丘脑-垂体-肾上腺(HPA)轴,免疫和内分泌途径以及微生物群的直接神经活性代谢潜能。
饮食模式影响肠道菌群,影响情绪改变行为
短期营养摄入和长期饮食模式都是影响肠道菌群多样性,组成和代谢功能的因素。
迄今为止,只有一项不受控制的饮食干预研究的人类数据表明,富含菊粉的蔬菜饮食增加了双歧杆菌,导致饱腹感和个人能力水平的改善(但在情绪或感知压力方面没有差异)。

类似地,最近的一项研究表明,在老年参与者中,通过1年的地中海饮食干预,菌群变化与认知功能的改善和炎症标志物C反应蛋白和白细胞介素-17的减少有关。
营养素通过直接影响菌群调节大脑健康
例如,由肠道微生物群发酵膳食纤维产生的短链脂肪酸具有重要的免疫调节功能。这种关系也可能是双向的,肠道微生物群参与了这些化合物的生物利用度。
由此,我们可以想到,通过膳食补充剂(益生菌和益生元)和食物(例如发酵食品:泡菜,酸奶和酸菜等)来调节肠道菌群,以此作为调节菌群-肠-脑轴的一种手段。
益生菌和益生元
益生菌和益生元对精神健康影响的证据有限且变化很大。
研究表明,单独乳酸菌或与双歧杆菌联合使用——可能会改善抑郁和焦虑。
但也有Meta分析表明,与对照组相比,补充益生元后的抑郁或焦虑症状没有显著差异。然而,这是在大部分非临床参与者的有限样本(n = 4-5个)中进行的,一般来说,生物干预可能在临床参与者中显示疗效,而不是在非临床参与者中。
发酵食品是另一种具有操纵肠-脑交流的食品
研究表明,食用发酵食品后,有望改善情绪。由于益生菌的生存能力和可变定居能力,这可能解释了物种/菌株及其组合之间的效力不一致;包括各种植物食物来源的饮食模式,可能更有利于促进各种益生元底物和益生菌菌株消耗。

菌群通过食物过敏来介导饮食与大脑健康的联系
自我报告食物过敏在抑郁症患者中更为常见:
抑郁症vs.健康对照(13% vs. 9%)
真正的食物过敏,则饮食中的过敏原会触发胃肠道粘膜中肥大细胞的IgE致敏,导致一系列炎症介质,从而削弱肠道通透性。肠道通透性的提高与革兰氏阴性肠杆菌的易位性增强和免疫激活有关,可能导致全身性炎症,包括神经炎症,这是抑郁症的特征。
需要进一步对真正的食物过敏者进行大规模研究,以阐明其促进抑郁症的发展。对非IgE介导的食物超敏感(即食物不耐受)的研究(如麸质和酪蛋白),也可能揭示饮食诱导的肠道微环境变化如何影响情绪。
HPA轴由大脑(下丘脑),垂体和肾上腺组成,调节糖皮质激素的产生,并与精神疾病的病理生理有关。
超过60%的抑郁症患者表现出过多的皮质醇产生或对HPA系统产生其他干扰,例如对地塞米松抑制试验的反应改变和促肾上腺皮质激素水平降低。在临床恢复后,观察到HPA轴活性改变的一些指标的正常化,表明其在疾病病理生理学中发挥作用。
此外,儿童早期创伤可能会导致HPA轴永久失调,从而导致整个生命周期内出现精神疾病的风险增加。例如,与母亲分离的动物改变了HPA轴,从而表现出对成年期压力和记忆力障碍的反应。
营养素(例如维生素C)的临床干预试验报告显示,皮质醇对急性反应性降低健康成年人的生理压力。
Omega-3脂肪酸干预研究还表明,健康成年人和抑郁症患者的皮质醇水平都有改善。
同样,使用富含多酚的食物(如石榴汁和黑巧克力)进行的干预研究也报告了健康个体皮质醇水平的降低。


这种影响可能是通过心理应激后对下丘脑激活的促炎反应的调节而介导的。由于肠脑轴在心理健康中的新角色,益生菌也已被研究为针对HPA轴的潜在干预措施。
在健康成年人中进行的初步临床干预研究证实了这些结果。例如,在一项双盲,随机,对照试验中,与安慰剂相比,在健康个体中,多菌株益生菌干预24小时无尿皮质醇和自我报告的应激改善的结果。但在一项针对60名抑郁症患者的类似益生菌临床试验中,两组之间的血液皮质醇水平无显著差异。
成年海马体的神经发生水平与认知和情绪直接相关。
抑郁症患者的血清BDNF水平降低。
海马体是边缘系统的关键组成部分,在学习,记忆形成和情绪中起着核心作用。
啮齿类动物功能研究表明,成年海马体的神经发生水平与认知和情绪直接相关。例如,在小鼠中,海马神经发生水平增加与学习和记忆能力的改善有关,而海马神经元的减少通常与抑郁症某些方面行为有关。
BDNF(脑源性神经营养因子)是一种神经营养蛋白,在海马体中高度表达,并参与关键的细胞功能
确实,BDNF是典型的分子,可以解释饮食,运动和抗抑郁疗法对抑郁症和焦虑症行为的作用。重型抑郁症患者的血清BDNF水平降低,而BDNF对抑郁症发病机制的保护作用已获得了一些实验性支持。
证据表明,可以通过饮食调节BDNF和成人海马神经发生的调控。动物模型表明,高脂肪和高糖的西式饮食会损害神经发生并降低海马体内的BDNF水平,并对认知能力产生不利影响。
在动物模型中的大量研究表明,饮食成分(例如omega-3脂肪酸,益生菌和维生素)的有益作用。在精神病理,衰老和疾病的情况下,诸如白藜芦醇,蓝莓,绿茶,姜黄素和可可等多种多酚化合物,也已被证明可以逆转不良变化并保持成年海马神经发生的完整性。

此外,动物模型表明,其他饮食指标(包括卡路里摄入,进餐频率和进餐质地)可能会调节海马神经发生水平。
观察性研究提供了进一步的证据,表明健康的饮食习惯与较大的海马体积之间存在直接的联系,而不受多种解释因素(例如年龄,性别,教育程度)的影响。其他饮食范例,例如通过持续减少每日总食物摄入量或间歇性禁食(例如,每隔一天进食)来限制热量,也可能影响BDNF的表达。
最近的人类干预研究表明,西式饮食会损害海马依赖性学习和记忆能力。
当然,也可以通过本文中提到的其他途径,例如通过肠道菌群和炎症途径来调节神经发生。
色氨酸的主要生理途径是沿着犬尿氨酸途径,它会产生神经毒性的喹诺林酸和神经保护性的神经氨酸。
色氨酸是饮食中必须提供的一种必需氨基酸,是许多关键神经活性分子的重要组成部分。
在精神病学中,对色氨酸的可用性和代谢的关注主要集中在其转化为血清素,血清素是绝大多数抗抑郁药和一线抗焦虑药的治疗目标。
然而,色氨酸的主要生理途径是犬尿氨酸途径,它会产生神经毒性的喹诺林酸和神经保护性的神经氨酸。人们越来越多认识到导致犬尿氨酸产生增加的外周机制的重要性,并且认识到沿该途径产生的代谢物是一系列神经和精神疾病的重要神经生物学介质,包括但不限于抑郁症和精神分裂症。
此外,这种代谢级联反应的启动可能是由于压力或免疫系统和炎症途径的激活。这使得色氨酸在这一途径上的代谢可用性成为心理健康管理中的一个重要考虑因素。
我们日常生活中很多食物都含有色氨酸,包括鸡肉,金枪鱼,燕麦,花生,香蕉,牛奶,奶酪和巧克力。


尽管大部分来自摄入蛋白质的色氨酸都在小肠中吸收,但也还有大量可能到达结肠,肠道菌群在其命运和活动中起着关键作用。
在使用饮食干预措施进行心理健康预防和治疗的背景下,了解色氨酸的可用性和新陈代谢可能很重要。例如,增加的蛋白质摄入量可导致色氨酸的利用率增加,碳水化合物摄入量的变化会影响游离色氨酸的水平,非酯化脂肪酸可从生理上置换白蛋白中的色氨酸。
与色氨酸竞争通过血脑屏障运输的其他氨基酸的可用性波动也会影响中枢神经系统的代谢库。已经尝试了直接补充色氨酸作为抑郁症患者的一种干预措施,以改善血清素能信号传导。
还有一个研究调查了饮食干预通过其他方式调节尿氨酸代谢的作用,包括调节吲哚胺2,3双加氧酶(IDO)活性。体外和动物模型已经报道了个别的饮食成分,如姜黄素和绿茶,以及饮食方案,包括生酮饮食和禁食来调节尿氨酸途径的活性。
初步干预研究还表明,饮食方案,如热量限制和个人饮食成分,包括益生菌干预、白藜芦醇和红茶可能会调节尿氨酸代谢。例如,在最近一项针对60名抑郁症患者的试验中,与安慰剂相比,益生菌干预显著降低了尿氨酸水平,增加了3-羟基尿氨酸水平。
抑郁症的许多核心症状,如疲劳和认知障碍,都与中枢和外周线粒体功能障碍和生物发生减少相一致。
抑郁症和其他原发性精神疾病,包括双相情感障碍和精神分裂症一样,与线粒体功能障碍有关。
氧化磷酸化的破坏和线粒体ATP生成的受损可能导致神经可塑性失调和神经发生的减少,这两者都是抑郁症神经生物学的核心要素。
最近的一项研究显示,小鼠的线粒体移植恢复了海马体的ATP生成,逆转了脂多糖诱导的抑郁症模型,这一新证据支持了抑郁症的病理生理学因素。
大量临床前证据表明,不良饮食可能导致线粒体功能障碍。高脂饮食与线粒体生物发生异常有关,也与自由基生成增加、炎症和胰岛素抵抗有关。
高热量高碳水化合物饮食和高盐饮食有相似的作用,这些都是劣质西式饮食的核心成分。也有可能是由不良饮食引起的线粒体功能障碍的跨代遗传。在人类中,关于热量限制对线粒体功能的潜在有益影响,有不同的数据。
一些人类研究表明,限制热量的线粒体生物发生的标记物增加。
另一项研究显示,柠檬酸合成酶(一种线粒体含量的标志物)水平的提高和其他动物研究表明线粒体解偶联蛋白活性增强。到目前为止,还没有关于抑郁症患者热量限制的研究来测量线粒体功能障碍。


生酮饮食
有人提出了一种饮食模式来逆转线粒体功能障碍,特别是抑郁症患者从有氧到糖酵解能量的转变,就是生酮饮食。尽管在人类身上评估这一假设的临床试验仍在等待。生酮饮食增加线粒体解偶联蛋白的活性和水平。
注:生酮饮食(ketogenic-diet,简称KD)是一个超低碳,高脂肪饮食法。主要原理是用脂肪来替换碳水化合物,身体消耗完葡萄糖后,慢慢开始燃烧脂肪,给自己身体供能,身体慢慢进入生酮状态,最终,让身体进入一个高速燃脂的状态。
线粒体生物发生的改变在多大程度上介导了健康地中海饮食对抑郁症的有益影响尚不清楚。一些食物衍生物也有增加线粒体生物发生的假定作用,槲皮素,n -乙酰半胱氨酸和白藜芦醇都有一些支持的证据。
表观遗传过程可以影响DNA甲基化年龄,这与成年人的抑郁症有关以及许多其他神经发育结局和合并症,包括认知功能,酒精依赖,躁郁症和减少的海马体积,但不减少精神分裂症。
表观遗传学描述了在不改变基础DNA序列的情况下控制基因活性并使发育发生的分子机制。
极少有研究评估营养干预对甲基化年龄的影响,但已有研究发现其降低的证据。表观遗传状态受遗传序列,内部和外部环境以及发育过程中发生的随机过程的影响。产前发育,配子形成和青春期敏感时期的环境影响与患有与抑郁症有共同途径的慢性疾病风险有关,包括心脏代谢和神经发育障碍。这种现象被称为“健康与疾病的发展起源”(DOHaD)。
在DOHaD环境中,营养是对表观遗传学研究最多的环境影响因素。检验荷兰饥荒影响的研究表明,由于早期发育期间的营养不良,表观遗传失调与成人疾病风险有关。
很少有观察性的人类研究评估过表观遗传变化在介导早期营养对神经发育结局的影响中的作用,并且大多数是横断面的。最近的一项综述得出的结论是,有证据表明,某些早期营养摄入(例如母乳喂养和母亲肥胖)会影响表观遗传状态,进而可能介导儿童和青少年的心理病理学,例如内在化和外在化行为。
另外一项研究发现由于蛋白质和能量不足而在婴儿期住院的成年人在神经精神病风险基因中表现出DNA甲基化变化。体外细胞培养实验和啮齿动物研究表明,大量营养素的限制或过量存在对许多不同基因的多种表观遗传机制具有可再现的作用,包括与代谢和行为有关的基因。代谢扰动已成为基因组和表观基因组改变的驱动力,通过饮食,饮食中的影响得以保存在基因中。


营养丰富的饮食成分,包括叶酸,生物素,B6和B12等维生素;多酚,如姜黄素,白藜芦醇和染料木黄酮;脂肪酸和ω-3脂肪酸已通过多种机制影响表观遗传状态。此外,丁酸通常被认为是膳食纤维发酵过程中产生的有益微生物代谢产物,也可以影响宿主细胞的表观遗传状态。
饮食、情绪障碍和肥胖之间的多因素关系是双向和复杂的。Meta分析数据显示,肥胖症的男性和女性患抑郁症的风险增加55%,而抑郁症的个体患肥胖症的风险增加58%。
最近的一篇综述报道了饮食,情绪障碍和肥胖之间关系的几种相互关联的途径。途径包括HPA轴,其异常调节,过度活化以及糖皮质激素的过度合成和分泌与情绪障碍和肥胖症有关。
此外,研究表明接触高脂饮食(包括5-羟色胺和多巴胺)后,参与调节神经系统奖励回路,情绪和饮食摄入的各种神经递质水平降低。
为了减轻与压力有关的焦虑(由于被称为情感饮食和舒适食物的现象),长期压力和HPA轴过度活化可能导致过多食用西式食物和随后的肥胖。
在情绪障碍和肥胖症中都显示较高水平的炎症和相关细胞因子,提示其潜在病因之间存在另一个共同的联系。
在一项横断面研究中,肥胖在抑郁症与炎症标志物(即白介素6和C反应蛋白)之间的介导作用中,推测关系的因果关系是由抑郁症导致肥胖,炎症标志物升高。肥胖的这种发炎作用反过来推动体重增加和高复发率之间的关系。
不过这也说明了,限制热量和减肥饮食可能是减轻炎症状态和抑郁症状的可靠方法。同时,SMILES临床试验的结果表明,为期12周的地中海饮食干预对在没有体重变化的情况下降低临床抑郁症的症状。同样,前瞻性观察性研究反复报道了饮食质量与常见精神障碍之间的关联性证据,这些关联性与体重的测量无关。
性别差异也不能忽视
最近的一项Meta分析表明,饮食干预可能男性比女性受益。进一步的Meta分析显示,肥胖降低了男性患抑郁症的风险,而女性则增加了风险。可能有许多生物行为机制负责这种潜在的性别特异性效应,需要进一步研究。
首先,女性可能具有更大的改变脂肪或葡萄糖代谢的能力,以应对饮食干预。
其次, 男性在食物选择上更加以快乐为导向(这可能是由于多巴胺受体的差异所致),使得坚持健康饮食变得更加困难。
第三,男人更倾向于与男性气质相关的食物(例如红肉),而不是被认为更“女性化”的水果和蔬菜。
越来越多的证据支持饮食干预作为精神障碍的辅助治疗方法的潜在用途。本文主要讨论了饮食,肥胖和抑郁之间的相互作用(9种途径),当然饮食也可能通过其他慢性疾病影响抑郁,这些疾病通常与抑郁并存,包括糖尿病,代谢综合征和心血管疾病。
营养精神病学领域还处于萌芽状态。现有文献主要是临床前动物研究。还需要进一步的研究来确定可能影响饮食干预措施和饮食习惯的个体因素(例如年龄,BMI,共病),行为(例如改变的动机)和生物学(例如氧化应激,炎症)因素,可能影响治疗反应的因素等。
营养精神病学研究领域的扩展,使人们了解在什么情况下,哪类干预,适合什么人,为精神疾病患者制定新的针对性策略和临床指南,更加个性化地解决每个人的需求。
如果你深陷抑郁,请寻求专业治疗;
如果你朋友抑郁,那么请记得尊重。
相关阅读:
最新研究速递 | 柳叶刀:肠道微生物群在神经系统疾病中的作用
参考文献:
Lassale C, Batty GD, Baghdadli A, Jacka F, Sanchez-Villegas A,Kivimaki M, et al. Healthy dietary indices and risk of depressiveoutcomes: a systematic review and meta-analysis of observational studies. Mol Psychiatry. 2019;24:965–86.
Khalid S, Williams CM, Reynolds SA. Is there an associationbetween diet and depression in children and adolescents? Asystematic review. Br J Nutr. 2016;116:2097–108.
Borge TC, Aase H, Brantsæter AL, Biele G. The importance ofmaternal diet quality during pregnancy on cognitive and behavioural outcomes in children: a systematic review and metaanalysis. BMJ Open. 2017;7:e016777.
Hepgul N, Pariante CM, Baraldi S, Borsini A, Bufalino C,Russell A, et al. Depression and anxiety in patients receivinginterferon-alpha: the role of illness perceptions. J Health Psychol.2018;23:1405–14.
Köhler‐Forsberg O, Lydholm CN, Hjorthøj C, Nordentoft M,Mors O, Benros ME. Efficacy of anti‐inflammatory treatment onmajor depressive disorder or depressive symptoms: meta‐analysis of clinical trials. Acta Psychiatr Scand. 2019;139:404–19.
Rapaport MH, Nierenberg AA, Schettler PJ, Kinkead B, CardoosA, Walker R, et al. Inflammation as a predictive biomarker forresponse to omega-3 fatty acids in major depressive disorder: aproof of concept study. Mol Psychiatry. 2016;21:71–9.
Borsini A, Alboni S, Horowitz MA, Tojo LM, Cannazza G, SuKP, et al. Rescue of IL-1beta-induced reduction of human neurogenesis by omega-3 fatty acids and antidepressants. BrainBehav Immun. 2017;65:230–8.
Reichelt AC, Loughman A, Bernard A, Raipuria M, Abbott KN,Dachtler J, Van TT, Moore RJ. An intermittent hypercaloricdiet alters gut microbiota, prefrontal cortical gene expression andsocial behaviours in rats. Nutritional neuroscience.2020;23:613–27.
Long-Smith C, O’Riordan KJ, Clarke G, Stanton C, Dinan TG,Cryan JF. Microbiota-gut-brain axis: new therapeutic opportunities. Annual review of pharmacology and toxicology. 2020;60(Jan):477–502.
Marx Wolfgang,Lane Melissa,Hockey Meghan et al. Diet and depression: exploring the biological mechanisms of action.[J] .Mol Psychiatry, 2020
Liu RT, Walsh RF, Sheehan AE. Prebiotics and probiotics fordepression and anxiety: a systematic review and meta-analysis ofcontrolled clinical trials. Neuroscience & BiobehavioralReviews. 2019;102(Jul):13–23.71. Aslam H, Green J, Jacka FN, Collier F, Berk M, Pasco J,Dawson SL. Fermented foods, the gut and mental health: amechanistic overview with implications for depression andanxiety. Nutritional neuroscience. 2020;23(Sep):659–71.
Lerner BA, Green PH, Lebwohl B. Going against the grains:gluten-free diets in patients without celiac disease—worthwhileor not? Dig Dis Sci. 2019;64:1740–7.

谷禾健康
细胞间的代谢相互作用影响微生物群落的组成,从而影响它们在生态系统中的功能。众所周知,在交换代谢物的竞争下,浓度梯度限制了相互作用发生的距离。
然而,相互作用距离通常是在二维系统中量化的,或者不考虑竞争或其他代谢物的去除,这些条件可能与自然生态系统并不经常相匹配。
近日,来自荷兰阿姆斯特丹大学分子和生命科学研究所的一项研究题为“Microbial competition reduces metabolic interaction distances to thelow μm-range”分析了细胞间距离对三维水体系中交换代谢物竞争的单向交叉喂食的影响。
该研究利用反应扩散模型计算有效相互作用距离,并通过在不同空间结构中生长1μm大小的代谢物生成、受体和竞争细胞的合成联合体进行实验验证。

研究发现,接收器不能与距离它们平均15μm的生产者相互作用,因为产品浓度梯度在靠近生产者细胞的地方变平。因此开发了一个聚合协议,并改变了受体细胞的产物亲和力,以表明在生产者-受体聚集体中,即使是低亲和力的受体细胞也可以与生产者相互作用。
研究结果表明,竞争或其他代谢物去除三维系统中的公共物品可将代谢相互作用距离缩短至低1µm范围,这突出了浓度梯度作为细胞相互作用物理约束的重要性。
在致密的生物膜(细胞之间为0 µm)以及海洋(细胞之间> 100µµm)中都观察到了微生物的相互作用,这表明细胞以不同的距离相互作用。这些相互作用影响环境中的选择压力,因此影响微生物群落的结构和演化。从全球生物地球化学通量到人类健康,这些群落在许多生态系统中都发挥着重要作用,因此了解和控制这些相互作用非常重要。
参与相互作用的代谢物或信号分子可通过依赖于接触和不依赖于接触的转移机制进行交换。依赖于接触的机制需要较短的细胞间距离,并使用例如细胞,囊泡链或纳米管之间的直接接触进行交换。
与接触无关的机制要求将产生的化合物被动或主动转运到细胞外空间,然后通过扩散和对流运动。
与接触无关的相互作用可以是局部的(主要是在相邻细胞之间),也可以是整体的(在整个种群内),这取决于浓度梯度的分布。例如,酿酒酵母利用其胞外酶转化酶分裂蔗糖,从而在细胞周围形成葡萄糖和果糖梯度。在高蔗糖浓度下,聚集的酵母细胞和单个酵母细胞都可以生长(全局相互作用),而在低蔗糖浓度下,只有聚集的酵母细胞可以生长(局部相互作用)。
对于乳酸乳球菌的细胞外蛋白酶观察到相似的模式,其活性导致细胞周围的肽梯度。在高细胞密度下,蛋白酶阳性和蛋白酶阴性细胞都生长,而在低细胞密度下,主要是蛋白酶阳性细胞生长,因为只有它们才能从其产生的肽中受益。
接触无关的相互作用是局部的还是全局的,取决于细胞间的距离和浓度梯度分布,而浓度梯度分布受代谢物来源、代谢物汇以及它们之间的扩散和对流速率的影响。例如,胞外环境中的代谢产物可以是细胞外环境中的代谢产物。
代谢物库可以是代谢物消耗细胞、代谢物降解酶、液体流量或系统体积,因为稀释会降低代谢物浓度。虽然在这些研究中,源和汇的确切性质往往只是含蓄地提到,但它们的重要性是众所周知的。例如,当细胞彼此靠近时,代价高昂的合作交互作用更有可能进化,因为合作者与野生型非合作者竞争排出的代谢物。
因此,相互作用的选择通常是通过在琼脂平板上共培养细胞来完成的,并且还描述了相互作用的细胞进化出聚集表型。
这些示例表明,在存在代谢物去除池的情况下,浓度梯度会限制可能发生相互作用的距离。然而,尚不清楚这种相互作用发生在多远的距离。先前的研究要么在二维系统中量化了这些距离,要么没有去除代谢物的沉陷,而天然微生物群落则生活在三维环境中,竞争的代谢物消费者和其他类型的消费者很有可能存在去除代谢物的场所。
因此,我们将计算和实验分析相结合,以提供更系统和定量的视角,研究细胞间距离对代谢相互作用在三个维度上的影响以及存在代谢物去除池的影响。我们专注于葡萄糖在静态水系统中的扩散。
反应扩散模型和实验结果表明,在这些条件下,受体细胞不能与固定在15μm平均距离的葡萄糖生成细胞相互作用。然而,即使由于受体葡萄糖输入系统的遗传变异而导致受体细胞对产品的亲和力较低,生产者-受体的聚集也会促进代谢相互作用。
这些结果表明,对于糖,有机酸和氨基酸的竞争或三维水系统中其他代谢物的去除,可将代谢相互作用距离降低至低µm范围。
一个综合性联合体的设计与三维空间结构的生长
研究人员分析了细胞和细胞之间的相互作用的三维信息(表1.4)。为了在实验上验证模型结果,用四种乳酸乳杆菌菌株构建了合成联合体。
(1) 一种吸收乳糖并在细胞内水解成葡萄糖和半乳糖的“生产者”。它被设计成不代谢葡萄糖,因此当细胞生长在半乳糖上时,葡萄糖就会分泌出来。
(2)一种表达绿色荧光蛋白的“受体”,可以吸收葡萄糖而不是乳糖生长。由于受试者只能靠葡萄糖生长,所以他们的生长表明他们所在位置的葡萄糖可用性。
(3) 一种“非生产商”会吸收乳糖。它同时利用葡萄糖和半乳糖部分进行生长,因此不分泌葡萄糖。
(4) “竞争性葡萄糖消费者”(图2A)。
为了在三维系统中共同培养这些细胞,将葡萄糖产生者和受体(单向交叉喂食者)封装在平均直径为40μm的固化琼脂糖珠中。选择乳酸菌是因为与其他模型生物(例如大肠杆菌,酿酒酵母)它的新陈代谢和生物量产量对氧气的变化不敏感,它可以在这些琼脂糖珠中达到高细胞浓度。对于阴性对照,葡萄糖产生者被葡萄糖“非生产者”取代。
细胞以单独的细胞或聚集体嵌入珠子中。在培养过程中,琼脂糖珠用油或CDM分。油分离阻止了葡萄糖从珠子扩散,因此每个琼脂糖珠作为一个单独的隔间。
总体而言,数据表明,在具有代谢物消耗池的三维系统中,可获得陡峭的浓度梯度,并且彼此之间平均相距15μm的细胞无法通过葡萄糖交叉补给进行相互作用。可以通过将细胞聚集在低µm范围内来克服这种物理限制,如通过细胞聚集(物理接触)所实现的。
在存在代谢物-水槽的情况下,涉及扩散系数与葡萄糖相似的分子的相互作用(例如其他糖,有机酸,氨基酸)要求细胞间距离在低µm范围内。一致地,许多细胞外底物降解酶附着在细胞上,从而使源(酶)靠近受体(细胞)。
转化酶例如位于酿酒酵母的周质空间中,乳酸乳球菌的蛋白酶附着在细胞壁上,在真菌和细菌中,纤维素体也附着在细胞壁上。
Hauert等认为,当生产者也从自己的产品中受益时(例如细胞外酶),只有当酶的生产成本很高时,细胞的空间结构定位才是有利的。
因此,细胞外酶在细胞壁上的附着表明这些酶是昂贵的。这与巴赫曼(Bachmann)等人的观点一致,他在乳酸乳球菌中显示,蛋白酶阴性菌株与细胞壁结合的蛋白酶的竞争优于蛋白酶阳性菌株,除非它们的间隔大于1mm(细胞密度低于103个细胞/ mL)。
图一:二维和三维反应扩散系统中的预测浓度梯度
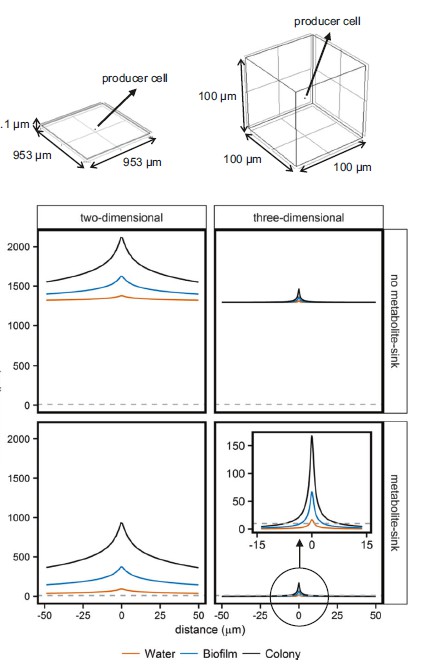

在存在和不存在代谢物池的情况下,将产生葡萄糖的细胞置于二维或三维空间中。通过改变扩散系数模拟了不同的环境。葡萄糖在水中的扩散系数(Ds)设置为6.7×10-10,生物膜中的葡萄糖扩散系数(Deff,biofilm,s)设置为Ds的0.25倍,并且扩散将菌落中的葡萄糖系数(Deff,biofilm,s)设置为Ds的0.10倍。
在COMSOL Multiphysics中,一项与时间有关的研究在数分钟内产生了浓度梯度该图显示了孵育5小时后穿过生产细胞的水平线上的浓度。
为了提高可见度,两个图都将x轴范围设为相似,因此对于二维系统,仅显示了部分浓度梯度曲线。水平虚线表示浓度为10μM。
图二:三维空间结构环境中的代谢相互作用
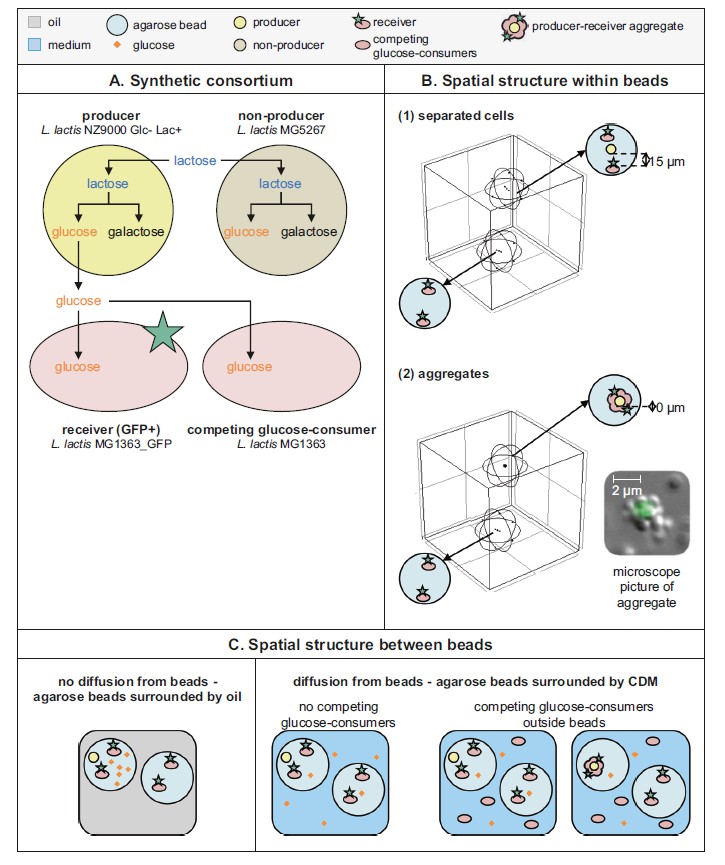
A 用于合成合成菌群的四种乳酸乳球菌菌株:
(1)吸收乳糖并分泌葡萄糖的“生产者”
(2)吸收葡萄糖并表达GFP的“接收者”
(3)“非生产者” “摄取乳糖但不分泌葡萄糖的”和“竞争葡萄糖的消费者”
(4)摄取葡萄糖(补充信息第2节,图S3)。
B 琼脂糖珠内的三维空间结构。
细胞之间15μm的距离相当于3×108细菌/ mL的均匀分布。显微图像显示聚集体,出于可见性原因,表达GFP的细胞被非荧光细胞包围。实验中使用的聚集体形成相反的方向:(非)生产者细胞被表达GFP的受体包围。
C 琼脂糖珠之间的三维空间结构。
聚集体仅在存在竞争性葡萄糖消耗剂的情况下孵育。
图3:葡萄糖转运蛋白亲和力和Vmax对受体反应的影响。
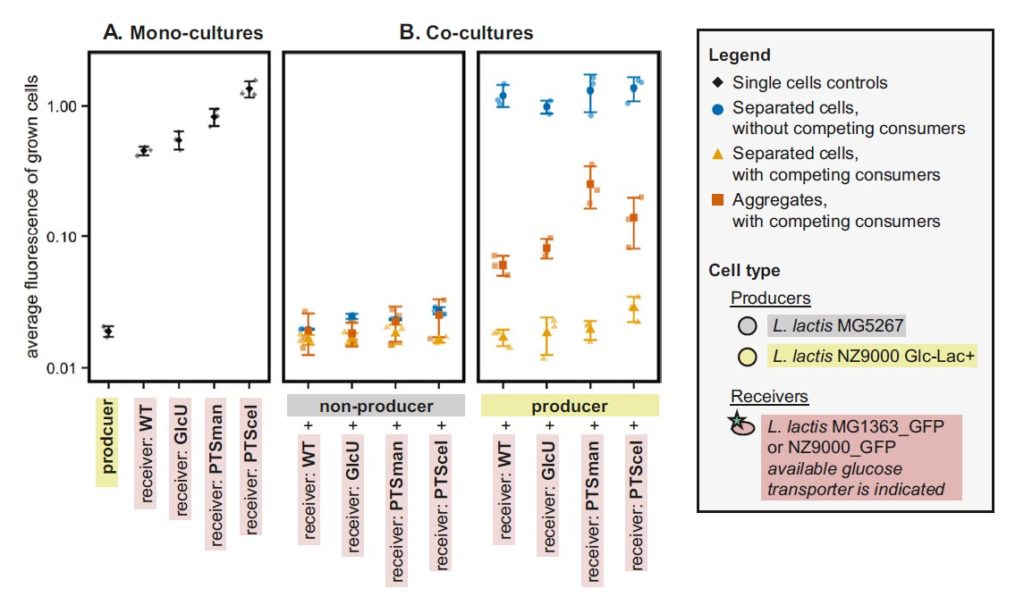
较大的化合物(如酶)具有较低的扩散系数,预计在存在代谢物汇的情况下,相互作用距离会增加。但是,这里的浓度梯度也会影响相互作用中涉及的分子机制的演变。
大型聚集性资源(如颗粒铁(> 0.4µm))的缓慢扩散会导致例如细胞内铁的吸收受到限制。
因此,可以推测细胞通过分泌与铁颗粒结合的铁载体,提取铁离子并随后形成快速扩散的铁-铁载体络合物来提高铁的吸收速率。
代谢物的交换是活细胞的关键特征,是细胞外营养物质代谢和与其他细胞相互作用的基础。这项研究指出了浓度梯度可能会影响细胞相互作用的限制因素和机会,以及会如何影响它们的进化及其在微生物聚生体中的作用,这些为未来研究人员理解和指导微生物的互作提供了证据视角。
参考文献:
van Tatenhove-Pel Rinke J,Rijavec Tomaž,Lapanje Aleš et al. Microbial competition reduces metabolic interaction distances to the low µm-range.[J] .ISME J, 2020, undefined: undefined.
相关阅读:

谷禾健康
识别和纠正疾病患者微生物组的重要第一步,就是要了解健康微生物组的特性,及没有明显疾病情况下的许多不同的微生物生态。
我们知道体内DNA的细微差异引起巨大的表型多样性,也就是说大家都能区分彼此。相比之下,人体微生物的宏基因组(微生物在我们体内的总DNA含量)变化更大,只有三分之一的组成基因存在于大多数健康个体中。所以要理解健康微生物组这些差异是重大的挑战。
本文,我们将对以下方面展开讨论分析:
人类对健康人微生物组的理解史;
健康人肠道微生物群的组成和功能;
健康人微生物多样性的范围;
可能的驱动因素,如地理,饮食和生活方式等;
健康微生物群的核心标志;
微生物群的抵抗力,弹性和稳定性;
有助于人类健康和疾病预防的微生物代谢途径和种类;
现代农业与人类健康肠道微生物。
“健康” 是指没有任何明显的疾病。
“微生物组” 指人体的整个微生物组,但目前大部分可获得的数据描述的都是肠道微生物组,因此这里讨论的许多发现都来自肠道微生物组。
关于健康微生物组,有两种假设:
第一种假设
试图确定在健康个体中普遍存在一组“核心”微生物群,其假设是不存在此类微生物就表明失调。但健康个体之间的生态多样性研究表明,健康人的微生物组本来就有足够的差异,无法支持这样的假设。
第二种假设
健康的“功能核心”:一种代谢和其他分子功能的补充,由特定栖息地内的微生物群执行,但不一定由不同人体内相同的生物体提供。
这样的核心:
可能要作为遗传潜能存在,就像人类基因组一样必须严格,不能编码出严重的有害突变才能健康;
或者可能要在个体内部有良好的表达和调控,以保持健康(也就是说,它必须由RNA转录组编码或以蛋白质或小分子产物的形式存在);
当然也可能是这两者组合。
当然,功能核心:
必须至少包括个体微生物生命所必需的内部管理功能,这些功能必须通过基因组正确表达;
还可能包括特定于人类生态系统中微生物生态位的功能;
可能包括人类细胞无法完成的过程,因此代表了共生宿主与微生物关系的潜在基础。
核心功能的适应性
健康的微生物组可能会进一步通过其随时间变化表现出来。直白地说,一个健康相关的微生物群必须对外界或内部变化有一定程度的适应性。外界例如饮食或药物等干扰,内部变化例如年龄的改变。
即使某个特定的群落结构提供了所有必要的核心功能,但如果没有这种适应性,就不能长期保证核心功能。
因此,微生物组对压力和干扰的抵抗力及其之后恢复健康功能的能力是表征健康微生物组的潜在特性之一。
早期的研究试图通过培养和表征菌的生理特性来确定正常的微生物群,主要在肠道中。这类研究最能突出在实验室环境中生长良好的生物,例如大肠杆菌。这种偏见导致人们认为大肠杆菌是人类肠道微生物组中丰富而普遍的成员。
在1970年代引入了严格的厌氧技术,仅肠道就可以回收300多种细菌。此外,对选择性培养基中标准系列稀释液中活细胞的计数可以对这些物种进行定量。
对该时代的四项大型研究的总结,研究了141名美国人在不同饮食下的粪便样本,发现拟杆菌(Bacteroides)和厌氧球菌(anaerobic cocci)属细菌普遍存在且数量丰富,而梭菌属细菌则普遍存在,但丰度较低,尽管没有在所有受试者中均观察到单一物种。其他普遍但含量较低的细菌包括双歧杆菌属(Bifidobacterium),真细菌属(Eubacterium),乳杆菌属(Lactobacillus)和链球菌属(Streptococcus),以及兼性厌氧菌如大肠埃希氏菌(Escherichia)。
当时已经有人怀疑,还有大量与人类有关的微生物种类没有被发现,一项研究估计在一个健康的结肠中同时存在大约400种微生物。然而,发现它们也有很多障碍,比如某些微生物的培养要求严格,以及培养所需的劳动密集型性质等。
此外,并不是所有的微生物都可以通过在选择性培养基上单独培养而被很好地区分为物种或菌株。例如,不同种类的高产拟杆菌属特别难以解开。另外,这种关于群落组成的研究甚至更难扩展到非细菌性微生物,例如病毒和真菌,对于微生物含量低于肠道的人体栖息地的研究更加不切实际。需要新方法来研究健康微生物组的这些方面。
DNA测序和荧光原位杂交(FISH)等与传统培养无关的技术现已广泛使用,它们的民主化使微生物样品的DNA含量得以直接研究。使用针对16S核糖体RNA基因的FISH进行的早期研究表明,在西欧队列中,至少三分之二的肠道细菌可归因于大约六个物种/属水平的一组六组细菌:两个拟杆菌,两个梭菌,链球菌 /乳球菌和直肠细菌。从那以后,这被证明是乐观的,甚至在当时,这些样本之间的丰度也观察到了很大的差异(标准偏差为均值的〜60–80%)。
某些直接从样品中直接测序16S rRNA基因的早期努力表明,对应于已知物种的85%至95%的细菌丰度可归因于与拟杆菌,梭状芽孢杆菌XIVa和梭状芽孢杆菌第四类有关的三个细菌群。16S研究还显示,健康人之间以及一个人内紧密联系的生物地理位点(例如粘膜和粪便样本)之间的生物分类组成存在很大差异。然而,在所有这些研究中,当时的大多数(75–80%)序列簇与任何文献记载的物种都不匹配,这解释了以前工作中对多样性的低估很多。
随着数据库的完善和技术的进步,目前16s尤其是在肠道菌群方面分辨率也已经达到很高了,种方面能达到70%以上,40%左右能分到菌株。而且肠道菌群检测远不是检出了每种菌的含量,还需要大规模人群队列和疾病样本的训练以及模型构建,因此16s有其标准化和相对适合的成本,在肠道菌群中还是首选的方案。
此外,大规模并行shotgun测序(高通量测序技术)的出现已基本解决了这种微生物“暗物质”的分类学组成,虽然功能多样性的显著百分比(高达50%)和非参考群体的组成仍有待确定。
最初的发现印证了人与人之间的巨大差异(甚至在双胞胎之间),但也暗示了存在着所有个体共有的一组微生物基因。 这有助于建立这样一种模式:就好比在每个生物体中保存的管家基因一样,“核心微生物群”可以在功能层面而不是在分类学层面上被定义。
人类和人类文明有上千年的变化,人类肠道微生物群也随着饮食的变化而变化。例如,狩猎-采集社会的肠道微生物群落在一年中显示出剧烈的变化,反映了食物供应的变化。此外,在这些社会中,女性和男性成员的微生物群也存在重大差异,女性的微生物群更像食草动物,而男性成员的微生物群更像食肉动物。
从早期文明到现代文明,肠道微生物群的变化也反映了卫生的变化,这在城市和农村社区之间仍然存在。
在过去的几十年里,随着卫生条件的改善、加工食品和药物(尤其是抗生素)的广泛使用,现代生活方式似乎对人类肠道微生物群落的多样性产生了重大影响,总体上减少了其多样性。
人们吃的东西对肠道微生物组成的影响要比基因强得多。设想这样两种情况:
没有血缘关系却生活在一起的人(有相似的饮食习惯和生活方式);
生活在不同地方的有血缘关系的人;
后者的微生物组表现出更大的差异。
此后,在测序和其他分子测定法的通量和成本效益仍在不断提高的基础上,开展了大规模项目,以表征微生物成分的多样性及其功能潜力。
2010年,人体肠道基因组(MetaHIT)研究报告了来自124个欧洲成年人(主要是“健康”人群)粪便样本的肠道基因组,当时该序列超过了以往所有微生物组研究的测序量近200倍。
2012年,人类微生物组计划(HMP)报告了对242位来自美国的健康成年人进行16S谱分析的结果,并对139个人的亚组进行了宏基因组测序,结果代表了分布在五个主要身体区域之间的18个身体栖息地。一项针对2型糖尿病的大型中国研究很快贡献了145个肠道宏基因组,其中大约一半来自非糖尿病对照组。此后,MetaHIT协会继续发布来自欧洲成年人的新的肠道宏基因组。
人类微生物组群项目(HMP)和人类肠道宏基因组学(MetaHIT)倡议是第一个定义健康人体微生物组组成和功能的大型微生物组项目。这些努力为了解宿主相关微生物群落的生理和功能特性提供了基础。
随着新一代测序、全基因组鸟枪测序、全球代谢组学、先进的计算策略以及人性化动物模型和基于培养的人类类器官系统等新技术的出现,对微生物组的理解正在迅速进步。
健康的肠道菌群因为个体差异较大,很难有一个精确的定义,但通过大量人群样本数据的了解我们可以明确什么样的肠道菌群是不健康的。
# 多样性
首先从多样性的角度,一般除了2岁以下婴幼儿,大部分的健康人的肠道菌群构成大概有200~2000种菌,如果数量过低或过高都不算健康。过低表明饮食结构过于单一或经历了抗生素等处理。过高表明可能出现了很多不属于肠道环境的菌。
# 核心肠道菌群的占比
另外主要从核心肠道菌群的占比来评估。健康的肠道菌群虽然构成多样,但是存在一部分核心的肠道菌群,它们是肠道菌群的基石,如果这些菌的占比不足30%表示肠道菌群环境已不由这些健康菌占主导,很可能为病原菌提供生存环境。
这些核心的肠道菌群包括:
拟杆菌属(Bacteroides),
柔嫩梭菌属 (Faecalibacterium),
普雷沃氏菌属 (Prevotella),
瘤胃球菌属 (Ruminococcus);
此外, 粪球菌属(Coprococcus),Blautia,
Dorea,毛螺菌属(Lachnospira),
罗氏菌属(Roseburia)也属于常见菌属,但占比通常不会很高。
在婴幼儿的肠道菌群中核心菌包括:
双歧杆菌属(Bifidobacterium),韦荣氏球菌属(Veillonella),通常2岁以下婴幼儿的肠道菌群这两种菌一般至少占比30%以上。
如果你的肠道菌群构成中基本见不到这些核心菌或占比很小,那么肠道菌群很可能已经出现问题。
古细菌,病毒,真菌和其他真核生物
古细菌
人类微生物组除了我们关注最多的细菌外,还包括古细菌,病毒和真核生物。在健康的人类微生物组(主要是肠道)中,已鉴定出少量古细菌属。其中产甲烷菌属在肠道中最多,特别是史密斯甲烷短杆菌(Methanobrevibacter smithii)非常适合人类肠道,可优化其他微生物对膳食多糖的消化,并在常见肠道细菌(如多形拟杆菌,Bacteroides thetaiotaomicron)存在的情况下适应其基因表达。
病毒
人类的病毒特别广泛是健康人类生态系统不可或缺的一部分。病毒有高变特性,因此每个人都有一个独特的病毒,主要由噬菌体组成(估计有5%的肠道细菌基因编码原噬菌体蛋白)。噬菌体还提供了另一种途径,在其他亲缘较远的细菌之间进行水平基因转移。由于古细菌,病毒和真核生物的分子谱分析技术仍不及细菌,这些生物的分子功能信息仍然有限。
真核微生物
在人体中发现的最著名的真核微生物(主要是真菌和原生生物)虽然通常是致病菌,但此类真核生物在健康人群中也普遍存在,特别是念珠菌(Candida),马拉色菌(Malassezia) 和Saccharomyces。
不同部位的相互作用至少负责健康微生物组的部分生态和免疫平衡。例如,在皮肤生化环境中细菌和真菌之间存在明显的竞争,或是乳酸菌控制肠道和阴道真菌。
人类与真菌之间存在直接的相互关系:其中最典型的特征是益生菌(Saccharomyces boulardii),最初是为了对抗霍乱而分离的。一些原生动物甚至是健康微生物群落的常见物种,变异比细菌更大。此外,某些原生动物的存在,例如常见的芽囊原虫属Blastocystis,与降低胃肠道疾病的风险有关。
健康微生物组的地理变异
对比来自不同国家的肠道微生物组的研究,已经确定了微生物组成的系统差异。比较欧洲的MetaHIT,美国的HMP和中国的糖尿病队列这三大块,发现国家间的生物分类组成差异大大超过了个体间的差异,这不止是由于技术差异或者实验方法造成的,地理学因素也是微生物组大规模变化的原因之一,包括北美和南美,欧洲和非洲,韩国和日本以及俄罗斯和中国的农村和城市人口之间的差异。
在这种变化的可能驱动因素中,主要因素是饮食,以及其他因素,包括地理,早期生命暴露和遗传等。还没有一项研究表明健康微生物群大量种群间差异的原因是这些因素其中的任何一个。
有意思的是菌株水平上也存在地理差异,特别是当菌株特征比微生物丰度曲线表现出更大的时间稳定性。该领域的研究是初步的,但表明国家或大洲之间的菌株差异不是特别明显。诸如拟杆菌属(Bacteroides coprocola)和普雷沃氏菌(Privotella copri)之类的物种在跨越人群的抗生素抗性基因中表现出最大的差异和菌株水平变异。
虽然在所有地点的微生物组的分类组成中有巨大的个体间差异,但在一个特定地点,个体间代谢途径的丰富程度相当一致。
此外,微生物组的组成在生命的最初几年发生了巨大的变化,但这种功能性特征在生命早期就已经建立,并在此后保持稳定,至少在肠道。这表明,“核心”健康微生物群的一个定义是,它们的组合、代谢模块和调节途径共同促进了与宿主相关的稳定生态。
这个核心包括至少三类功能
第一,也是最简单的,所有微生物生命所必需的管家功能,例如转录和翻译、能量生产和结构成分。
第二,这一核心包括人体相关的微生物群在身体部位的特定过程,如与宿主细胞表面的黏附,以及与宿主与微生物相互作用有关的化合物的产生(包括必需的维生素,如维生素K和免疫刺激化合物)。
第三,不同的身体栖息地各有其特殊的核心功能。例如,在肠道中,核心功能包括糖胺聚糖的生物降解、几种短链脂肪酸的产生、特定脂多糖的富集以及维生素和必需氨基酸的产生。在一个特定的人群中,哪种功能趋于丰富,会受到长期的选择压力(如饮食)的影响。
健康微生物群的另一个标志是生态系统的恢复力。
从微生物生态学角度来看,健康的其他标志是抵抗干扰(可能是由于病原体进入,饮食或药物治疗而引起)以及随后恢复健康状态的能力。这些特性分别被称为阻力和回弹力。
系统在冲击(阻力)期间持续存在的能力以及在干扰影响(恢复)后恢复到基线的能力决定了整体的弹性。从概念上讲,下图对此进行了说明。
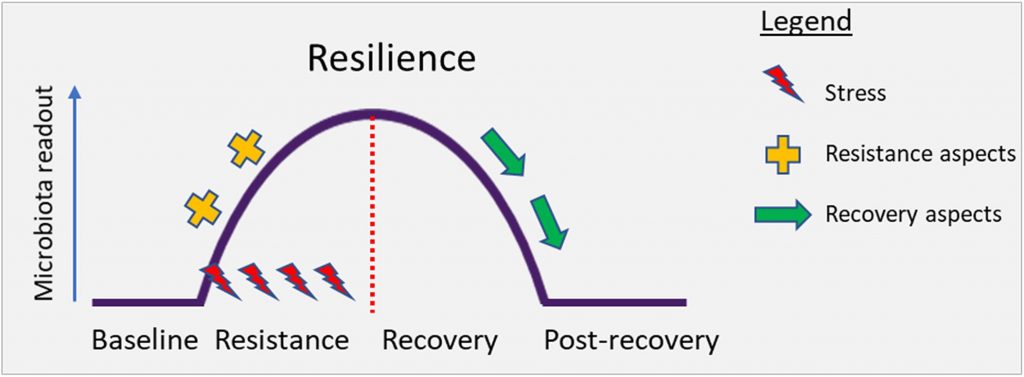

S. K. Dogra et al., Front. Microbiol.2020
例如,经过抗生素治疗后,健康的肠道菌群通常会在几周到几个月后恢复到以前的状态。因此,微生物健康的最新定义明确地不包括单一静态,而是动态平衡。
原生状态、扰动、回到原始状态或过渡到新状态概念图解
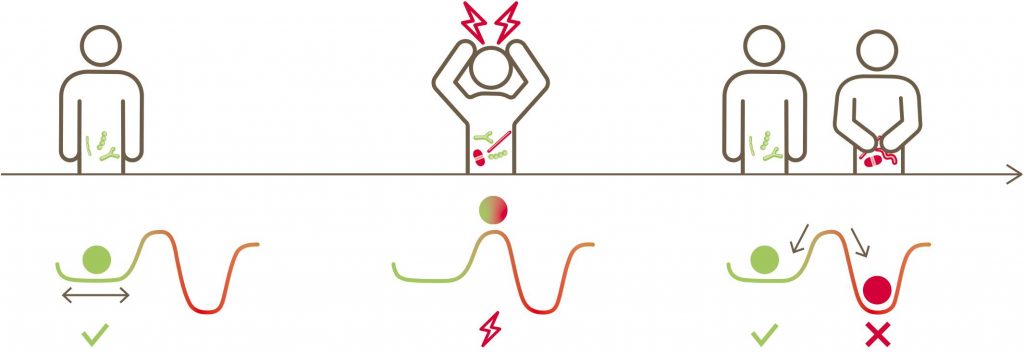

S. K. Dogra et al., Front. Microbiol.2020
一个有弹性的微生物群在受到扰动后将恢复到原来的平衡状态,而非弹性微生物群将转向一个改变后的新状态。
在这种观点下,健康的微生物组相当于潜在动力系统的吸引子。吸引子同时捕获抵抗力和弹性,因为系统将抵抗吸引者的偏离,并且除非波动(可能是由于外部扰动或内部随机性引起的)足够大,否则它将倾向于返回到稳态区域。
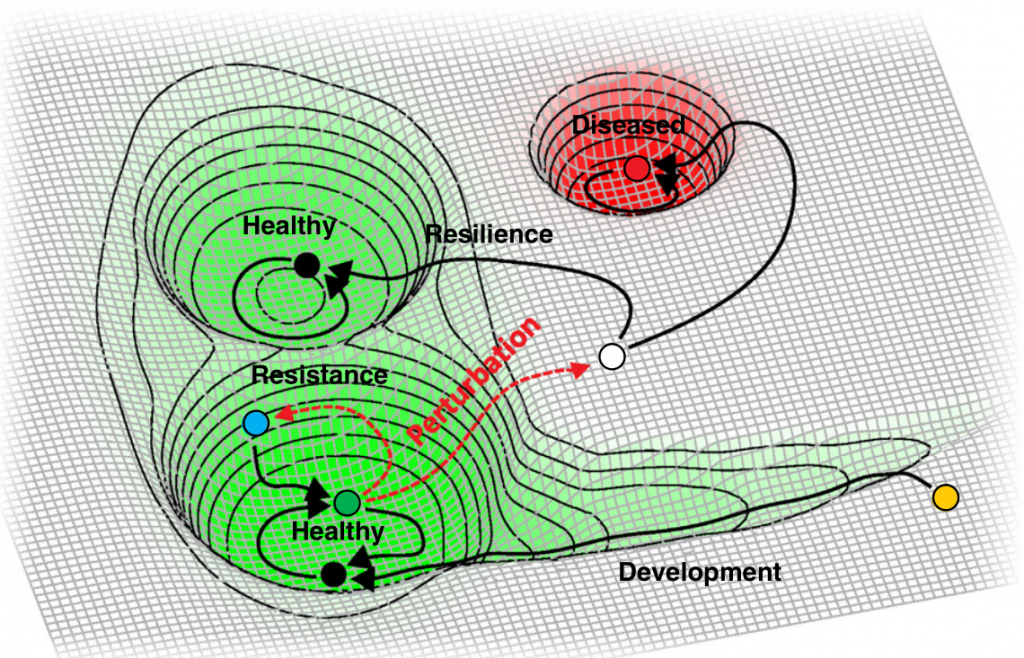

Lloyd-Price et al. Genome Medicine (2016)
人类微生物组中最明显的例子可能是健康阴道中的群落状态类型之间的转换。不是所有菌群类型都具有相同的稳定性。肠道微生物组也在不断变化,随着时间的流逝而增加和减少物种,不同的分类单元具有不同的稳定性,并且某些微生物在肠道中持续存在多年。
特定分类群持续存在的机制还不明确,但是有趣的是推测这种机制是否可能与微生物组组装背后的驱动原理有关。如果特定的群落确实主要是为了组装来填补一组适合栖息地的功能生态位,那么在特定的组装中提供关键的代谢,信号传导,免疫调节或其他作用的物种可能比功能外围的物种在时间上更稳定。
因此,动力学与微生物组的分类学多样性和巨大的分子功能潜力之间的耦合,提醒了人类微生物组的复杂性,因此也难以定义简单的微生物健康概念。
植物和人类微生物群是相互关联的。
健康土壤,健康植物与健康肠道微生物
人类的肠道微生物群和植物的土壤和根茎微生物群在相似的环境条件下存在
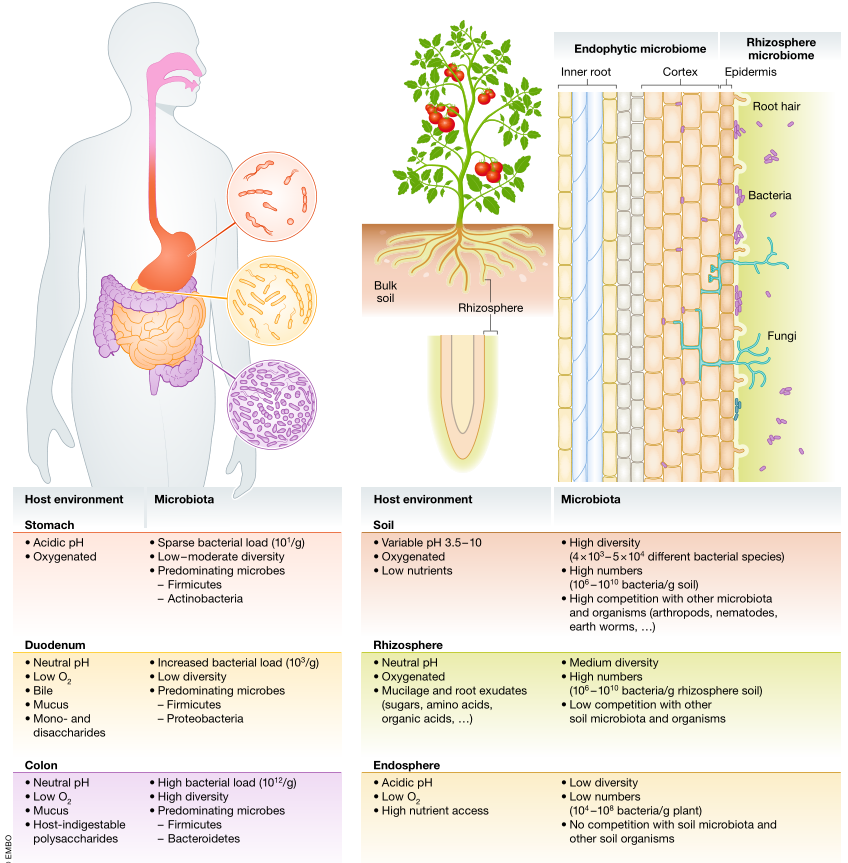
Heribert Hirt, EMBO Reports (2020)
最近的研究表明,根部和肠道微生物群落的存在条件相似。
两者都是开放系统,其特征在于氧气,水和pH的梯度会产生多种不同的生态位。
两种系统都从环境中继承其微生物物种:分别是人类的食物和植物的土壤。植物和肠道系统由大量相似的细菌门组成(Firmicutes,Bacteroidetes,Proteobacteria,Actinobacteria)
与人类粪便转移类似,从抑制疾病的土壤中移植有益微生物可以保护植物免受各种疾病的侵害。
对不同的哺乳动物食草动物和食肉动物的研究表明,肠道微生物组从食用原始植物材料中吸收纳入了一些成员。
根与肠微生物合成必需的氨基酸,维生素和许多其他调节其宿主免疫系统的次生代谢产物。
因此,植物和肠微生物群可以被视为对宿主健康至关重要的代谢器官。
植物微生物群对人体肠道微生物群的直接和间接影响
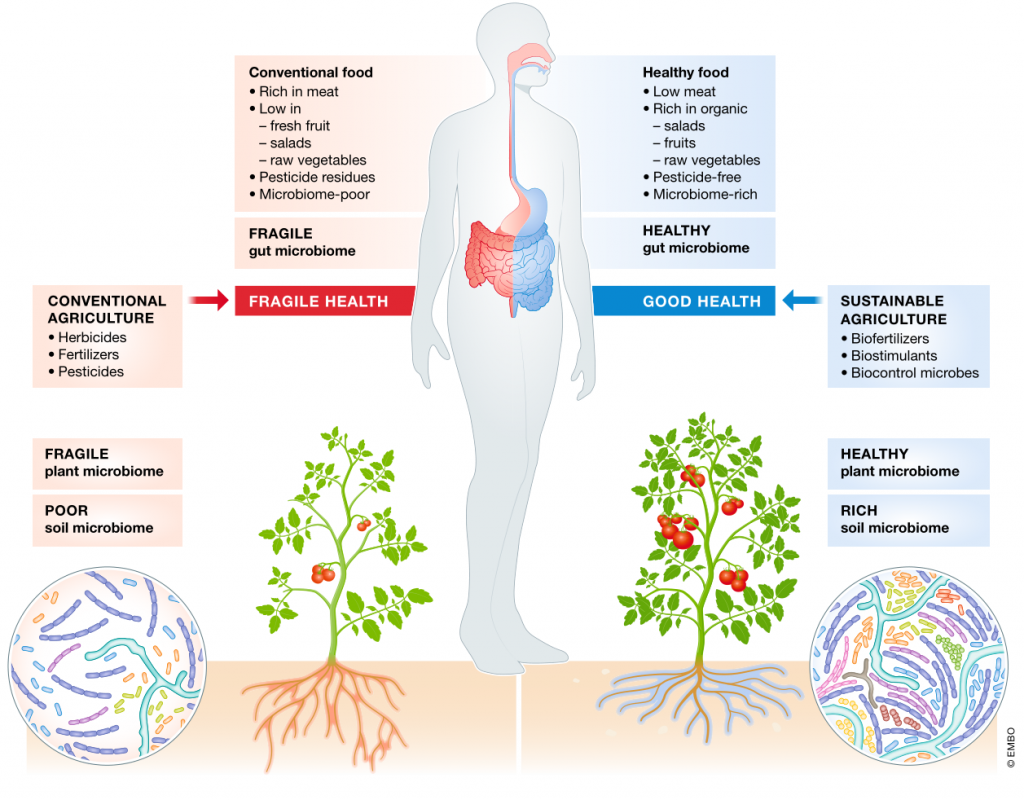

Heribert Hirt, EMBO Reports (2020)
饮食与微生物组
西方饮食中肉类消费量增加,蛋白质的高输入也极大地影响了肠道微生物组,某些微生物会抑制有益菌并改变我们的饮食习惯,以偏爱更不健康的食物。
人类少数必需化合物是由微生物产生的,微生物本身是必需氨基酸和维生素的重要生产者。例如,钴胺素(维生素B12)不能由植物或动物产生;它是由植物微生物群或反刍动物肠道中的微生物合成的。
土壤、气候对菌群的影响
土壤侵蚀和气候变化也影响微生物多样性,并导致大面积耕地及其微生物群的损失。因此,今天的作物缺乏许多重要的共生菌来生产或增加维生素、矿物质、抗氧化剂和其他对植物和人类健康有益的代谢物。
农药对菌群的影响
大量研究表明许多常用农药对人体健康的负面影响。例如,大多数铜基杀真菌剂对固氮细菌具有有害作用。
许多新鲜水果,蔬菜长期保存和运输用到农药,这些化学物质不仅会通过食物进入人体肠道,还会杀死植物微生物。
除草剂——草甘膦杀灭杂草,确保作物高产。然而草甘膦对人类的急性毒性较低,但草甘膦对于人类而言是潜在的致癌物质。草甘膦对土壤、植物和肠道有益微生物的抑制浓度远低于致病微生物。就人体肠道微生物组而言,对有益菌双歧杆菌(Bifidobacterium sp.)以及肠球菌属的抑制作用比致病菌梭状芽孢杆菌和沙门氏菌的抑制作用大。
农业抗生素影响微生物组
农业使用的抗生素大约是人类医学的四倍。
在农场中滥用抗生素来促进牲畜的生长,这极大地促进了耐药细菌的出现;
动物排出的抗生素会改变微生物的功能和土壤,水道及其他生物群落的组成;
食用抗生素处理过的动物的肥料施肥的田地上的新鲜农产品的消费可以将抗性基因传播到人类肠道微生物组,并进一步出现具有多重耐药性的人类病原体。
人体胃肠道微生物组对于维持人体健康至关重要。鉴于健康个体中微生物群落结构的复杂性和多样性,目前可能难以界定基线“健康”微生物群的特征属性。随着人类微生物组研究取得了技术进步,需要利用大样本人群来确定这个边界以及哪些关键因素会影响和塑造肠道微生物。重要的是更好地了解数万种不同微生物物种在与宿主之间的复杂相互作用网络中的功能和作用。
此外,减少肥料,杀虫剂和除草剂的使用量提供健康的绿色食品有助于人类建立和维持健康的肠道微生物组。因此,有关微生物群落对宿主代谢和健康的整体作用的研究不应仅仅停留在人体肠道微生物组上,而应扩展到植物的微生物群及其在植物生长发育中的功能。
参考文献:
Ruan Wenly,Engevik Melinda A,Spinler Jennifer K et al. Healthy Human Gastrointestinal Microbiome: Composition and Function After a Decade of Exploration.[J] .Dig Dis Sci, 2020, 65: 695-705.
Reza Musarrat Maisha,Finlay B Brett,Pettersson Sven,Gut microbes, ageing & organ function: a chameleon in modern biology?[J] .EMBO Mol Med, 2019, 11: e9872.
Lloyd-Price et al., The healthy human microbiome. Genome Medicine (2016) 8:51
Heribert Hirt, Healthy soils for healthy plants forhealthy humans.EMBO Reports (2020) 21: e51069
Haahtela T. A biodiversity hypothesis. Allergy. 2019 Aug;74(8):1445-1456. doi: 10.1111/all.13763. Epub 2019 Apr 4. PMID: 30835837.
Roslund MI, Puhakka R, Grönroos M, Nurminen N, Oikarinen S, Gazali AM, Cinek O, Kramná L, Siter N, Vari HK, Soininen L, Parajuli A, Rajaniemi J, Kinnunen T, Laitinen OH, Hyöty H, Sinkkonen A; ADELE research group. Biodiversity intervention enhances immune regulation and health-associated commensal microbiota among daycare children. Sci Adv. 2020 Oct 14;6(42):eaba2578. doi: 10.1126/sciadv.aba2578. PMID: 33055153; PMCID: PMC7556828.
相关阅读:

谷禾健康
S. pneumoniae (or pneumococcus,肺炎链球菌)是中耳炎、细菌性脑膜炎、败血症和社区获得性肺炎的主要病因。世卫组织将肺炎链球菌列为一种抗生素耐药的“优先病原体”。
肺炎链球菌是下呼吸道感染的主要原因之一,每年在全世界造成近100万儿童死亡。肺炎链球菌在人类宿主中占据不同的生态位,导致共生和致病性存在。侵袭性肺炎链球菌疾病是一个多步骤的过程。它是通过肺炎链球菌渗透进入富含糖的粘液层,然后粘附到人鼻咽的上皮细胞层而引发的。通常,微生物在鼻咽中定殖较长时间而不会引起疾病。
另外,由于未知的原因,它可以扩散到中耳,肺,大脑或血液中。直接从鼻咽或最经常通过肺部进入血液,可以进入中枢神经系统,心脏和脾脏。在所有这些组织中,肺炎链球菌都受到免疫系统和各种环境条件的攻击。

刚刚,来自美国宾夕法尼亚州匹兹堡卡内基梅隆大学生物科学系的一篇综合评论文章题为:“The pneumococcal social network” 综合阐述了人类致病菌肺炎链球菌分泌肽的机制和功能特点。讨论了三大类小肽的序列特征、转运机制和受体:双甘氨酸肽、Rap、RGG、NprR、PlcR和PRGX(RRNPP)结合肽,以及含羊毛硫氨酸的小肽。强调影响携带和致病的因素,特别是遗传多样性、微生物竞争、生物膜发育和环境适应。
注:羊毛硫氨酸是一类结构独特的肽类,因其序列中含有羊毛硫类非常规氨基酸而得名。
革兰氏阳性细菌使用一系列分泌肽来控制种群水平的行为,以响应环境提示。最近肺炎链球菌多肽研究的扩展揭示了一个相互作用的信号系统的复杂网络,其中多个多肽被整合到同一信号通路中,允许多个进入该通路的点,并向新的方向扩展信息内容。
此外,由于多肽存在于细胞外环境中,因此存在串扰、群体感应(QS)以及株内、株间和种间相互作用的机会。关于人群行为导致疾病的方式的知识为设计和开发抗感染策略提供了一条途径。
社会行为在生物体内广泛存在。蚁群的形成、蝗虫的协调运动和鱼类的浅水化都是复杂的社会行为例子。这些行为通过保护人们免受捕食、增加食物供应或比竞争对手具有战略优势而受益。细菌也不例外。
细菌执行群体感应(QS):细胞密度相关的信号传导,导致群体水平的反应。QS的早期证明是在海洋细菌fischeri弧菌中,高细胞密度诱导生物发光,这是细菌与鱿鱼共生关系的一部分。从那时起,细菌群的行为就被牵涉到细胞过程中,比如基因转移、运动、抗生素的产生和生物膜的形成。
细胞-细胞间的通讯是由供体细胞分泌到细胞外环境中的信号分子来协调的,并由产生细胞和邻近细胞感应到。感知导致基因表达的变化,最终触发同步的群体行为。这种细胞-细胞通讯的核心是通过自体诱导剂-2(AI-2)和肽发出信号。
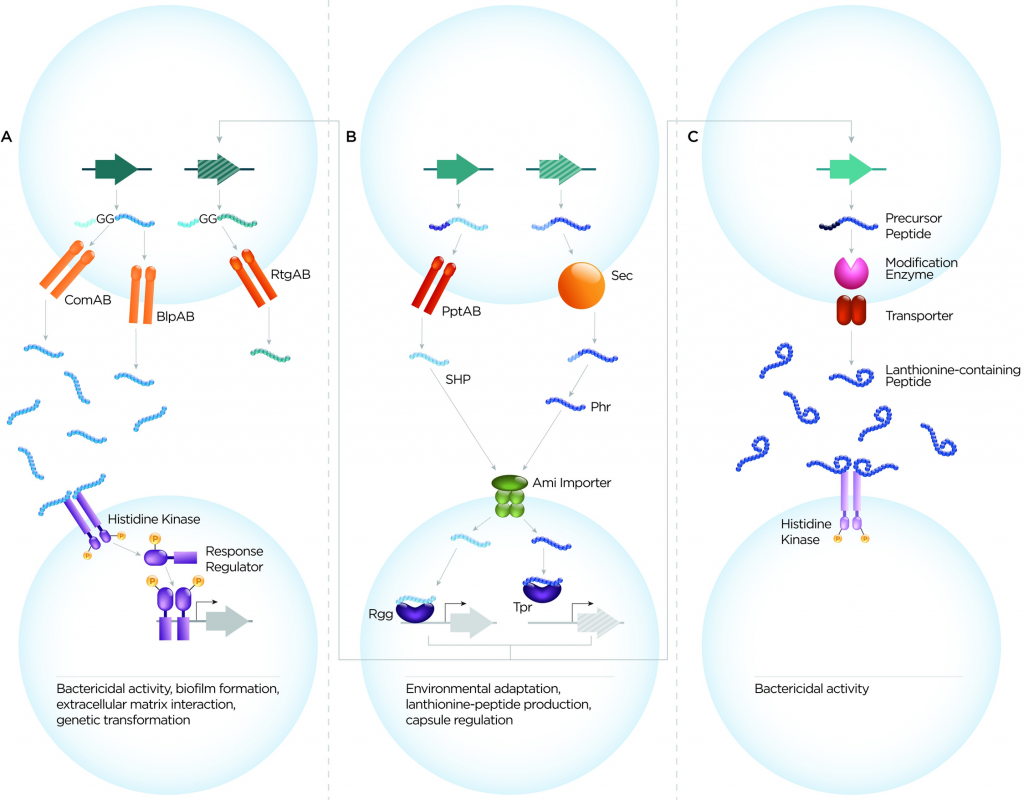
肺炎链球菌细胞-细胞通信系统可根据肽序列、转运蛋白和受体分为3大类(图1)。这些是(1)双甘氨酸肽,(2)与QS蛋白RRNPP超家族相关的肽,(3)含羊毛硫氨酸肽。从它们的作用来看,肽介导的细胞-细胞通信系统至少履行了三个主要功能:确保遗传多样性、微生物竞争和环境适应。
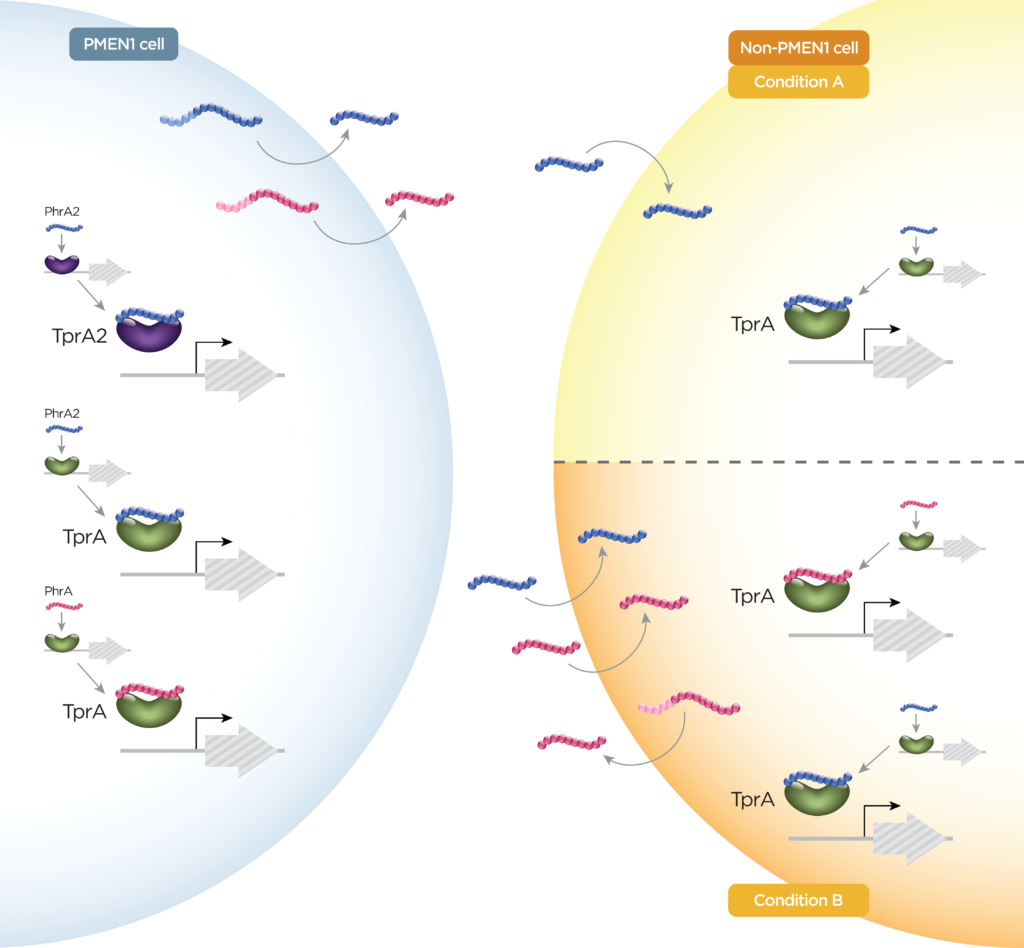
图1 肺炎链球菌分泌肽的三大类示意图

双甘氨酸肽
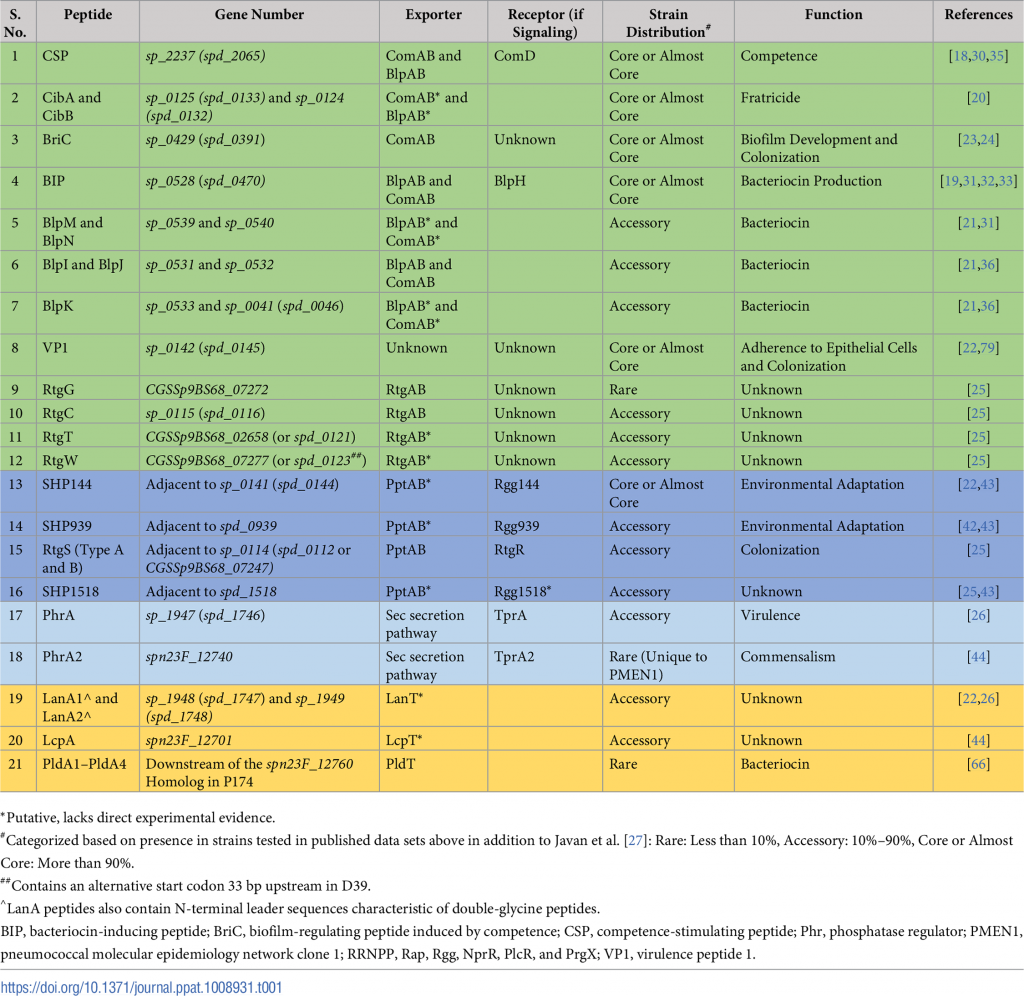
最具特征的肺炎链球菌双甘氨酸肽是能力刺激肽(CSP)。其他例子包括细菌素诱导肽(BIP)、感受态诱导细菌素(Ciba和CibB)、细菌素免疫区的肽(BIR)、毒力肽1(VP1)、感受态诱导的生物膜调节肽(BRIC)、RTG位点的肽和LANA(表1)。此外,比较基因组学方法已经揭示了额外的双甘氨酸肽,这些多肽还有待鉴定。
表1:核糖体合成肽在肺炎链球菌中的实验研究

双甘氨酸肽的特征是保守的N末端前导序列,该序列终止于Gly–Gly残基(或更常见于Gly–Ala或Gly–Ser)。前导将这些肽引导至含有肽酶的ATP结合盒(ABC)转运蛋白(具有C39肽酶结构域),其可切割前导序列并将肽输出到细胞外。 CSP和BIP的基因座分别编码同源的输出蛋白ComAB和BlpAB。移码突变使BlpAB在大约60%的菌株中失去功能。
QS蛋白的RRNPP超家族的肽
该组中的肽通过与它们的同源细胞质转录因子直接相互作用而发出信号,它们是RRNPP超家族的成员(图1B)。如前所述,这些肽可以基于多种序列特征进行分类。在肺炎链球菌中,已经鉴定了短疏水肽(SHP)SHP144,SHP939,SHP1518和RtgS,以及磷酸酶调节剂(Phr)肽PhrA和PhrA2。
在整个链球菌中,糖基转移酶(Rgg)蛋白质的调节基因与它们的同源SHP结合后被激活,后者通常在rgg基因附近编码。 SHP仅在出口后才有活性,这是由通常短于35个残基的前体多肽加工而成的。尚未在肺炎链球菌中研究加工蛋白酶,但在其他链球菌物种中,膜结合金属蛋白酶(Eep)有助于加工。
多种链球菌物种通过ABC转运蛋白PptAB出口SHP,其粪便肠球菌的同源物出口性信息素。在缺乏已知靶序列的情况下,将SHP引导至其转运蛋白的机制仍不清楚。成熟的肽通过寡肽渗透酶系统重新导入细胞,在那里它们与同源的Rgg调节剂相互作用,从而改变细胞的转录状态。
在肺炎链球菌中,RtgS是唯一显示其前体肽由PptAB出口并由Ami寡肽进口商AmiACDEF内化的SHP。鉴于PptAB在物种间的广泛功能,其他肺炎链球菌SHP可能利用相同的机制进行进出口。
含羊毛硫氨酸肽类
这是由革兰氏阳性细菌产生的一族小肽(19-38个氨基酸),具有各种结构和功能。这些是环状肽,其特征是翻译后修饰,导致硫醚氨基酸、羊毛硫氨酸和甲基羊毛硫氨酸的引入(图1C)。当前肽中的丝氨酸或苏氨酸残基脱水并与半胱氨酸硫醇连接时,它们的特征结构由LanM修饰酶形成。该肽通过专用的LanT转运蛋白输出。许多含羊毛硫氨酸的肽形成细菌素的两大类中的一种。
这些肽被称为羊毛硫氨酸抗生素或羊毛硫氨酸的抗生素。用于加工羊毛硫氨酸的肽,修饰酶,免疫蛋白和转运蛋白的基因通常以簇的形式组织。肺炎链球菌中有许多这样的簇。其中,已经研究了与Tpr / Phr和肺炎链球菌素簇相关的瘦肽。
肺炎链球菌肽介导的细胞-细胞通讯系统提供多种功能,而不是相互排斥。细胞-细胞通信系统可以被概念化为控制种群水平结构和行为的电路。
许多细胞-细胞通讯回路对不同的环境刺激作出反应,如菌群密度、营养状况、pH值、氧气供应和抗生素压力。来自细胞-细胞通讯系统的信号在群体水平的变化中表现为聚合。这些可能会影响生物膜的发育或与细胞表面成分的改变有关,如膜成分和胶囊表达。
细胞-细胞通讯系统的行为也可能伴随着DNA摄取、自残或杀菌能力的改变。这些生理变化可能改变细胞获得抗生素抗性基因的倾向,并影响疫苗逃逸株的出现。
此外,细胞间通讯系统还可以调节宿主基质的降解、生物膜的发育和营养吸收能力。因此,这些特性共同影响致病潜能、抗生素耐药性和对疫苗的反应。
调节群体反应的能力为肺炎链球菌细胞提供了相对于居住在鼻咽的其他微生物物种的竞争优势。细胞间的通讯系统使微生物能够改变其转录模式,以获得合适的表型,从而优化种群水平的适应性。它能在DNA水平和转录水平上引入多样性,从而能够长期维持鼻咽部的共生生活方式,在无症状时期,肺炎链球菌在高度可变的生物膜中被发现。
此外,一般来说,“转录适应”不仅有助于在动态的鼻咽中存活,而且在感染期间微生物从一个宿主生态位迁移到另一个宿主生态位时也能促进存活。
当检测到CSP,双甘氨酸肽家族的典型代表就会被激活。肺炎链球菌泛基因组拥有comC的6个不同等位基因,其中大多数菌株编码2个等位基因变体中的1个。在混合种群中,能力不仅在空间上局限于生物膜内的某个区域;信号可能局限于单个的信息类型(框1)。CSP是由高细胞密度、pH值增加、氧利用率和抗生素压力等因素引起的。
除了在环境中扩散外,CSP还通过其他机制(包括自分泌信号和细胞-细胞接触)向邻近细胞发出信号。激活会导致一个菌株的10%的基因发生转录变化。最典型的反应是激活转化机制,允许通过重组将外源DNA纳入,并且在这样做的过程中,通过等位基因的交换和基因拥有量的变化来促进遗传多样性的产生(图2)。
全基因组内遗传物质的交换测试了新颖的遗传组合,其中单个等位基因或基因片段已经克服了通过选择修剪的问题。
图2 多个双甘氨酸肽的分级活化
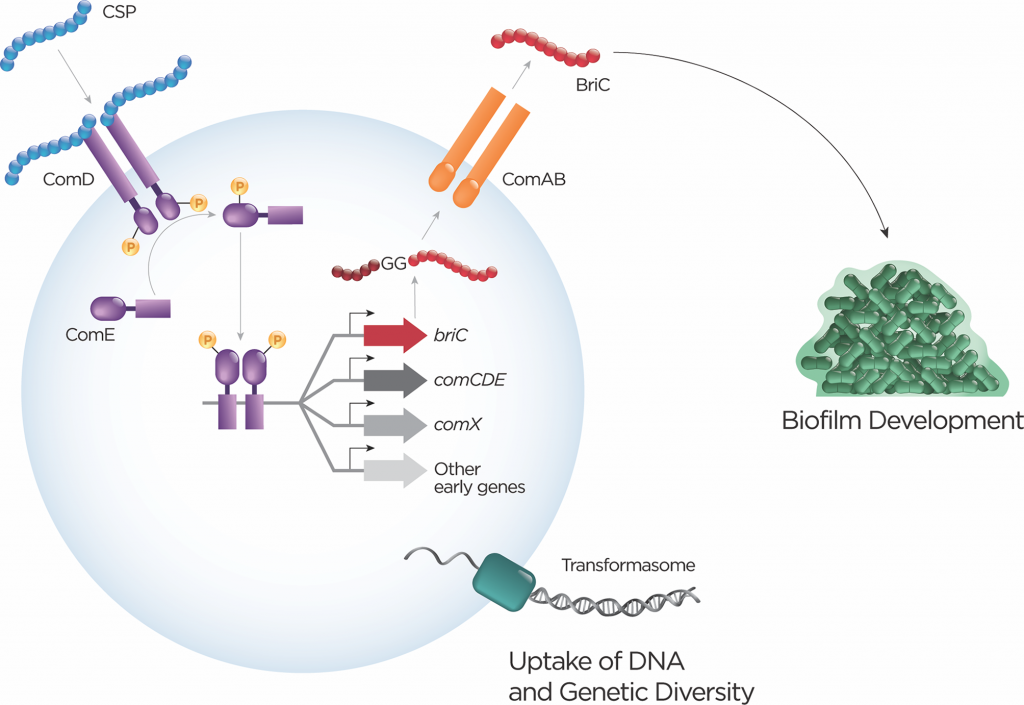

种内和种间竞争是呼吸道肺炎链球菌生活方式的一部分。杀菌活性通过双甘氨酸和羊毛硫氨酸的肽家族中的小分子产生。限制竞争者生长的能力对于细菌定殖很重要。此外,受害者可能成为DNA的来源,增加了进化的可能性。
能力激活导致许多这些效应分子的产生,包括CibAB和BIR基因座。双甘氨酸肽CibAB引发异源分解或反式裂解,并以细胞接触依赖性方式负责非功能细胞的裂解。与乳酸乳球菌IFPL105相似,据信这种杀微生物剂是通过将细菌素插入敏感细胞的膜中而导致其细胞能量消耗而产生的。
此后,细胞壁水解酶(包括自溶素LytA,溶菌酶LytC和莫林蛋白酶水解酶CbpD)的作用引起裂解。跨膜肽CibC保护细胞免受CibAB的同素分解作用。 CibAB从非感受态细胞释放的营养物质和DNA可能会使攻击细胞受益。在定殖的鼠模型中,CibAB诱导的同素分解作用使常驻菌株具有通过入侵菌株抵抗竞争和定殖的能力。
IR编码多种效应基因,赋予其抑菌作用或抑制作用的免疫力。 BIR基因座是同系的,但细菌素(假定的和特征的)和免疫蛋白在不同菌株之间差异很大。blpIJ,blpMN和blpK的产品已证实具有杀菌活性。这些是双甘氨酸肽,由BlpAB输出,并与同源免疫蛋白共转录。所有这些肽都显示出菌株间的活性,并且在体内,BlpMN和BlpIJ细菌素在定植过程中提供了比免疫缺陷型菌株更具竞争优势的菌株。
除肺炎链球菌外,细菌素基因座的表达还抑制了一些其他革兰氏阳性细菌,包括化脓性链球菌,链球菌性微生物,口腔链球菌和乳酸乳球菌,但不抑制其他细菌,例如变形链球菌,粪肠球菌或单核细胞增生李斯特菌。
BIR基因座的表达是由BIP诱导的,BIP由blpC和BIR的上游编码。与CSP相似,BIP结合膜结合的组氨酸激酶(B1pH),并且在肽及其受体之间具有特异性。这种特异性限制了竞争性表型之间的串扰。与能力途径的激活相似,BIP的表达是由抗生素诱导的,并且会增加pH值。而且,在能力和细菌素系统之间存在串扰,其中BIP的产生是在CSP刺激后诱导的。
另一类细菌素是羊毛硫氨酸的肽,其特征是肺炎链球菌素(pld)。pld基因座在肺炎链球菌分离株中很少见,其特征是有4个串联推定的短肽同源物(PldA1-PldA4)。这4种肽中的3种PldA1-3对细胞的杀菌活性是必需的,而第4种肽PldA4对于该表型是必不可少的。
除具有杀菌特性外,肺炎链球菌素(PldA1-3)还可作为自动诱导信号肽,通过位点(pldK)中编码的组氨酸激酶进行信号传导,从而激活pld位点。免疫是由邻近的ABC运输商PldFE授予的。这些肽的信号传导和杀菌作用是相互联系的:当Pld肽的信号传导低时,PldA2不会诱导细菌抑制作用。另外,肺炎链球菌素在小鼠定植过程中为肺炎链球菌菌株提供了竞争优势。
最后,全面的比较基因组筛选揭示了许多其他肽,它们在肺炎链球菌菌株之间分布不同(从稀有到核心),而且是肺炎链球菌特有的或在链球菌种之间共有。许多细胞与推定的转运蛋白,修饰蛋白或免疫蛋白一起组织成操纵子。肺炎链球菌菌株和相关物种内的多样性分布与种内和种间微生物竞争中的作用一致。
肺炎链球菌有一个细胞间通信系统网络,可调节其对宿主环境的适应性。肺炎链球菌只能使用糖来产生其代谢能。此外,糖被用于胶囊生产和信号传递。此外,通过磷酸转移酶系统(PTS)转运糖可以触发磷酸化依赖性信号通路。此外,宿主糖的降解不仅是营养的来源,而且还是宿主粘附,定植和致病性的主要因素。
几种细胞间通讯肽的表达对宿主碳水化合物的水平有反应:甘露糖和半乳糖诱导SHP144和SHP939,半乳糖诱导PhrA。另外,这些肽以及PhrA2和VP1在丰富的培养基中被阻遏。Rgg144 / SHP144系统是核心。当自诱导肽SHP144导入细胞并结合Rgg144时,它被激活。Rgg144受主要营养调节剂CodY和谷氨酰胺/谷氨酸代谢的负面控制。
SHP939是一种正向调节Rgg939的自诱导肽。该系统是辅助基因组的一部分。 Rgg939 / SHP939调控的基因多样性随环境条件的变化而变化,当在甘露糖上生长时具有广泛的调节子,而在半乳糖中则具有有限的调节子。
Rgg系统并非孤立运行,而是看起来形成了一个连接的网络。因此,为了最大程度地诱导SHP144和SHP939,需要存在非同源Rgg调节剂。此外,Rgg1518控制其邻近基因座(SPD_1513–1517),这些基因也受Rgg144和Rgg939调控。这种串扰可能会超出物种的范围。 SHP939的序列与化脓性链球菌中发现的SHP3的序列相同,无乳链球菌和米氏链球菌中的SHP序列相差1个残基。SHP序列的相似性可能使肺炎链球菌Rgg系统受到其他常驻细菌的影响。
类似于TprA / PhrA,PhrA2与TprA2相互作用,导致TprA2调节子的阻遏并诱导紧靠下游的羊毛硫氨酸肽(LcpA)的表达。有证据表明TprA2 / PhrA2与TprA / PhrA系统之间存在单向串扰(图3)。
图3 如PhrA和PhrA2所示的肽串扰示例
肺炎链球菌所编码肽的多样性凸显了群落水平表型在引入遗传多样性、微生物竞争和环境适应方面的重要性。一些研究证实了细胞间通讯在肺炎链球菌毒力中的作用。需要更多的研究来绘制这些系统在感染过程中的时空表达。
上呼吸道是许多细菌物种的常住栖息地。为了在宿主环境中立足,肺炎链球菌细胞必须在竞争中生存并抵抗住其他寄居物种的挑战。肺炎链球菌产生大量的肽,直接或间接地通过激活下游分子(例如,由CSP激活CibAB或CbpD)介导杀菌活性。虽然其中一些细菌素已被鉴定,但许多假定细菌素有待功能鉴定,尤其是在目标菌株/物种和刺激其合成的环境条件下。
多种肺炎链球菌肽与生物膜形成有关,包括CSP、BriC和VP1。生物膜在运输和疾病中起着关键作用。载体包括一种生物膜生长模式,它本身能够实现局部细胞-细胞通讯(通过肽和AI-2)和吸收生物膜基质中提供的DNA。
此外,生物膜为表型异质性提供了一个平台,这是一个未被充分研究的课题,可能有助于肺炎链球菌适应慢性感染和适应宿主生态位。生物膜促进发病。它们不仅是细菌传播的场所,而且从生物膜中传播的细菌比它们的生物膜或浮游生物具有更强的毒性。
综上所述,肺炎链球菌信号肽和生物膜紧密相连:生物膜中的局部环境可能促进细胞-细胞通讯的条件,而肽会影响生物膜的发育和向其他组织和新宿主的扩散。
肽对调控网络的控制使肺炎链球菌不仅在单个细胞水平上,而且在整个群体水平上对其环境作出快速反应。系统间的串扰和由多个肽对一条通路的调节使不同的信号得以整合,可能会增加反应的动态性和复杂性。研究表明,多链肺炎链球菌共定植是一种相对常见的现象,突出了跨菌株肽交换的重要性。PhrA2和PhrA存在串扰(图3)。
来自同一家族或不同家族的肽可以在同一途径中发出信号。例如,通过CSP的信号转导可诱导多个双甘氨酸蛋白,而SHP144诱导VP1(一种双甘氨酸肽)的水平。这种肽激活的层次结构描述了生物途径激活的多个入口点的存在。这表明这些途径的激活是严格控制的,代表了相关但不同的可能性。
一种可能性是,不同的信号组合允许诱导整个通路或通路的一部分,提供与反应相关的类型、幅度和代谢成本的粒度。或者,当遇到合适的条件时,细胞可以激活肽信号,为诱导下游通路做好准备。
抗生素耐药性是一个全球性的公共卫生问题,因此迫切需要开发有效的抗感染药物。非抗生素抗感染药物有望减少抗生素耐药性的出现。开发破坏细胞-细胞通讯的化合物的不同策略包括抑制信号产生、信号降解或阻断信号转导。在化脓链球菌和其他一些链球菌中,化合物的使用会破坏Rgg/SHP信号传导,从而破坏生物膜的形成。在肺炎链球菌中,使用竞争性类似物可以抑制能力发展和水平基因转移。此外,以可溶性LMIP形式靶向PhrA肽的抗感染药物可降低肺炎链球菌介导的小鼠发病率。不同的抗感染药物是如何影响细菌适应度和促进耐药性发展的,还有待检验。肺炎链球菌细胞间通讯系统的多样性为探索不同的可能性提供了许多机会。对肺炎链球菌群体水平表型的研究为开发新的抗感染靶向肽介导系统以及这些系统调控的途径提供了机会。
参考文献:
Aggarwal SD, Yesilkaya H, Dawid S, Hiller NL (2020) Thepneumococcal social network. PLoS Pathog 16(10): e1008931.
Bassler BL. Small talk: Cell-to-cell communication in bacteria.Cell. 2002;109:421–424. pmid:12086599
Slager J, Kjos M, Attaiech L, Veening JW. Antibiotic-inducedreplication stress triggers bacterial competence by increasing gene dosage nearthe origin. Cell. 2014;157:395–406. pmid:24725406
Miller EL, Evans BA, Cornejo OE, Roberts IS, Rozen DE. Pherotypepolymorphism in Streptococcus pneumoniae has no obvious effects on populationstructure and recombination. Genome Biol Evol. 2017;9:2546–2559. pmid:28992304
Junges R, Sturød K, Salvadori G, Åmdal HA, Chen T, Petersen FC.Characterization of a signaling system in Streptococcus mitis that mediatesinterspecies communication with Streptococcus pneumoniae. Appl EnvironMicrobiol Microbiol. 2019;85:e02297–e02218. pmid:30389765
谷禾健康
细菌感染严重威胁人类健康。如果你去医院就医,医生怀疑你有感染,可能会要求进行革兰氏染色检查。该测试还可以帮助医生了解细菌是革兰氏阴性还是革兰氏阳性,从而选择合适的治疗方式。
然而你可能对于革兰氏阳性菌阴性菌这些概念并不熟悉,今天为大家讲解下,革兰氏阳性菌,革兰氏阴性菌的区别,革兰氏染色法,常见的革兰氏阳性菌类型以及相应的治疗。
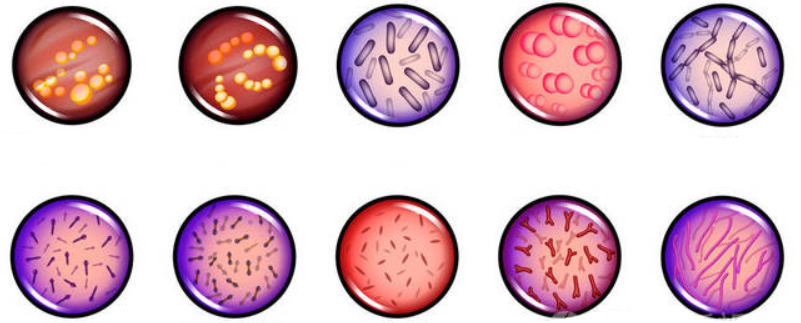
首先,我们了解下,革兰氏阳性菌和阴性菌各自的特征。
革兰氏阳性菌特征
革兰氏阳性细菌的主要特征是它们的结构。一般有以下特点:
没有外膜。革兰氏阳性细菌没有外膜,但革兰氏阴性细菌有。
复杂的细胞壁。包围细胞质膜的细胞壁由肽聚糖、多糖、磷壁和蛋白质组成。它很容易吸收外来物质。
厚的肽聚糖层。在革兰氏阳性细菌中,肽聚糖有40到80层厚。
某些表面附属物。革兰氏阳性细菌可能有鞭毛,可以帮助它们移动。它们很少有被称为菌毛的毛发状结构。
革兰氏阴性菌特征
革兰氏阳性菌和革兰氏阴性菌具有不同的结构。通常,革兰氏阴性菌具有以下特征:
外脂质膜
肽聚糖薄层(2 – 3纳米)
通常不含磷壁酸
可以有鞭毛或毛
主要的区别是外脂膜。它很难渗透,这给了革兰氏阴性细菌额外的保护。革兰氏阳性细菌没有这种特征。
由于这种差异,革兰氏阴性细菌更难杀死。这意味着革兰氏阳性菌和革兰氏阴性菌需要不同的处理方法。
革兰氏阳性菌和阴性菌是怎么鉴定的呢?
——通过革兰氏染色法。
革兰氏染色法是一种根据细菌细胞壁进行细菌分类的方法。它使科学家能够确定一个有机体是革兰氏阳性还是革兰氏阴性。这项使用显微镜的测试是由汉斯·克里斯蒂安·克拉姆在1884年发明的。
在这个过程中,结晶紫染料被应用到细菌样本上。这种化学染料能将较厚的肽聚糖层染色。
在显微镜下,革兰氏阳性细菌呈现紫蓝色,因为它们厚厚的肽聚糖膜可以容纳染料。由于阳性结果,这种细菌被称为革兰氏阳性细菌。
革兰氏阴性菌染成粉红色。它们的肽聚糖层更薄,所以不会保留蓝色。测试结果为阴性。
革兰氏染色可在各种类型的标本上进行,包括:
血液、组织、粪便、尿液、痰液等。
医生将样本送到实验室进行革兰氏染色检测,从而了解是否是细菌导致了该症状,以及存在哪些类型的细菌。他们也可能会要求其他测试来帮助诊断疾病。这对于确定后续的治疗方案很有帮助。
根据不同的特性,革兰氏阳性菌可进一步分为以下几类:
革兰氏阳性球菌
革兰氏阳性球菌呈圆形或椭圆形。术语“球菌”的意思是球形,表明细菌通常是圆形的。
以下是革兰氏阳性球菌:
葡萄球菌属
葡萄球菌呈葡萄状丛生生长。正常情况下,它们存在于我们的皮肤和粘膜上,不会造成任何问题。但如果葡萄球菌进入人体,它们会引起严重的感染。
链球菌属
链球菌呈链状生长。这是因为细胞在分裂后并没有完全分离。
与葡萄球菌一样,链球菌通常存在于体内。它们通常存在于皮肤、口腔、肠道和生殖道。
链球菌可分为以下类别:
S. pyogenes (Group A)、S. agalactiae (Group B)、Enterococci (Group D)、S. viridans、S. pneumoniae
革兰氏阳性杆菌
当革兰氏阳性细菌呈棒状时,它们被称为杆菌。这些细菌大多在皮肤上发现,但有些会导致严重的疾病。
革兰氏阳性杆菌根据其产生孢子的能力被进一步分类。
有芽胞型:
芽孢杆菌和梭状芽孢杆菌可以形成孢子,这有助于细菌在高温等恶劣条件下生存。
这些杆菌根据它们对氧气的需要被细分。芽孢杆菌需要氧气才能生存(需氧),而梭状芽孢杆菌不需要氧气(厌氧)。
无芽胞型:
李斯特菌(Listeria )和棒状杆菌(Corynebacterium)不产生孢子。李斯特菌是厌氧菌,棒状杆菌是好氧菌。
许多革兰氏阳性菌都是致病菌。虽然有100多种致病性革兰氏阳性菌,但最显著的种类包括:
葡萄球菌
葡萄球菌通常是细菌感染的原因。大多数情况是由以下物种引起的。其他致病性葡萄球菌不太常见,很少导致疾病。
金黄色葡萄球菌
金黄色葡萄球菌是致病性最强的葡萄球菌。它导致了大多数葡萄球菌感染,包括:
皮肤感染,如蜂窝织炎和毛囊炎、感染性关节炎、脓肿、心内膜炎、细菌性肺炎、食物中毒、中毒性休克综合征、烫伤皮肤综合征、MRSA等。
表皮葡萄球菌
通常,表皮链球菌会在免疫系统薄弱或住院的人身上引起感染。会引起:
导尿管等医疗器械感染、菌血症、纵隔炎、手术部位感染、眼部角膜炎、眼内炎(内眼感染)等。
腐生葡萄球菌
通常见于生殖道和会阴。会引起: 无并发症的尿路感染(最常见)、尿道炎、前列腺炎、急性肾盂肾炎 、附睾炎等。
链球菌
链球菌也是常见的致病菌。以下是最常见的生物体。一般来说,其他链球菌群可引起食源性疾病,引起喉咙痛。
肺炎链球菌
肺炎链球菌是社区获得性肺炎最常见的病因。它还引起:红眼、鼻窦感染、脑膜炎等。
酿脓链球菌
会引起脓毒性咽喉炎、蜂窝织炎、咽炎、脓疱炎、猩红热、风湿热、坏死性筋膜炎、肾小球肾炎等。
无乳链球菌
通常会引起新生儿感染,包括:败血症、肺炎、脑膜炎、化脓性关节炎。
肠球菌
肠球菌主要在结肠中发现。它们会引起胆道和泌尿道感染。
芽孢杆菌
作为孢子形成细菌,杆菌产生孢子并释放毒素。大多数杆菌对人类没有致病性,但以下两种可导致严重的疾病。
炭疽杆菌(Bacillus anthracis )
炭疽杆菌孢子产生炭疽毒素,引起严重的疾病。人类可通过吸入或接触受感染的动物而感染炭疽。
根据炭疽热的传播方式,它可以引起各种症状。这些可能包括:恶心、呕吐、腹痛、咳血、高烧等。
蜡样芽胞杆菌(Bacillus cereus )
蜡样芽孢杆菌是一种孢子形成细菌,存在于土壤和一些食物中。它与吃未煮熟或重新加热的米饭而导致的疾病有关。会引起:腹泻、恶心、伤口感染、呼吸道感染、眼内炎等。
梭菌(Clostridium)
大约有30种梭状芽孢杆菌会引起人类疾病。像杆菌一样,这些细菌形成毒素,导致严重的情况。梭状芽孢杆菌通常与食源性疾病有关,但最令人担忧的细菌包括:
肉毒梭状芽胞杆菌(Clostridium botulinum )
肉毒杆菌的孢子产生肉毒杆菌毒素,这是对人类最危险的毒素。它会导致肉毒中毒,包括:
食源性肉毒中毒(最常见)、婴儿肉毒中毒、伤口肉毒中毒、吸入肉毒中毒等。
产气荚膜梭菌(Clostridium perfringens)
产气荚膜通常与肉类生产和加工有关。如果一个人吃了受污染的肉,他们可能会食物中毒。症状包括持续不到24小时的腹泻和腹部绞痛。
艰难梭菌(Clostridium difficile )
艰难梭菌通常影响住院的老年人。一般发生在服用抗生素之后,引起结肠炎、腹部绞痛、严重腹泻。
破伤风梭菌(Clostridium tetani)
破伤风梭菌孢子产生破伤风毒素,一种神经毒性物质。一般它会出现在土壤、灰尘和生锈的工具上,如果这种毒素引起感染,就叫做破伤风。这是个严重的医疗紧急事件。
单核细胞增多性李斯特氏菌(Listeria monocytogenes )
唯一致病性李斯特菌是单核增生杆菌。在健康人身上,它通常会引起轻微的食源性疾病症状。但在免疫系统较弱的人群中,这种细菌会导致危及生命的情况:脑膜炎、败血症、利斯特氏菌病。
白喉杆菌(Corynebacterium diphtheriae)
约有30种棒状杆菌与人类疾病有关。然而,这些微生物很少引起疾病,通常影响免疫系统受损的人。白喉是本组的主要致病菌。引起:白喉、咽炎、呼吸道感染、感染性关节炎、皮肤感染、骨髓炎、心内膜炎等。
治疗革兰氏阳性细菌引起的疾病时,最佳选择取决于:细菌类型、抗生素耐药性、细菌是否形成毒素等。常见的治疗方法包括:
盘尼西林
一种常见的用于多种感染的抗生素。主要是干扰细菌的肽聚糖层,从而杀死细菌。主要用于链球菌感染,包括:链球菌性喉炎、鼻窦感染、尿路感染、蜂窝织炎等。
糖肽类
糖肽类抗生素常用于治疗耐药细菌引起的严重感染。像青霉素一样,它们通过破坏细菌的细胞壁来发挥作用。可以治疗:耐多药肺炎、MRSA、结肠炎。
红霉素
红霉素属于大环内酯类抗生素,大环内酯类抗生素还包括更知名的阿奇霉素和克拉霉素。它是一种能阻止细菌生长的抗生素,对革兰氏阳性和革兰氏阴性细菌都有效。
通常,红霉素是给对青霉素过敏的人开的。治疗的情况如下:细菌性肺炎、红眼、咽喉炎、葡萄球菌性皮肤感染等。
液体疗法
在某些情况下,治疗可能包括液体疗法。它有助于补充身体的水分水平和防止脱水。一般来说,治疗毒素引起的疾病需要液体管理。
抗毒素
对于与毒素相关的疾病,如炭疽和肉毒中毒,治疗包括抗毒素。这种药通过靶向和清除体内毒素而起作用。合适的抗毒素取决于具体的毒素。此外,它还与其他疗法结合使用。
革兰氏染色测试可以帮助医生诊断疾病,从而选择最佳的治疗方式,如果能更快速准确地了解是由何种菌引起的疾病,将会为治疗提供捷径。
【参考文献】
Gillespie S, et al. (2012). Medical microbiology and infection at a glance. West Sussex, UK: Wiley-Blackwell.
Kulkarni AP, et al. (2019). Current perspectives on treatment of gram-positive infections in India: What is the way forward?
Mahony J, et al. (2015). Gram-positive phages: From isolation to application. Lausanne, Switzerland: Frontiers Media SA
Malanovic N, et al. (2016). Gram-positive bacterial cell envelopes: The impact on the activity of antimicrobial peptides.
Sizar O, et al. (2019). Gram positive bacteria.
Vazquez-Guillamet C, et al. (2014). Treatment of gram-positive infections in critically ill patients.