 国家高新企业 | ISO9001认证
国家高新企业 | ISO9001认证 二级病原微生物安全实验室
二级病原微生物安全实验室- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
国家高新企业 | ISO9001认证 二级病原微生物安全实验室

谷禾健康
抗生素已经彻底改变了现代医学,但其有效性受到多重耐药细菌传播的威胁,而目前尚无有效的治疗方法。抗生素耐药性(AMR)通常由不必要的抗生素使用引起,是一种严重的全球健康和经济威胁。
特别值得关注的是,抗生素生产线不断减少,开发中的抗生素数量有限(数量和多样性都有限),以满足当前和预期的患者需求。此外,科学和经济挑战促使许多大型全球制药公司停止其抗生素开发计划,使得对新的感染控制方法的需求更加迫切。因此迫切需要用于治疗革兰氏阴性感染的新疗法。
最近一种潜在的方法是使用活的掠食性细菌。由于蛭弧菌Bdellovibrio bacteriovorus广泛存在于自然界中,而目前也尚没有与蛭弧菌相关的疾病报告,科学家认为有可能围绕它们建立一种安全无害的生物治疗手段,去抗击病原菌的感染。但毕竟蛭弧菌本身也是一种细菌,人和动物的免疫系统如果发现它们,会怎样对待这些“友军”?会增强对蛭弧菌的抵抗力吗?
本文讨论了支持掠食性细菌替代抗生素的可行性的基础科学。
蛭弧菌 Bdellovibrio bacteriovorus是寄生于其他细菌(也可无寄主而生存)并能导致其裂解的一类细菌。它虽然比通常的细菌小,能通过细菌滤器,有类似噬菌体的作用,但它不是病毒,确确实实是一类能”吃掉”细菌的细菌。
1962年首次发现于菜豆叶烧病假单胞菌体中,随后从土壤、污水中都分离到了这种细菌。根据其基本特性,命名为Bdellovibrio bacteriovorus。
其中,” Bdello”一词来自希腊字,是”水蛭”的意思,”vibrio”意为”弧菌”,而种名”bacteriovorus”是 “食细菌”的意思。“捕食”的对象正落在多出产致病菌的细菌类群——革兰氏阴性菌(如大肠杆菌、志贺氏菌等)中。

B. bacteriovorus通常会持续3-4 h消耗其革兰氏阴性细菌的猎物。这种掠夺性生命周期是一个复杂的过程,在分子水平上已经开始被理解。最初,B. bacteriovorus识别,附着并进入猎物细胞,对其进入口进行加固,穿越和重新密封。侵袭伴随着猎物细胞的圆形化,稳定的胶质细胞的形成以及猎物细胞的死亡。
同时,通过依次释放一组酶,B. bacteriovorus消化了猎物,并利用所形成的养分库长出了长丝。 单个细丝的同步分裂会产生奇数或偶数个子代细胞,每个子代细胞会产生鞭毛或滑行引擎(取决于条件),然后从死亡的猎物细胞中爆发并开始寻找新的猎物。 根据猎物细胞的大小,以及猎物细菌内部可用的养分,每个细胞平均释放4–6个B. bacteriovorus后代。 尽管被认为是专性的捕食性细菌,但B. bacteriovorus仍可转变为不依赖宿主的生活方式,可以在全培养基上的无菌生长。
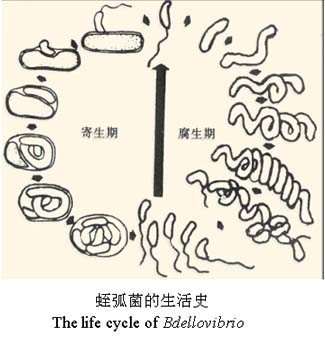
B. bacteriovorus生命周期和基因组的一些特征决定其成为一种潜在的抗革兰氏阴性细菌病原体的治疗剂。在捕食生命周期中,被捕食细胞在短时间内(<30分钟)被杀死,因此,被捕食者必须快速表达防御手段,以抵抗捕食,这是尚未见到的。
与某些抗生素不同,某些抗生素可引起一连串事件,导致细菌自溶和炎症分子释放,B. bacteriovorus细菌捕食不会导致猎物的最初溶解,因为在溶解之前,猎物的内容物是从稳定的蛭质体结构内消耗的。此外,没有单一的受体识别和附着猎物。在猎物入侵后,在数量和功能多样性方面,具有潜在遗传冗余的猎物破坏酶都出现了上调,这表明简单的猎物对B. bacteriovorus捕食的抵抗不太可能发生。
为了实现B. bacteriovorus的治疗潜力,必须在生物学相关系统中充分表征其对革兰氏阴性病原体的捕食性。 这种表征还必须同时解决诸如宿主反应,毒性,炎症,组织损伤或伤口愈合抑制等问题。
其中一些问题已通过体外细胞培养和体内动物模型解决,评估了掠食性细菌对免疫系统各个组成部分以及整个宿主的影响。 许多人类细胞系,包括角膜-上皮细胞,血液单核细胞,巨噬细胞,肾上皮细胞,肝上皮细胞和脾单核细胞,已经暴露于不同的捕食性细菌中,测定了不同暴露的时间范围、持续时间(2至24小时),下促炎和抗炎细胞因子水平。
已知可响应细菌外膜脂多糖(LPS)而被刺激的细胞因子,是宿主抵抗病原体所必需的。 这些研究表明,尽管B. bacteriovorus在免疫反应方面并不沉默,但暴露后产生的炎性细胞因子水平可忽略不计或很低。 B. bacteriovorus不像同时检测到革兰氏阴性病原体那样具有免疫刺激性,这可能部分是由于其独特的脂质A结构以及它拥有带鞘的鞭毛。
额外的细胞活力成像,细胞毒性测量以及暴露于掠食性细菌后对动物和人类培养细胞形态变化的评估初步表明,B. bacteriovorus对人细胞无毒,尽管还需要进行更多的研究。 Raghunathan及其同事使用人类巨噬细胞系(U937细胞)研大量掠食性细菌能够在细胞内存活长达24小时,从而确定了一段持久性和潜力捕食细胞内病原体的能力。
另外,通过在药理学抑制剂存在下进行吸收实验,证明了宿主肌动蛋白细胞骨架的作用及其在B. bacteriovorus吸收中的重排。B. bacteriovorus最终通过吞噬体途径被转运,这是由于它们靶向酸性液泡。 鼠巨噬细胞系中的类似观察结果支持了这一点,并且两者都是考虑给予B. bacteriovorus作为治疗剂的重要观察结果。 这些研究说明了B. bacteriovorus靶向细胞内病原体的潜力,而许多抗生素和其他生物控制剂(例如噬菌体)可能无法利用这些细菌。 尽管后一点正在积极研究中。 这些体外细胞实验以及下面描述的动物模型,是研究功效,缺乏毒性和潜在捕食者生物利用度的重要步骤。
早期体内宿主反应集中于B. bacteriovorus在吸热和放热脊椎动物肠道中的生存能力。研究了在鲶鱼(点状黄疸)、豹蛙(林蛙)、小鼠和兔子的肠道中实验投喂的B. bacteriovorus菌株MS7的生存能力和持久性,表明在接种后24-48小时内,B. bacteriovorus几乎没有恢复。通过饮用水给小鼠提供的3天B. bacteriovorus不能从肠道中完全恢复。同样,在注射后24小时左右,无论是使用无菌培养物,还是同时注射B. bacteriovorus,都没有或很少从兔回肠环中发现B. bacteriovorus。
B. bacteriovorus对动物没有致病性。这些研究已经扩展到包括评估宿主发病率、组织病理学、促炎和抗炎细胞因子水平、体内捕食性细菌传播以及长期评估大鼠、小鼠斑马鱼幼虫模型的一般健康状况。总的来说,在一系列动物模型中以及通过多种给药途径在体内给药已经证明,它们不会损害这些动物的健康。
事实证明,B. bacteriovorus对多种猎物具有体外功效,包括与肠道,口腔,伤口和眼部感染有关的细菌和生物制药。B. bacteriovorus成功地减少了实验室缓冲液和人血清中的病原体数量和生物膜中的猎物,而生物膜通常是抗生素治疗的重大阻碍。 重要的是,已经显示出许多具有多重耐药性的人类临床分离株容易被B. bacteriovorus捕食。 该清单包括许多需要新治疗的E(S)KAPE病原体,包括表达mcr-1的革兰氏阴性大肠菌素抗性分离株。 体外研究还调查了成功捕食所需的B. bacteriovorus与猎物的比率; 如果抗生素不能治疗感染,则作为将来的体内实验和临床的重要考虑因素。

尽管实验性体外系统,尤其是实验室缓冲液或富含生长培养基的环境可能显示出掠食性功效,但它们不足以替代体内治疗的复杂性,而体内治疗必须考虑到宿主的免疫反应以及病原体的生存策略。 动物感染模型在评估B. bacteriovorus在体内捕食革兰氏阴性细菌的能力以及解决此类宿主对宿主的任何反应(以及由此产生的安全性)方面都发挥了作用,越来越多的证据表明,至少在动物中,以及将来将这种掠食性细菌用于治疗应用的可行性,将其扩展到人类。
一项对感染肠炎沙门氏菌p125109的鸡群实验表明。 在B. bacteriovorus处理后的三天内,与对照动物相比,经细菌性芽孢杆菌处理的禽类的盲肠中沙门氏菌数量显着降低。 此外,与对照动物的许多盲肠相比,经B. bacteriovorus处理的禽中盲肠的外观是正常的。
进一步的重要研究表明B. bacteriovorus能成功地在体内捕食病原体。 使用大鼠模型,Shatzkes及其同事证实B. bacteriovorus可以治疗肺炎克雷伯菌感染的大鼠,显示与对照组相比,细菌性芽孢杆菌治疗的动物的病原体负担明显减少。 另外一项通过向大鼠尾静脉注射引发疾病的败血病模型。 B. bacteriovorus不能显着减少感染。 这是首次将B. bacteriovorus直接施用到血流中,并且是评估掠食性细菌清除血流感染能力的重要一步。
斑马鱼幼虫的物理特征以及与人类的广泛基因组同源性包括良好的理解,充分发展的免疫系统和光学透明的性质,非常适合于创新的活荧光显微镜检查。
诺丁汉大学伊丽莎白·肖克特和伦敦帝国理工学院的塞尔吉·莫斯托维小组用模式生物斑马鱼做了研究。

接下来,研究者放心地开始了“以菌治菌”环节。他们用一种叫福氏志贺氏菌(Shigella flexneri)的病原菌来感染斑马鱼——所选取的菌株同时对链霉素与羧苄青霉素具有耐药性。他们向斑马鱼的后脑先接种了致死剂量的志贺氏菌,再注射蛭弧菌。由于这两种细菌被带上了不同颜色的荧光蛋白标记,研究者得以观察它们的数量和分布变化情况。他们发现,相比于对照组,注射了蛭弧菌的斑马鱼后脑内志贺氏菌大量减少,被感染的斑马鱼在72小时后的存活率也更高。在更高分辨率的显微观察中,研究人员也找到了蛭弧菌在斑马鱼体内和体外都能够侵染并杀死志贺氏菌的证据。
研究者认为,在抗击志贺氏菌感染这件事上,蛭弧菌和斑马鱼免疫系统其实某程度上达成了巧妙的“配合”:蛭弧菌对志贺氏菌的“捕食”开始得非常迅速,能够在感染初期控制住志贺氏菌繁殖的势头,帮免疫系统减轻应对的压力。而等到大批白细胞赶来时,蛭弧菌已经饱餐过一顿,收拾残余志贺氏菌的工作,免疫系统自己也能完成好。
1897年,日本细菌学家志贺洁发现了志贺氏菌(Shigella).
作为杆菌性痢疾的祸首,这类细菌每年导致约1.63亿严重痢疾病例,并夺走超过100万人的生命,可谓是最臭名昭著的病原菌之一。志贺氏菌是一类革兰氏阴性的杆状细菌,可以制造出能杀死细胞的志贺毒素。
在最近的一项研究中,Russo及其同事证明了B. bacteriovorus 可用于显着减少实验感染小鼠肺部的鼠疫杆菌(yersinia pestis)数量。 但是并非所有的体内B. bacteriovorus 施用都已成功地减少了病原体数量。 尽管在先前的研究中使用组织培养模型在体外取得了令人鼓舞的结果,但给予B. bacteriovorus 治疗经牛莫拉氏菌(Moraxella bovis )感染的牛犊是牛角膜结膜炎的病原体,未能导致角膜溃疡形成的显着改善。
从上面的大量研究可以看出,越来越多的证据表明Bdellovibrio sp持续非病原性且足够长以具有治疗活性,对免疫微生物群的不良影响极小,并且不成为正常宿主微生物群的一部分。 无论是成功的还是不成功的体内试验,如本文所述,对于评估哪些适应症,给药途径(局部给药还是静脉给药)和感染部位都至关重要,潜在的掠食性细菌给药将最有效地对其进行评估。
治疗后动物和人类感染的康复结果取决于多种因素的相互作用,例如患者的免疫系统和总体健康状况包括肠道菌群,病原体的性质以及治疗的类型和提供方式。 在将体外和体内细菌捕食视为宿主反应的独立实体时,未捕获革兰氏阴性病原体的细菌捕食与宿主反应之间的相互作用和协同作用。

该模型表明,B. bacteriovorus在非致病性条件下持续了足够长的时间,可以在实验性感染期间有效地捕食志贺氏菌。 在这项工作中,用显微镜观察了与宿主免疫系统细胞的相互作用。 此外,这项研究发现B. bacteriovorus的最大治疗益处来自细菌捕食和宿主免疫系统的协同作用。 给药后,掠食性细菌种群最终会通过与宿主免疫系统的相互作用而清除,实际上这种清除可能对(自我)限制治疗有益。
在考虑治疗掠食性细菌感染时,研究人员可以向噬菌体研究和临床界寻求动力,以应对从体外到临床环境的挑战。 尽管在一些国家已经使用了多年,但最近的一些引人注目的病例已证明在临床环境中成功施用了噬菌体鸡尾酒来治疗耐药性感染。
从表面上看,B. bacteriovorus和噬菌体具有许多潜在的生物防治剂特性。 它们都表现出掠食性或寄生性生命周期,它们是自我复制和自我限制的,仅在存在易感宿主或猎物时才持续存在,并且在裂解性噬菌体的情况下,两者都具有广泛相似的生命周期。 当将它们引入动物和人类体内时,它们似乎也几乎没有副作用。 但是,两者之间存在一些显着差异,这可能会限制和区别它们在某些适应症中的使用。
首先,与噬菌体相比,B. bacteriovorus无疑是“活的”并且具有代谢活性,噬菌体是惰性颗粒,直到它们通过特定受体与宿主建立接触。 这可能对B. bacteriovorus既有好处,也有缺点,因为它可能使用需要能量的过程,例如噬菌体无法获得的主动“定位”猎物。 但是,如果未发现猎物,则B. bacteriovorus不能进入休眠状态,并且会死亡。 相反,噬菌体可以在不消耗任何能量的情况下保持“生存”许多年,但无法主动寻找宿主。 他们必须依靠随机概率遇到宿主,并通过比B. bacteriovorus更快速地复制并且具有更大的爆发来补偿这种相对的低效率。
先前对伤寒沙门氏菌和大肠杆菌的研究表明,噬菌体并不总是消除其宿主的全部种群。B. bacteriovorus和噬菌体的复制都可能受到最低宿主/猎物阈值的限制,在此阈值之下,它们无法无限期地维持其种群。 然而,这可能在治疗上是有益的,因为通常不需要消灭病原体即可显着减轻或完全缓解疾病症状。
单个噬菌体的宿主范围通常限于一个菌种或一个或两个密切相关的菌种内的许多菌株,这使得其种群的维持比B. bacteriovorus更具挑战性。需要考虑宿主特异性和细菌对噬菌体耐药性的快速获得;与之相比,广泛的细菌捕食范围和缺乏掠食性细菌的简单抗药性机制使其具有优势。 有趣的是,最近有科学金工作强调了将掠食性细菌与噬菌体一起使用的组合能力。 噬菌体敏感性和Bdellovibrio捕食的独立动力学使得在某些条件下更大程度地杀死了大肠杆菌。
当被引入到吸热动物体内时,B. bacteriovorus和噬菌体都将面临挑战性条件。 与细菌噬菌杆菌相比,噬菌体在遇到苛刻的物理和化学条件(例如更高的温度和极端的pH值)时更有可能具有弹性。噬菌体可以穿过血脑屏障,但如果不进行故意修饰,可能无法在细胞内持续存在。 确实,以前的研究表明,当它们进入血液时,它们可以被网状内皮系统迅速隔离。 至少在某些情况下,可以通过在哺乳动物中连续传播噬菌体来重新分离能够在哺乳动物中长时间循环的噬菌体来应对这一问题。 众所周知,噬菌体能够通过普遍的和专门的转导在细菌之间转移DNA,这是一种不适用于B. bacteriovorus的风险。可以通过仔细筛选噬菌体基因组以除去具有整合能力的特定噬菌体和其他可能带有编码细菌毒素或其他毒力因子的基因的噬菌体,来减轻这种风险。
Bdellovibrio和噬菌体的治疗应用需要两者的工业生产。 每种噬菌体都面临着不同的挑战,因为噬菌体的产生数量更大且速度更快,但是它们的产生可能需要大量培养病原性宿主细菌。 相反,可以使用非病原性的Bdellovibrio宿主,例如大肠杆菌和恶臭假单胞菌,但是需要将它们与最终的治疗制剂有效地分开。 另外,维持Bdellovibrio制剂的生存力可能比噬菌体更具挑战性。 有趣的是,从生产这些生物防治剂的治疗制剂的技术挑战中可以学到什么教训,以及确定将它们一起用作治疗剂是否可以实现任何协同作用。
最近从事DARPA病原体捕食计划的一些研究人员,已经改变了研究领域,并证明了使用活的全食性B. bacteriovorus在体外和体内杀死广泛的抗药性(AMR)临床病原体的潜力。体内进一步支持其作为治疗手段的承诺。要被视为可靠的治疗选择,需要来自未来的人类试验的有力证据表明,这种治疗有效,至少在特定情况下,对患者没有(或最小)负面影响,并且与现有治疗相比具有一定优势。
研究人员需要证明其临床价值并衡量治疗效果,以说明对患者的益处。但现在是否拥有所需的所有信息? 如果没有,还需要解决什么?
根据体内动物模型,研究人员需要在人类身上进行安全性和有效性试验。动物模型是至关重要的,但也有局限性,迄今为止的协议管理捕食性细菌之前,或不久之后,病原体的兴趣。需要对更确定的感染进行调查,并支持确定捕食性细菌的剂量范围(以及可能的病原体数量)、剂量数量和治疗感染所需的时间表。也许令人惊讶的是,在感染和康复期间,对人体不同部位病原体实际数量的了解仍然是一个非常发展的领域。大规模捕食者生长和纯化方法的发展目前受到限制,需要扩大更大规模的安全性和有效性试验。感染治疗是时间关键,因此评估稳定性,长期储存和提供一个活的,掠夺性的积极治疗需要进一步的工作。
总之,用另一种细菌来治疗细菌感染似乎是反直觉的,离使用它们作为药物改善患者的生存状况还有一段时间。虽然蛭弧菌对其他病原的杀伤能力还有待验证,而且考虑到人体内大量必要有益的共生菌群,如何避免大量蛭弧菌“伤及无辜”也是一个值得关注的问题。需要进一步的工作来评估捕食性细菌从给药部位的传播,并确定暴露对宿主或其驻留生态位微生物群的长期影响。
但是我们相信,未来以细菌对抗细菌治疗某些感染会是一条后抗生素时代新的机遇。
相关阅读:
参考文献:
Laloux G. Shedding light on the cell biology of the predatory bacterium Bdellovibrio bacteriovorus . Front Microbiol 2020; 10:3136
Harding CJ, Huwiler SG, Somers H, Lambert C, Ray LJ et al. A lysozyme with altered substrate specificity facilitates prey cell exit by the periplasmic predator Bdellovibrio bacteriovorus. Nat Commun 2020; 11:4817
Ardal C, Balasegaram M, Laxminarayan R, McAdams D, Outterson K et al. Antibiotic development – economic, regulatory and societal challenges. Nat Rev Microbiol. 2019
Tacconelli E, Carrara E, Savoldi A, Harbarth S, Mendelson M et al. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect Dis 2018; 18:318–327
Atterbury Robert J,Tyson Jess,Predatory bacteria as living antibiotics – where are we now?[J] .Microbiology (Reading), 2021, 167.
Laxminarayan R, Van Boeckel T, Frost I, Kariuki S, Khan EA et al. The Lancet infectious diseases Commission on antimicrobial resistance: 6 years later. Lancet Infect Dis 2020; 20:e51–e60

谷禾健康
一个多世纪前,埃利·梅奇尼科夫提出了人类健康和预期寿命可以通过操纵肠道菌群来改善的概念。
现在,科学家们已经将目光投向了如何利用微生物来促进人类健康。
来自美国国立卫生研究院国家过敏症和传染病研究所的Stacy及其同事的最新研究表明,有感染史的肠道菌群能够为宿主提供更强的抗感染力。也就是说急性感染后,共生微生物群也可以被“训练”以增强对异源感染的定植抗性。
定植抗性:微生物群阻碍病原体入侵肠道生态系统,这种现象称为定植抗性。定植抗性是肠道微生物群的原始功能之一,为宿主提供了明显的益处。
训练免疫:感染或免疫可以重新编程先天免疫细胞,产生对后续感染具有广泛保护作用的记忆反应,这一过程被称为“训练免疫”。
肠道微生物群是宿主防御网络的基础。除了塑造免疫系统的发展和维护,共生微生物通过战胜病原体获得必需的营养物质和分泌抗生素来确保宿主(和它们自己)的生存,这些因素共同使肠道成为入侵的病原菌不可生存的环境。
肠道微生物群的训练总览
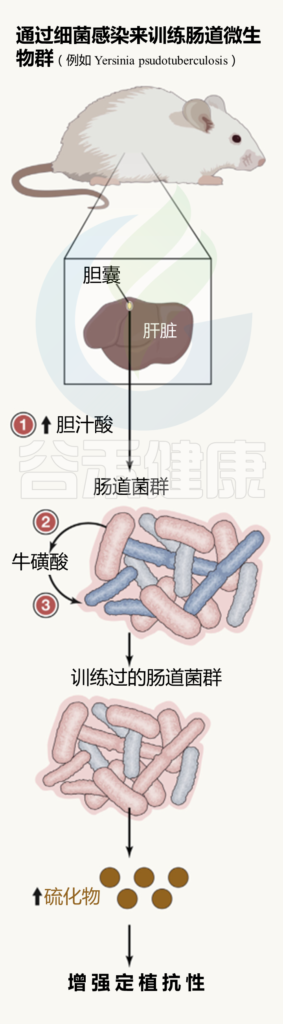
最初的肠道感染后:
( 如Y. pseudotuberculosis )
(1) 肝脏中与牛磺酸结合的胆汁酸产生增加,储存在胆囊,并释放到肠道;
(2) 在肠道中,胆汁酸被特定的微生物群化学解共轭;
(3)牛磺酸变得可利用,导致产生硫化氢的细菌大量繁殖,从而阻止肠道病原体的有氧生长。
下面我们来看Stacy等人是如何一步步进行的实验探究过程。
Stacy等人在研究中发现,与实验室特有的无病原体(SPF)微生物群的小鼠相比,拥有野生小鼠(即WildR)微生物群的自交系小鼠表现出对致命病毒感染的增强保护。
在这里,作者还发现野生小鼠比SPF小鼠对肠道肺炎克雷伯菌感染更具抵抗力。考虑到WildR小鼠的未知感染史,作者用肺炎克雷伯菌对以前感染过急性Yersinia pseudotuberculosis ( post-△yopM) 的SPF小鼠进行了挑战。
值得注意的是,post-△yopM 的小鼠在初次感染后的至少15周内对肺炎克雷伯菌感染的抵抗力也更强。与SPF重建的对照组相比,用post-△yopM小鼠的粪便重建无菌小鼠也增强了对肺炎克雷伯菌感染的保护,这证实了微生物群在抗性中的作用。
这些结果表明,感染史可以改变肠道微生物群,从而诱导定植抗性,因此称为“微生物群训练”。
Stacy等人使用代谢组学方法记录了清除了假结核杆菌Y. pseudotuberculosis△yopM的小鼠肠道中胆汁酸水平的升高。微生物群对初级胆汁酸的解偶联释放了偶联组分,如牛磺酸。
注:牛磺酸在胰腺中合成,对肌肉和大脑发育至关重要,是哺乳动物中最丰富的游离氨基酸。最近发现牛磺酸可以激活杯状细胞中的NLRP6炎症小体,并通过肠道上皮细胞调节稳定状态的抗微生物肽的产生。
在短暂的假结核杆菌感染后牛磺酸水平升高。之前的研究表明,牛磺酸支持解决肠道炎症和清除肠道病原体的柠檬酸杆菌感染。
他们发现,补充牛磺酸会损害柠檬酸杆菌C. rodentium和肺炎克雷伯菌的肠道定植。而当补充给无菌小鼠时,牛磺酸对肠道感染没有影响,这表明它通过调节共栖群落而不是直接影响病原体。
从机制上讲,牛磺酸可增强微生物群中硫化物的产生,硫化物是细胞呼吸的抑制剂,这是宿主被多种病原体入侵的关键。
Stacy等人用16S rDNA和宏基因组测序来解决这个问题。肠道炎症通常与肠杆菌科(Enterobacteriaceae)的大量繁殖有关。
参与对抗感染菌– δ 变形菌 Deltaproteobacteria
他们发现,与对照组小鼠相比,清除假结核杆菌△yopM感染的小鼠微生物群中变形菌门增加,特别是δ 变形菌纲(Deltaproteobacteria)。
宏基因组学揭示,δ 变形菌纲的这种繁殖导致感染小鼠的硫代谢途径增加。虽然在感染后的微生物群中没有发现代谢硫的菌(δ 变形菌纲Bilophila wadsworthia),但当补充到无菌小鼠中时,足以提高对肺炎克雷伯菌的定殖抗性。
牛磺酸的施用同样影响了微生物群;然而,这些变化与post-Yersinia状态相当不一致,包括梭状芽孢杆菌而不是δ变形菌的增殖。这些不一致的分类变化表明,仅补充牛磺酸并不能完全重现共生微生物群的感染后状态,多种因素和微生物群可能能够增强对继发感染的定殖抗性。
Stacy等人通过宏基因组学检测牛磺酸处理过的小鼠微生物组,发现与post-Yersinia菌感染后的微生物组相似,编码异化亚硫酸盐还原酶(dsr)的基因得到了富集。这种蛋白质参与牛磺酸酶转化为硫化物的最后一步。
当牛磺酸调节的微生物群在体外培养中暴露于牛磺酸时,会产生更多的硫化氢,这与肠道中牛磺酸水平较高支持能将这种代谢物转化为硫化物的物种生长的观点一致。
重要的是,硫化氢通过抑制细胞色素氧化酶来限制有氧呼吸,而细胞色素氧化酶是病原体在不可发酵的底物上进行有氧呼吸和生长的常用物质。因此,牛磺酸依赖的硫化氢的产生可能通过阻断有氧呼吸来提高定植抗性。
原发感染后牛磺酸代谢菌群的扩张增强了对继发感染的定植抗性
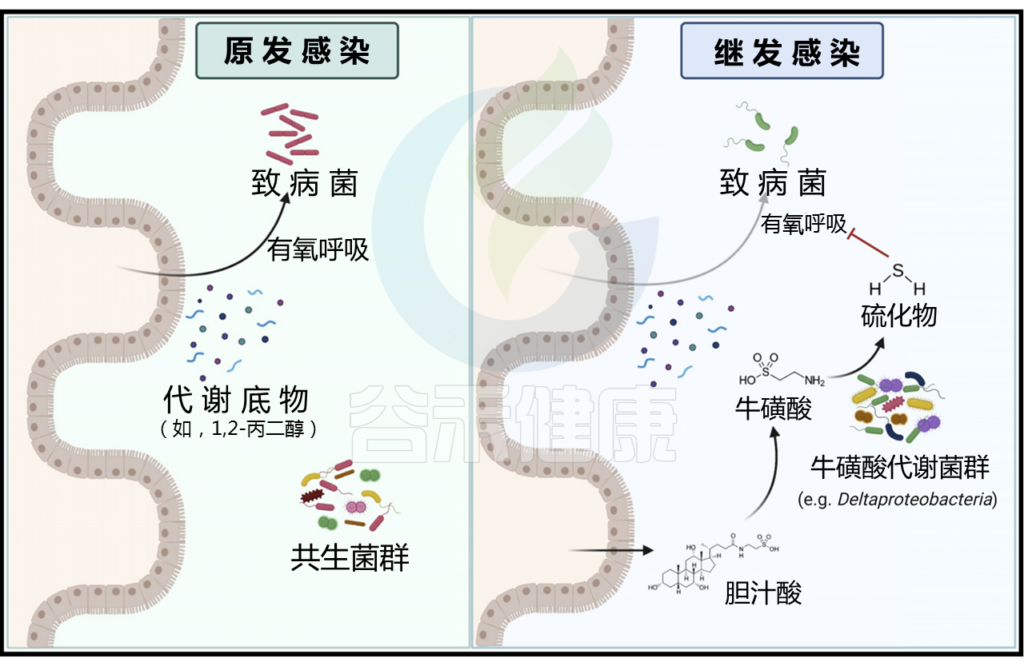
在原发感染(左)期间,病原体通过有氧发酵利用代谢底物,如1,2-丙二醇。清除感染后,宿主产生的胆汁酸水平增加,可以通过共生微生物群与牛磺酸解偶联。牛磺酸反过来被代谢成硫化物,抑制有氧呼吸,从而增强对继发感染的定植抗性(右)。
Stacy等人用两种方法验证:
(1) 使病原体不能在不可发酵的物质上生长
Stacy等人对肺炎克雷伯菌进行了高通量转座子测序。他们让12000个转座子突变体通过小鼠,然后测序,以确定与输入库相比丢失的突变体。
在未感染的小鼠中,导致细菌适应的因素包括与1,2-丙二醇利用有关的基因以及与细胞色素氧化酶bd-II同源的假定氧化酶。因此,肺炎克雷伯菌在低氧环境下对1,2丙二醇作用最为有利,低氧环境受硫化物抑制。
此外,缺乏在1,2-丙二醇或细胞色素氧化酶II上生长能力的肺炎克雷伯菌突变体在体内被它们的野生型对应物所取代,但当牛磺酸被补充到肠道生态系统中时却没有这种情况,表明牛磺酸破坏了病原体在1,2-丙二醇上的呼吸和生长能力。
(2) 隔离硫化氢
Stacy等人使用铋来隔离硫化物。铋对小鼠的处理阻止了微生物群产生硫化氢,导致了具有有氧呼吸功能的细菌数量的增加。
重要的是,铋处理还增加了对肺炎克雷伯菌感染的易感性,突出了硫化物隔离对定植抗性的影响。
这种对继发感染的增强抗性似乎是高度非特异性的,因为弱毒假结核分枝杆菌的初次感染可以防止肺炎克雷伯氏菌或柠檬酸杆菌的继发感染。
事实上,Stacy等人使用“wildR”小鼠的微生物群发现了相同的效果,这些小鼠是无菌小鼠,与野生小鼠的微生物群一起定居,野生小鼠可能有许多肠道感染的历史。
野生小鼠的肠道也显示出δ变形菌纲的扩张,但牛磺酸的水平降低而不是升高,这表明感染后牛磺酸可能会随着时间的推移而消耗,或者是不同的机制导致了这些小鼠中δ变形菌纲的大量繁殖。
Stacy等人没有用相同的病原体进行原发性和继发性感染,因此尚不清楚保护机制是否也适用于这种情况,以及增强定殖抗性的进化目的论是否是为了避免用相同的病原体进行二次感染。同样,确定感染后微生物群是否对不依赖硫化物敏感有氧呼吸的病原体无效也很重要。
Stacy等人的发现也存在一些问题。
首先,为什么在成功清除感染后胆汁酸分泌持续升高?
作者在post–Yersinia感染后的小鼠中发现了增大的胆囊,但这可能不是主要原因。
二、这种增强的殖民抵抗的状态持续多久?
Stacy等人在post-Yersinia感染后15天观察到,接受来自小鼠体内微生物群的无菌小鼠的定殖减少,但尚不清楚肠道牛磺酸水平是否表现相似,以及保护作用是否在超过15天的时间内减弱。
同样,确定多轮感染是否会进一步增强最终的定植抗性能力也很有意思。
Stacy等人的工作使用了生物化学、微生物学和生物信息学方法等,不仅形成了一个关于感染史如何导致肠道-肝脏保护轴的新概念框架,使“微生物群记忆”的概念得到推广,还提供了对宿主及其微生物群之间存在的互利途径的新见解。
这可能是通过精确调节微生物群来提高抗病原体入侵能力的重要一步。
相关阅读:
主要参考文献:
Stacy, A., Andrade-Oliveira, V., McCulloch, J.A.,Hild, B., Oh, J.H., Perez-Chaparro, P.J., Sim, C.K., Lim, A.I., Link, V.M., Enamorado, M., et al. (2021). Infection trains the host for microbiota-enhanced resistance to pathogens. Cell 184, this issue, 615–627.e18
Wong Andrea C, Levy Maayan,Microbial memories.[J] .Immunity, 2021, 54: 201-204.
King Irah L,Divangahi Maziar,Training the metaorganism: the microbial counterpart.[J] .Cell, 2021, 184: 574-576.
Rosshart, S.P., Vassallo, B.G., Angeletti, D.,Hutchinson, D.S., Morgan, A.P., Takeda, K., Hickman, H.D., McCulloch, J.A., Badger, J.H., Ajami, N.J., et al. (2017). Wild Mouse Gut Microbiota Promotes Host Fitness and Improves Disease Resistance. Cell 171, 1015–1028.

谷禾健康
功能性腹痛性疾病(FAPD)是儿童期最常见的疾病,全世界25%的儿童和婴儿受其影响。
功能性腹痛性疾病
小儿功能性腹痛性疾病,目前称为肠脑相互作用障碍,包括肠易激综合症,功能性消化不良,腹型偏头痛和功能性腹痛。
<罗马IV诊断标准见附录>
近日,英国和美国的研究科学家联合发表在《NATURE REVIEWS | DISEASE PRIMERS》上的一篇综述文章题为:“Paediatric functional abdominal pain disorders”(小儿功能性腹痛症)。


该文总结了FAPD患病率和致病性的最新进展,并特别关注了早期生命的影响和构建。对食源性发病机理和病理生理学有了更深入的了解,包括肠道器官(炎症,运动和微生物群),核心因素(心理方面,某些大脑区域的敏感性和/或连通性或活动性差异)以及外在因素(感染)。
此外,讨论了当前的诊断方法并着重介绍了这些疾病的治疗选择,特别是IBS和FAP-NOS。强调可用的药理干预措施仅限于儿童,因此,干预管理重点落在综合方法上,包括针对精神的干预措施(催眠疗法和认知行为疗法),饮食(益生菌)和经皮神经电刺激。这些方向将有助于理解病理生理学并更好地治疗这些疾病。
FAPDs的特征通常内脏痛觉过于敏感以及内脏刺激中枢感知增加而导致残疾,是遗传易感性和早期生活事件背景上叠加的社会心理因素和医学因素的最终结果。
功能性消化不良的症状特征主要来自胃肠道近端,涉及上腹部腹痛和/或餐后饱胀和/或早期饱腹感。
腹型偏头痛的特点是发作性长时间发作,剧烈,急性脐周,中线或弥漫性腹痛。这些痛苦的发作通常无能为力并干扰正常生活。
这些发作通常但不总是与其他症状(例如厌食,恶心,呕吐,头痛,畏光等)相关,这些症状可能在疼痛持续时间之前或与疼痛持续时间同时发生,并且这种症状性发作可能相隔数周至数月不等。
风险因素如下:


FAPDs是一种复杂的疾病,似乎是由于一个或多个元素的功能和/或更微妙的结构完整性的破坏造成的菌群-肠-脑轴。这些条件下相互作用的复杂性和多面性被整合到一个生物心理社会模型中。
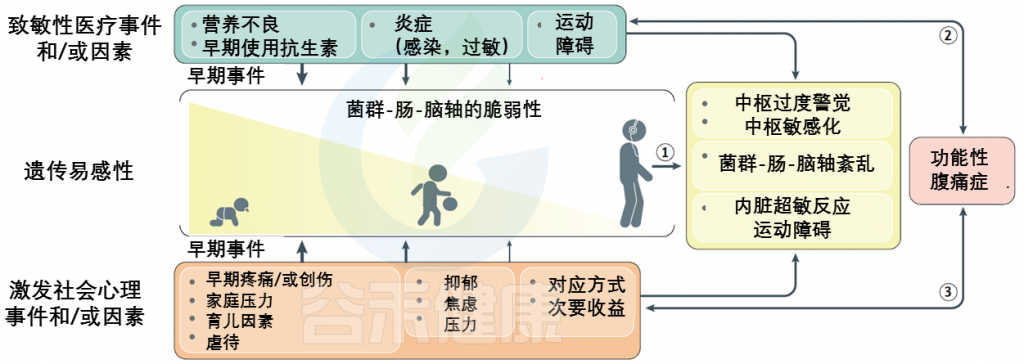

该模型的关键组成部分是内脏超敏反应和中枢敏感化。
内 脏 超 敏 反 应
内脏超敏反应描述了对周围信号的知觉反应(即痛觉过敏),并且可能是内脏传入信号处理变化的结果或疼痛下调变化的结果(例如,中枢敏感化)。
表现:疼痛的感觉阈值降低,也就是稍微一点点刺激就会感到疼痛。
可能原因:在肠扩张期间或由于暴露于食糜的化学成分改变引起,部分解释了对诸如饮食调整和抑酸等治疗的反应。
潜在诱因:炎症,感染和压力
从本质上讲,这些触发因素可引起粘膜通透性变化并导致炎症,释放促生因子,包括5-羟色胺,组胺,NGF,蛋白酶和前列腺素,它们能够激活传入伤害感受器(痛觉神经)上的受体,引起急性疼痛以及持久的功能和结构改变,这是维持慢性疼痛的关键。
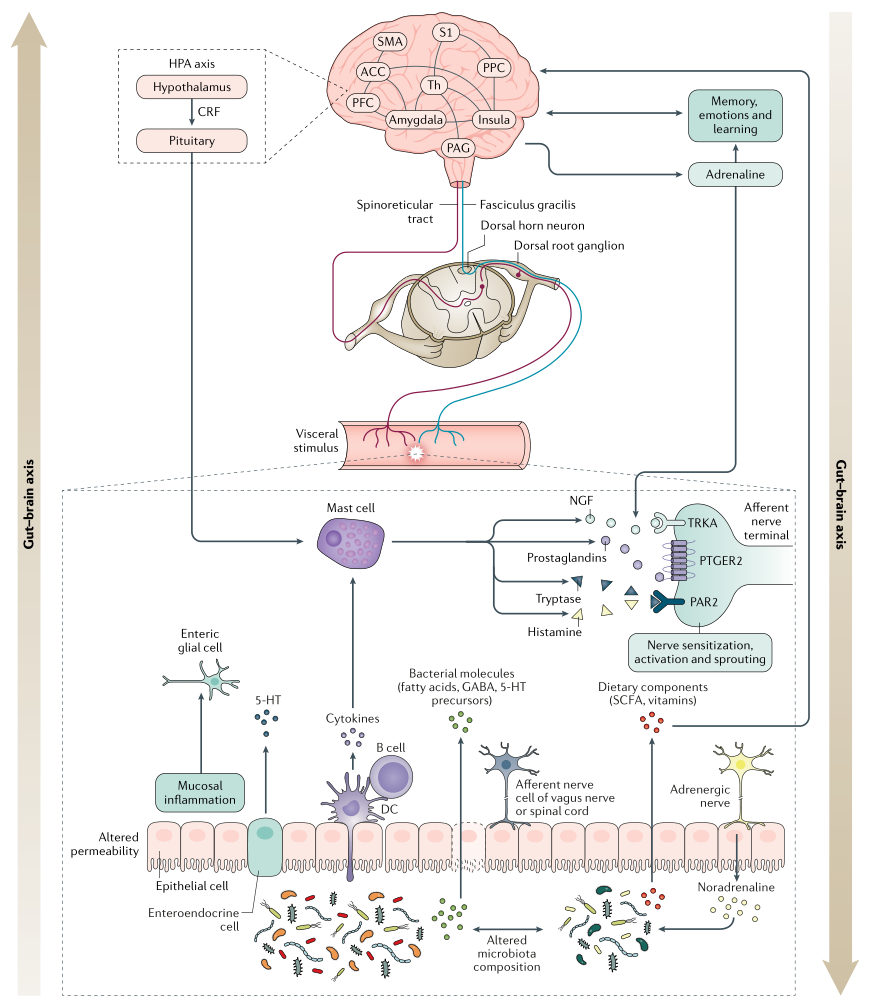

中 枢 敏 感 化
中枢敏化是指通过增强中枢神经系统内的神经元功能和神经信号来放大疼痛敏感性的现象,会引起疼痛超敏反应,并且是一种发展和维持慢性疼痛的机制。
除了疼痛超敏反应外,中枢敏化还会导致脑部活动的继发性变化,可以通过电生理或成像技术进行检测。患有FAPD的个体可能更倾向于发展来源不明的中央致敏。
2018年进行的一项系统性回顾涉及12项病例对照研究,发现功能性腹痛儿童的继发性痛觉过敏和皮质伤害感受改变。
生 命 早 期 事 件
在小儿FAPD中,损伤时机比导致微生物群-肠-脑轴成分破坏的多种因素或该轴本身的破坏更为关键。在生命的头二十年中,尤其是从围产期到儿童时期直至青春期,支撑其功能的肠脑轴所有元素的发育和成熟的关键过程至关重要。
包括手术(例如脐疝和幽门狭窄),细菌性胃肠道感染,炎性或免疫介导的疾病(例如乳糜泻,炎性肠病),以及在生命的最初几个月中发生无害的侮辱,例如婴儿期对牛奶过敏和早期使用抗生素等。
神 经 免 疫 相 互 作 用
在一些患有FAPD的儿童中,证据表明存在低度的肠道炎症,并在发病机理中发挥了神经免疫相互作用的作用。
一项研究发现,IBS患儿回结肠结肠粘膜中靠近神经的肥大细胞数量增加。神经纤维相关的肥大细胞计数与腹痛的强度和疼痛发作的频率相关。
胃 肠 蠕 动
胃肠动力异常促进FAPD的病理生理。具有上消化道症状的患者可能有胃排空延迟和/或胃适应性降低或两者都不存在。功能性下消化道症状的患者可能延迟,加速或正常结肠转移,和/或骨盆底运动障碍(肌肉协调障碍导致试图放松骨盆底肌肉时收缩)。
—功能性消化不良
—肠易激综合征
微 生 物 群
FAPD与肠道微生物群的多样性和菌群组成有关,尤其是IBS。2019年,一项系统评价86评估了健康个体和IBS患者(成人和儿童)的肠道菌群组成。发现双歧杆菌属的丰度下降。IBS患者中的抗炎作用与Faecalibacterium spp,尤其是Faecalibacterium prausnitzii一样,具有抗炎作用。
IBS患者的粪便微生物群的多样性降低或保持不变。粪便微生物群组成或代谢组组成可以预测患者可能从饮食中获益。
除细菌外,真菌或病毒也可能在FAPD病理生理中起作用。但是,有关真菌菌群失调或病毒菌群失调的可用数据有限,目前的研究主要集中于炎症性肠病,而不是IBS。
营 养
在患有FAPD的儿童中起着至关重要的作用。
饮食失调(例如神经性贪食症)可能导致IBS患儿肥胖的发生率增加。
碳水化合物的吸收不良可引起胃肠道症状。吸收不良可能与特定的酶缺乏症有关(例如乳糖酶缺乏症中的乳糖;蔗糖和异麦芽糖酶缺乏症中的蔗糖和淀粉)或与碳水化合物(如果糖,山梨糖醇或甘露醇)的吸收能力有限有关。
在糖较小的情况下,所引起的渗透负荷可导致肠腔膨胀小和快速转运。对于诸如果聚糖等较大的糖,人不具有消化的酶促能力,因此,吸收不良的糖会被结肠菌群发酵,从而导致气体形成和潜在的结肠扩张。
心 理 因 素
肠脑轴的生理很复杂,包括传入和传出成分。
下丘脑-垂体-肾上腺轴通过皮质醇和促肾上腺皮质激素释放因子的释放(在压力条件下均会增加),以及通过刺激肥大细胞和诱导脯氨酸释放的反馈回路,促进肠-脑相互作用。
研究表明,在成年人和有FAPDs的儿科患者中,压力都会增加肠道炎症标志物,如粪便钙卫蛋白和CRP。
肠道与大脑的相互作用涉及自上而下和自下而上的过程,因此,肠道菌群也可以影响脑功能。在压力条件下,肠道菌群可以增加上皮屏障的通透性,使抗原和/或病原体能够通过并引起炎症反应。由此产生的循环性促炎细胞因子可能会与中枢神经系统通讯,刺激大脑的免疫反应,从而可能导致或加重心理症状(例如焦虑和抑郁)。
几项研究表明,与健康儿童相比,患有家族性帕金森病的儿童的心理压力和行为问题有所增加。心理因素可能既是FAPD的起因,也是后果。

无论采用哪种治疗方法,患者与临床医生之间有效的沟通和病人教育,是成功进行FAPD管理的核心。必须花足够的时间向每位患者的照护者解释诊断,并讨论生物心理社会模型。
FAPD的管理受到几个因素的限制
1. 由于FAPD生物心理社会特性,每个孩子都有一套独特的病理生理因素,并且对疗法的反应也不同。
2. FAPD患儿的证据基础很小,许多治疗建议都是基于对成年人的研究。 儿童通常不会像成年人那样做出反应。
3. 一些有效的治疗方法是行为疗法(饮食疗法和心理疗法),由于缺乏专职医疗保健专业人员以及缺乏保险,这些方法并不容易。
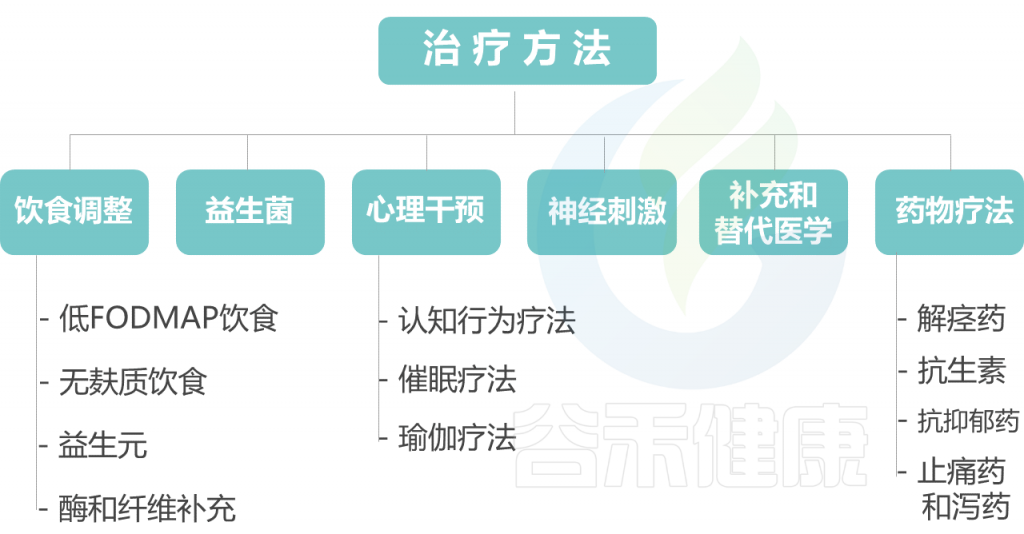

以下对每种治疗方法展开讨论。
饮 食 调 整


益 生 菌
多项研究检查了益生菌治疗FAPD的功效(大多成年IBS患者)。这些研究表明,益生菌或特定菌特定组合可能是有效的,但研究有局限性,如样品量,致盲性,所用益生菌的差异以及不同的剂量,其作用仍不清楚。
在患有IBS的儿童中,两项研究发现鼠李糖乳杆菌GG可有效减轻腹痛症状,而一项研究则报告腹痛没有改善,但感觉到的腹胀有所改善。在不同的研究中,疼痛程度再次有所不同。
在一个多中心,IBS儿童中进行的交叉RCT研究中,VSL#3(八种益生菌菌株的混合物)在改善症状和改善生活质量方面比安慰剂更安全,更有效。
心 理 干 预
心理干预迄今为止,诸如CBT和催眠治疗等心理干预已被证明是管理FAPD的最成功干预措施,并主张将其普遍纳入管理策略。
认知行为疗法(CBT)
CBT是治疗IBS的研究最多的心理疗法,其目的是改变认知,情绪和行为,这些不良情绪行为等都可能加剧或维持IBS症状。
在成人和儿童中的研究表明,与教育和其他控制干预措施相比,CBT在改善疼痛和参与社交生活的能力以及减轻焦虑或抑郁症状方面是有效的。
十二项随机对照试验涉及990名IBS儿童(7-18岁),显示出CBT对生活质量,学校表现和社会参与产生了长期的,持久的有益影响。CBT可以作为面对面的治疗提供,也可以通过互联网面向儿童或通过电话面向父母。
已报道,CBT在改善肠易激综合征儿童的疼痛和残疾方面具有有效性。患有家族性帕金森病的儿童的社区康复治疗主要集中在学习应对症状和减少残疾上,这些治疗应该是综合护理方法的理想组成部分。
催 眠 疗 法
在治疗师的指导下,患者会进入催眠状态,以响应改变主观体验,感知,情感,思想或行为的建议。
在成年人中,催眠疗法已被证明可导致结肠运动改变,改善内脏超敏性并减少长期持续存在的心理因素,例如躯体化和心理压力。
然而,在IBS患儿中,尽管接受了标准疗法的儿童的疼痛评分和躯体化评分均较低,但在催眠治疗后并未发现超敏反应降低(通过实验测试)。
IBS或FAP-NOS(n = 412;6-18岁)儿童的5个随机对照试验,无论是单独治疗还是在家中通过CD进行催眠治疗,均产生了长期有益影响。
在1年和5年的随访中,接受催眠治疗的患者中分别有85%和68%已没有症状,而对照组中分别只有25%和20%的患者缓解
瑜 珈 疗 法
瑜伽练习已被证明可以改善应激诱导的副交感神经系统活性不足。三个随机对照试验,包括127名IBS儿童(7-18岁),瑜伽疗法在减少腹痛方面具有积极作用。
但是,一项荷兰研究表明,在1年的随访中,瑜伽干预并没有比不包括CBT的标准治疗更有效。由于学习方法的重大缺陷,尚不能提出将瑜伽作为FAPD儿童的常规干预措施的建议。
神 经 刺 激
多项研究表明,脊髓和大脑的电刺激在调节疼痛途径中的功效,能够减少脊髓和中央杏仁核中50%以上的神经元放电,从而减轻内脏疼痛。
一项研究表明,经皮神经电刺激(PENFS)可以调节杏仁核和脊髓神经元的反应特性,并显着降低大鼠内脏超敏反应的发展。
随后,同一组受试者在115名FAPD的青少年(11至18岁)的随机对照试验中显示,具有主动装置的PENFS可以改善健康状况,并显着减轻疼痛和残疾 与假刺激组。
此外,PENFS的有益作用在2个月的随访期间得以维持。尽管有些耳部不适,但未报告严重的不良反应。
补 充 和 替 代 医 学
补充和替代医学包括不同的方法,从针灸和阿育吠陀医学到脊椎指压疗法、整骨疗法、顺势疗法、精神治疗、按摩和冥想等身心技术。大约40%的澳大利亚和荷兰诊断为糖尿病肾病的儿童使用其中某些替代疗法,其中草药治疗是最常见的(46%)。
许多补充疗法被公众认为是“自然的”,比现代医学的医疗设备更安全、更温和。FAPDs儿童对对抗疗法药物相关可能产生不良反应的恐惧,常规治疗没太大效果,让父母希望寻求补充和替代治疗。
然而,到目前为止,评估草药疗法、针灸、顺势疗法、身心疗法或肌肉骨骼操作(如整骨疗法和脊椎指压疗法)对患有家族性帕金森病的儿童的效果的随机对照试验尚不可用。
由于糖尿病周围神经病变的自发缓解率很高(30-70%),采取逐步的管理方法是明智的;必要时,教育、识别和改变压力因素和饮食干预可能是第一步。当症状持续或再次出现时,下一步可能是开始一种心理治疗,如认知行为疗法和催眠疗法,或PENFS。
药 物 疗 法


胃肠症状的严重程度和发生频率对不同患者的功能状态有不同的影响。患者报告的健康相关生活质量(HRQOL)测量应该有助于深入了解FAPDs对儿童和青少年功能状态的不同影响,包括身体、情感、行为、社会和认知方面。
评估症状和HRQOL影响的患者报告结果已成为成人临床状态和治疗结果的公认指标。但在儿童环境中,自我报告的HRQOL措施在仪器开发和临床应用中遇到了挑战,部分原因是担心儿童能否可靠地自我报告与健康相关的信息。
量化个体胃肠道症状对于开发以患者为中心和针对症状的干预措施很重要,这可以改善整体HRQOL。例如,一项针对259名患有功能性便秘,功能性腹痛或IBS的儿科患者的多中心研究发现,胃肠道症状最能预测整体受损。
FAPD在儿童时期很常见,FAPD患病率和/或复杂性可能会增加。因此,需要有来自所有地理区域以及世界各地精心设计的流行病学研究,才能真正了解问题的性质和规模。
特别重要的是,我们目前对导致儿童FAPD发病机理的潜在因素的了解,需要使用来自FAPD患儿的组织样本来解决免疫功能障碍和神经功能障碍的基础和转化科学研究。如上所述,早期的影响,即在复杂的肠脑轴各要素发展的关键阶段发生的事件,通常是辱骂或创伤(心理或医学方面的事件),使易感人群容易患上FAPD。
异常改变肠道菌群的因素可能会改变肠神经系统的结构或功能完整性,进而改变其与中枢神经系统的复杂且平衡的相互作用。结果可能是胃肠道重新调整为过敏状态,而大脑则重新编程为过度警觉状态。
当然,其他遗传因素,社会因素和生理变化,例如在青春期或青春期发生的那些,也可能促进症状发展。
现代工具越来越多地与焦虑和压力相关联,也可能会对FAPD产生重大影响。
总体而言,在过去十年中,儿童家庭保护方案取得了相当大的进展。对所有影响因素,及其与触发FAPD最终发展有关时机的更深入了解,可能带来更合适的预防或治疗策略。
【附录】
功能性腹痛疾病的罗马IV诊断标准
肠易激综合征
该标准必须满足至少2个月,并包括以下所有:
• 每月至少有4天腹痛伴有排便和/或排便次数的改变和/或大便外观的改变
• 腹痛不能随着便秘的解决而消失(疼痛消除的儿童患有功能性便秘,非肠易激综合征)
• 经过适当评估,症状不能完全由另一种疾病解释
功能性消化不良
诊断前必须至少满足2个月的标准,并且必须包括以下一种或多种症状,每月至少4天:
• 餐后饱腹感
• 早期饱腹感
• 与排便无关的上腹部疼痛或灼热感
• 经过适当评估,这种症状不能完全由另一种医学状况来解释
腹型偏头痛
必须在诊断前至少6个月内达到标准,并包括以下所有至少发生两次的情况:
• 持续1小时或更长时间的剧烈、急性脐周、中线或弥漫性腹痛的阵发性发作(应为最严重和最痛苦的症状)
• 发作间隔数周至数月;疼痛使人丧失能力并干扰正常活动;个体患者的定型模式和症状
• 疼痛与以下两种或两种以上症状相关:厌食、恶心、呕吐、头痛、畏光或苍白
• 经适当评估,症状不能完全由另一种医学状况解释,
功能性腹痛
未另行规定标准必须在诊断前至少2个月完成,每月至少四次,包括以下所有内容:
• 并非仅在生理事件(例如,饮食和月经)期间发生的偶发性或持续性腹痛;肠易激综合征、功能性消化不良或腹部偏头痛的标准不足
• 经过适当评估,腹痛不能由其他医疗状况完全解释
相关阅读:
主要参考文献:
Sjölund, J. et al. Prevalence and progression of recurrent abdominal pain, from early childhood to adolescence. Clin. Gastroenterol. Hepatol. 2020
Paediatric functional abdominal pain disorders.[J] .Nat Rev Dis Primers, 2020, 6: 88.
Rutten, J. M. T . M., Korterink, J. J., Venmans, L. M. A. J., Benninga, M. A. & T abbers, M. M. Nonpharmacologic treatment of functional abdominal pain disorders: a systematic review. Pediatrics 135, 522–535
Masuy, I., Van Oudenhove, L. & T ack, J. Review article: treatment options for functional dyspepsia. Aliment. Pharmacol. Ther. 49, 1 134–1 172 (2019)
Russell, A. C., Stone, A. L. & Walker, L. S. Nausea in children with functional abdominal pain predicts poor health outcomes in young adulthood. Clin. Gastroenterol. Hepatol. 15, 706–71 1 (2017)
Maragkoudaki, M. et al. Lactobacillus reuteri DSM 17938 and a placebo both significantly reduced symptoms in children with functional abdominal pain. Acta Paediatr. 106, 1857–1862 (2017).